
浙江古田山亚热带常绿阔叶林叶衰老物候影响因子研究
2023-04-11 03:34:38寿佳君薛乾怀王鑫洋唐欣然陈沁杜彦君
热带亚热带植物学报 2023年2期
寿佳君, 薛乾怀, 王鑫洋, 唐欣然, 陈沁, 杜彦君
浙江古田山亚热带常绿阔叶林叶衰老物候影响因子研究
寿佳君, 薛乾怀, 王鑫洋, 唐欣然, 陈沁, 杜彦君*
(海南大学林学院,海口 570100)
该文选取浙江省古田山亚热带常绿阔叶林72种木本植物,探究气候因素、系统发育关系和功能性状对亚热带常绿阔叶林叶衰老物候的影响。结果表明,叶变色期在9—12月,落叶期在10—12月。每月落叶物种数与月均温、月均降水量和月均日照时数没有显著相关性,每月叶变色物种数与月均温和月均日照时数呈弱相关;落叶性对叶变色期和落叶期具有显著影响;植物间系统发育关系对叶变色期和落叶期没有显著影响。因此,生物和非生物因子都会影响常绿阔叶树种的叶衰老,这对于提高秋季物候预测模型具有重要价值。
叶变色物候;亚热带常绿阔叶林;落叶;功能性状;系统发育保守性
物候学是研究自然界植物、动物和气候、水文、土壤等环境条件的周期变化之间相互关系的科学[1]。植物物候是指示气候变化的重要指标,其周期性变更是植物适应环境变化的重要方式[2]。自工业革命以来,全球气温上升了0.6 ℃,春季和冬季平均温度上升了1 ℃以上,气候对植物物候期产生了明显的影响,具体表现为春季物候提前,夏秋物候推迟,植物生长季节延长[3]。在全球气候变化的大背景下,物候学作为一门传统科学体现了其重要价值,物候记录直接反映综合影响下生物对环境的响应,是全球环境变化的最直接和最有效的证据[4]。
植物叶片作为植物光合作用的主要器官,它对生态系统的能量流动和物质循环十分重要[5]。目前,国外对植物叶物候已经开展了不少研究[6–7],而国内只是进行了展叶物候期调查[8]。植物叶物候主要包括展叶、叶变色和落叶等,研究叶物候动态可以了解植物的生活史策略以及不同物种对环境的竞争。目前对于植物展叶物候研究较多,研究焦点主要聚集在展叶物候。和展叶物候一样,植物的叶衰老,即叶变色和落叶,对于树木适应环境同样重要,它们共同决定了植物生长季节的长度,共同影响了植物的生长与发育,决定了植物对碳的固定和对环境的适应[9]。因此,亟需对叶变色和落叶物候的格局及其驱动因子进行研究[9]。
影响叶衰老的因子主要包括非生物因子和生物因子。非生物因素主要包括光周期、温度、降水等因素。有研究表明日照时间的减少[10]、温度降低[6]、降水减少[11]会促进叶衰老。Bullock等[12]在查尔梅的研究表明,一些植物落叶受日照时间的影响,如红鸡蛋花();吴朝阳等[13]利用14 536个地面时间序列和30多年遥感观测研究表明,随着全球气候变暖,北半球秋季叶衰老的总体开始时间将比目前预期的要早,特别是在干旱地区。Zani等[9]在温带森林对秋季物候研究表明温度、光照水平升高等因素导致了叶衰老期的提前。此外,叶衰老物候也可能受到生物因子的调控,如系统发育关系[14]、其他功能性状如生活型[14]等。Panchen等[14]在阿诺德植物园和柏林植物园的研究表明,乔木比灌木和藤本植物叶衰老期更早,这可能是由于垂直温度的差异使得乔木冠层的温度低于灌木[15],或者是由于长期的环境选择使得乔木对温度的敏感性高于灌木[15]。同时,Panchen等[14]还认为叶衰老期与系统发育无关。然而,目前对叶衰老物候影响因素的研究主要集中在温带树种,对亚热带树种的研究十分缺乏。
亚热带常绿阔叶林蕴含着丰富的生物多样性,具有十分重要的生态价值和经济价值。我国幅员辽阔,亚热带面积大,亚热带常绿阔叶林作为热带雨林和温带森林间的过度类型广泛分布于我国东部地区,是一种典型的地带性植被类型。由于亚热带气候条件具有明显的多样性和过渡性,植物物候可能与温带地区有所不同。为探究亚热带常绿阔叶林中叶衰老物候的影响因素,本研究选取浙江古田山亚热带常绿阔叶林24 hm2样地内的72种木本植物,主要探究温度、降水、日照时数等气候因素及植物亲缘关系和功能类型对落叶物候和叶变色物候的影响,这不仅对了解亚热带植物秋季物候的影响因素,而且为研究植物对环境的适应机制具有一定的意义。
1 材料和方法
1.1 研究区概况
研究地点位于浙江省衢州市开化县苏庄镇境内的古田山国家级自然保护区(118°03′50′′~118°11′ 12.2′′ E, 29°10′19′′~29°17′41′′ N)。该样地地处中亚热带东部,浙、赣、皖三省交界处,呈现山地地貌,地形复杂多变,又由于地处中亚热带湿润季风区, 古田山四季分明,气候变化明显,分布着典型的中亚热带常绿阔叶林。古田山山势呈东北西南走向, 主峰青尖峰海拔1 258 m。古田山年均温为15.3 ℃,年均降水量为1 787 mm。每年3—6月和9月为雨季, 7—8月是伏旱期,10—2月为干季[16]。古田山24 hm2样地中共有159种约140 700棵胸径(diameter at breast height, DBH)大于1 cm的乔木和灌木[17]。
1.2 物候数据收集
2018年11月,在古田山24 hm2样地内选取72种木本植物,每物种随机选取1~12株,共286株进行秋季物候观测,2018.11—2021.02每隔2周记录地面数据[18]。使用双筒望远镜进行观测,主要观察并记录树木冠层叶变色和落叶百分比以及观测日期。叶变色和落叶百分比分别指植物冠层叶片变色数量占冠层总叶片数量的比例以及冠层落叶部分占冠层总叶片数量的比例。在计算落叶百分比时,观测人员除了看冠层的落叶情况还会辅以观察已经落到地面上的叶片进行判断。一般来说,常绿阔叶林树种一年四季都在不停的落叶和展叶[19]。因此只有当植物冠层叶变色百分比大于5%时,才将该天的日期记为该株植物叶变色开始的时间。在植物叶变色的基础上将植物落叶百分比大于5%的日期记为该株植物落叶开始时间。对于每个个体,每年只记录叶变色期和落叶期各1个日期,一旦地面监测记录表中叶变色或者落叶达到5%以上, 就完成了对其叶变色期和落叶期的记录。
1.3 气象数据
由于无法获取开化县的气象数据,本次研究利用月度气候数据进行分析,包括月均温和月均降雨量(2018—2020年),均从“气候研究单位0.5°×0.5°栅格数据集”上获取(CRU TS ver. 4.04, http://www.cru. uea.ac.uk/)[20],同时我们按照纬度换算对应的日照时间,并计算月均日照时数。
1.4 功能群分类
按照《中国植物志》(http://www.iplant.cn/frps)中的分类原则,将所有目标树种的生活型分为灌木型和乔木型;根据树木的落叶性分为落叶型树种和常绿型树种。依据果实类型分为非肉质果实型和肉质果实型。翅果、蒴果、荚果、坚果等果肉干瘪水分少的称为非肉质果实类型,而肉质聚合果、柑果、浆果、核果和梨果等果肉中富含水分的则为肉质果实类型[21]。根据功能性状,本研究的72种木本植物可分为36种灌木和36种乔木;37种干果和35种肉质果;34种落叶树种和38种常绿树种(表1)。这些植物大多为古田山的本土优势树种,如林冠层优势种有甜槠()、米槠()、木荷()、短柄枹()和马尾松()等[22]。
1.5 构建系统发育树
利用系统发育树和系统发育信号Blomberg’s来检验系统发育对物种间叶变色期和落叶期变异的约束条件。用Blomberg’s值法计算的谱系信号衡量功能性状的系统发育信号强度[23]。利用Kress等的DNA条码技术,建立古田山定位站24 hm2样地159种物种的系统发育树[24]:采集159种木本植物的嫩叶,然后分别提取DNA信息;利用H-A、L和K标记进行PCR扩增和测序;将测序结果用MUSCLE软件进行比对[25],再用R软件中的“Phylotools”程序包[26]进行序列的拼接;最后用最大似然法对159物种建立系统发育树, 以自助法重复1 000次确定每个节点的支持率。Blomberg’s值基于系统发育树的布朗运动模型计算。=1是布朗运动模型的期望值,表明明显的系统发育保守性,而=0表明没有系统发育保守性[27],当0<<1时,系统发育保守性由实际和随机模拟的系统发育信号间差异的显著性决定。本研究使用R软件选取72种物种构建系统发育树。
1.6 数据分析
所研究的72种木本植物中记录有落叶时间的物种51种,记录有叶变色时间的物种69种。我们将时间转化为儒略日期(以每年的1月1日作为第1天,1月2日作为第2天,依此类推,将观测到的叶变色日期和落叶日期转化成距离当年1月1日的天数)与此同时,我们还将同一树种不同个体的天数算其平均值作为该树种最终落叶期或叶变色期(表1),此外,我们还将儒略日期重新换算成月份,从而记录每个月的叶变色和落叶树种数。
我们在分析古田山亚热带常绿阔叶林秋季叶物候的整体格局时,首先分析了木本植物的物候叶变色期与落叶期在时间序列上的分布格局。其次, 为了分析环境条件与秋季叶物候之间的关系,我们计算了古田山2018—2020年3年的每月平均温度和平均降水,我们还根据古田山的纬度计算了每月平均日照时长,然后将群落中每月叶变色和落叶的物种数与月均温、月均降水量和月均日照时数作多元线性回归分析,使用Akaike信息标准(AIC)优化模型,确定哪些环境因素对每月叶变色物种数和落叶物种数有影响[28]。同时,使用独立样本检验分析不同功能群树种的秋季叶变色和落叶物候期特征的差别。所有数据分析均在R软件中完成[29]。
2 结果和分析
2.1 气候因素的影响
古田山地处亚热带季风气候,季节特征分明, 气候变化明显,夏季高温多雨且日照时间长,冬季温和少雨且日照时间短(图1)。51种植物的落叶现象集中出现在每年的最后3个月,即10—12月, 其中2%的物种叶变色期集中在10月,75%在11月, 23%在12月。所有物种平均落叶日期为11月24日, 落叶最早的是浙闽樱桃(, 10月22日),最晚的是石楠(, 12月26日),相差65 d。
69种物种的叶变色现象集中出现在9—12月, 其中32%的物种集中在9月,64%在10月,3%在11月,1%在12月。所有物种叶变色平均日期为10月4日,叶变色最早的是浙闽樱桃(9月9日),最晚的是浙江新木姜子(, 12月5日),相差87 d。
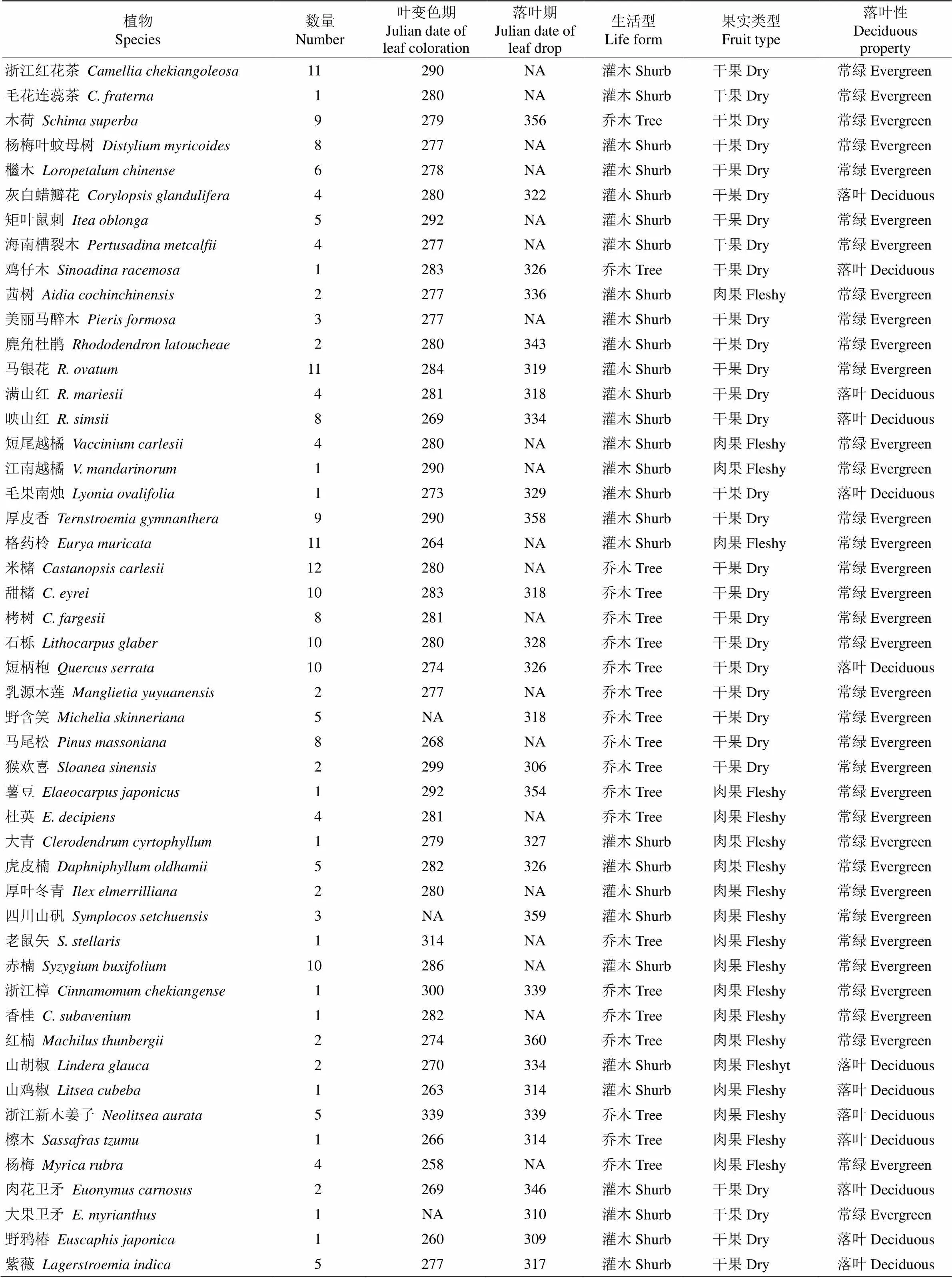
表1 浙江古田山72种木本植物的叶衰老期、功能性状
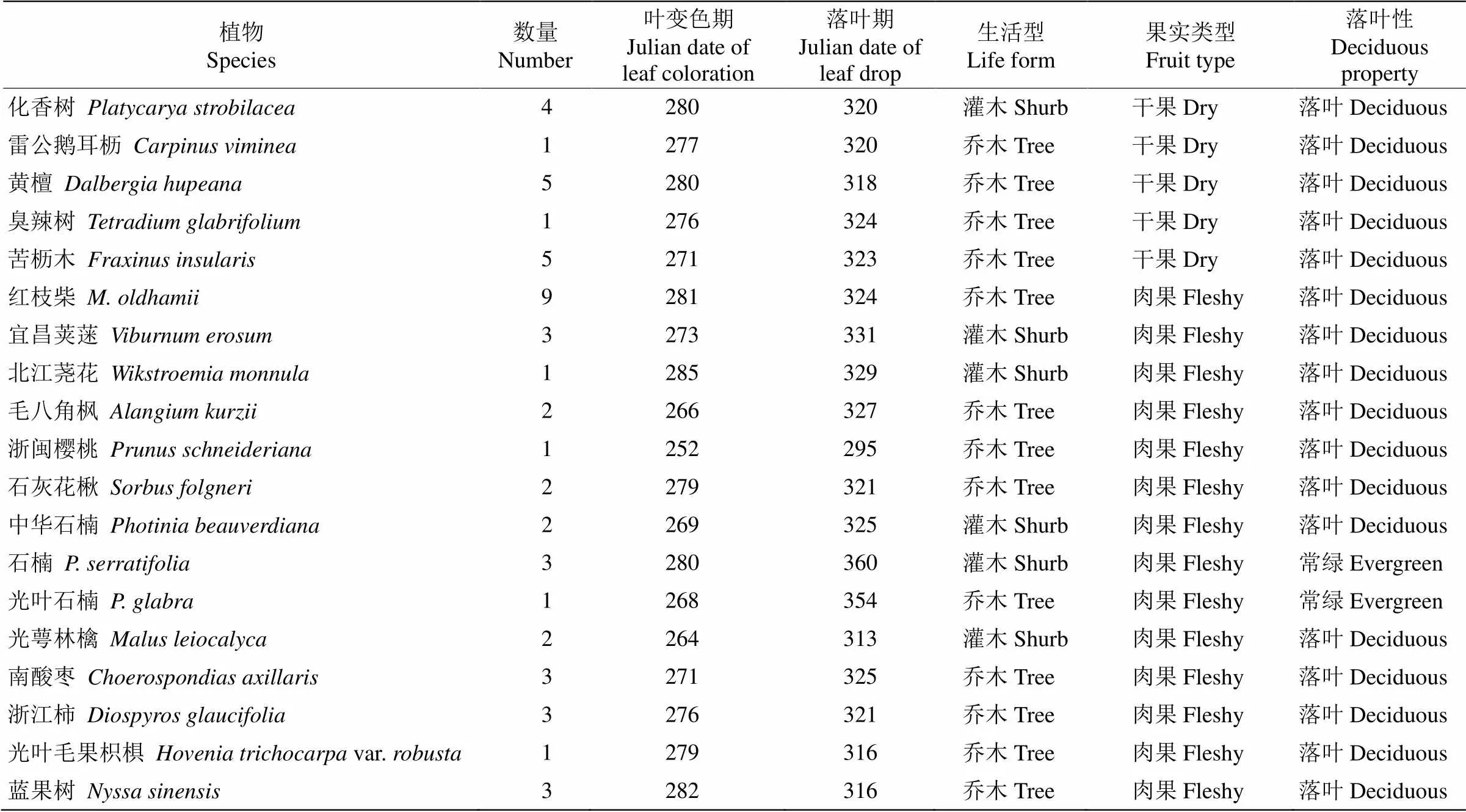
续表(Continued)
NA: 数据空缺。
NA: Missing data.
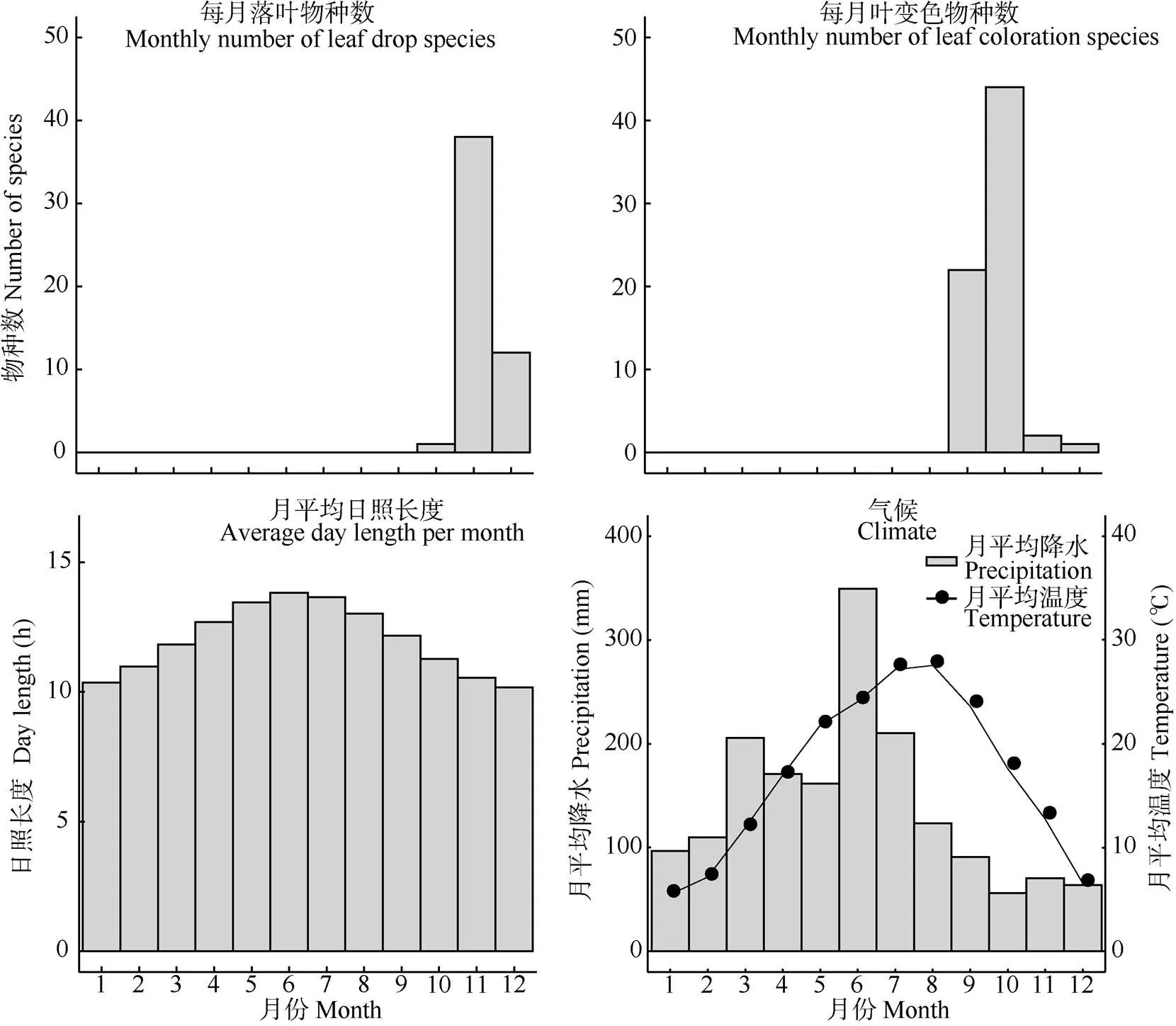
图1 古田山植物叶衰老物候和气候特征
古田山每月落叶植物种数和月均降水量、月均温和月均日照时数间没有显著相关性,每月叶变色植物种数和月均降水量没有显著相关性,但是每月叶变色植物种数与月均温(=0.060)和月均日照时数都呈弱相关性(=0.057, 表2)。
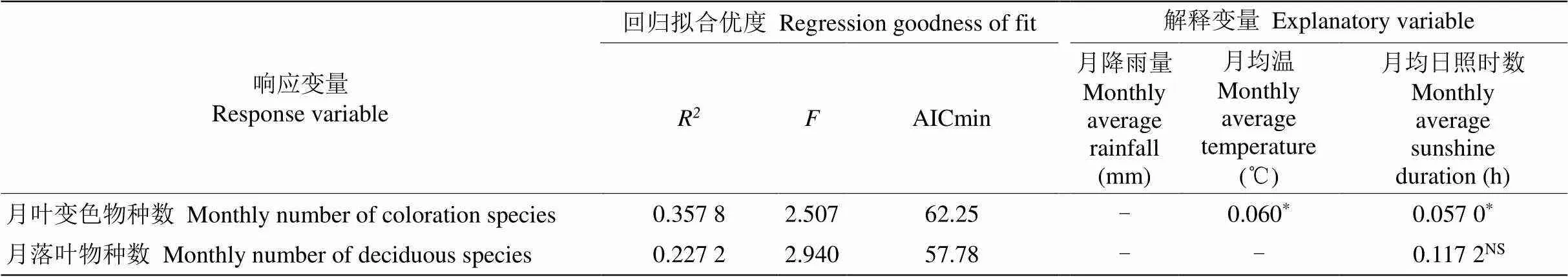
表2 气候因子和叶变色物种数或落叶物种数的多元回归分析
*: 0.05<<0.1; NS: 不显著; -: 不包含在模型中的变量。
*: 0.05<<0.1; NS: Not significant; -: Variable were not included in model.
2.2 谱系关系的影响
古田山亚热带常绿阔叶林群落中所研究的69物种的叶变色期谱系信号不显著(=0.338,=0.13); 51种物种的落叶期谱系信号也不显著(=0.256,= 0.689),表明所研究的物种亲缘关系的远近并不影响其叶变色期和落叶期。我们还将植物按照不同功能性状分别计算了Blomberg’s值,检测功能性状与物种间系统发育关系的相关性,结果都显示谱系信号不显著(表3)。
2.3 功能性状的影响
关于植物叶变色,植物落叶性对叶变色期具有显著影响,落叶树种叶变色期比常绿树种早7 d。但是木本植物的生活型和果实类型对叶变色时间都没有显著影响,植物的落叶性对其落叶期有显著影响。落叶树种落叶期比常绿树种早14 d。植物的生活型和果实类型对落叶期也没有显著影响(图2)。
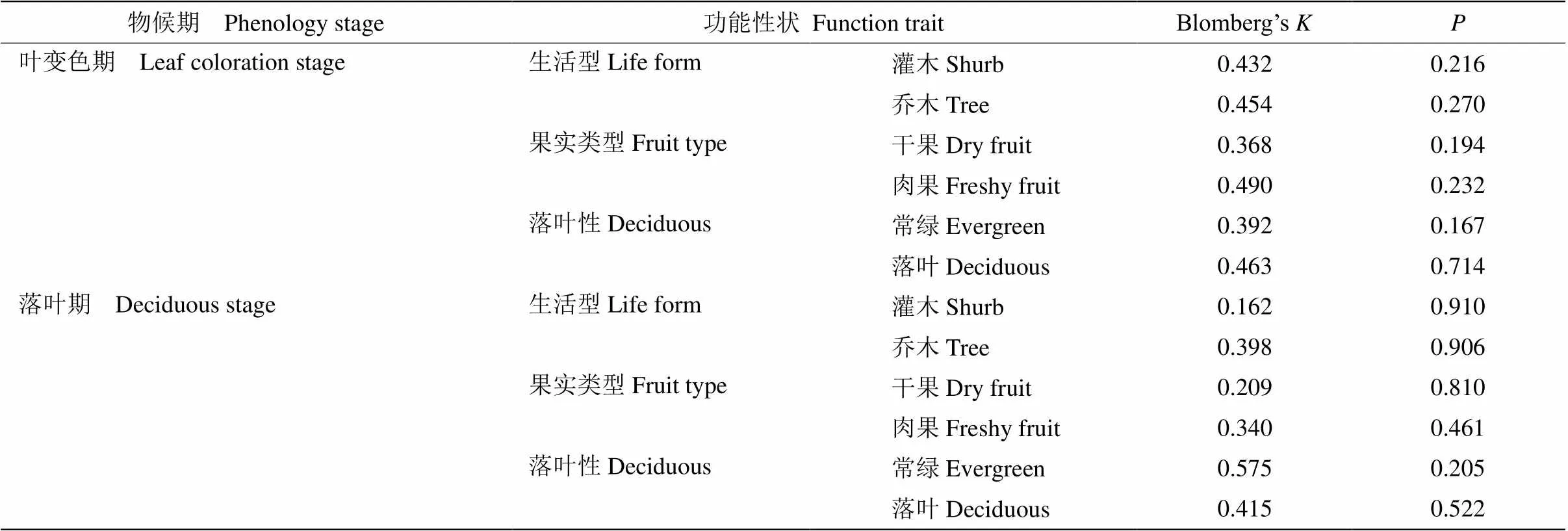
表3 古田山72种木本植物叶衰老物候的Blomberg’s K值
3 结论和讨论
3.1 气候因素对植物秋季叶物候的影响
本次研究的72种木本植物从群落水平上看叶变色现象集中出现在9—12月,即在亚热带植物叶变色开始日期主要集中在秋冬季节,与之前在温带的研究结果不同,温带落叶林植物叶衰老期(落叶与叶变色之和占该植物总叶片的50%)的开始日期集中在7—9月[14]。这可能是因为亚热带秋冬气候比温带更加温暖且主要分布树种是常绿树种,而温带地区落叶树种占主导地位,落叶树种较常绿树种对温度变化更为敏感,为了抵御不利环境,更早地做出响应。我们的研究表明每月叶变色的植物物种数和月平均降水量没有显著的相关性。这与Seyfert[30]的研究结果不同,他们认为土壤含水量的水平会影响植物叶变色的开始。然而,Estrella等[31]报道每月的降水总量和每月的干燥天数都与叶变色不相关, 供水可能不是叶变色的触发因素,这与本研究结果一致。
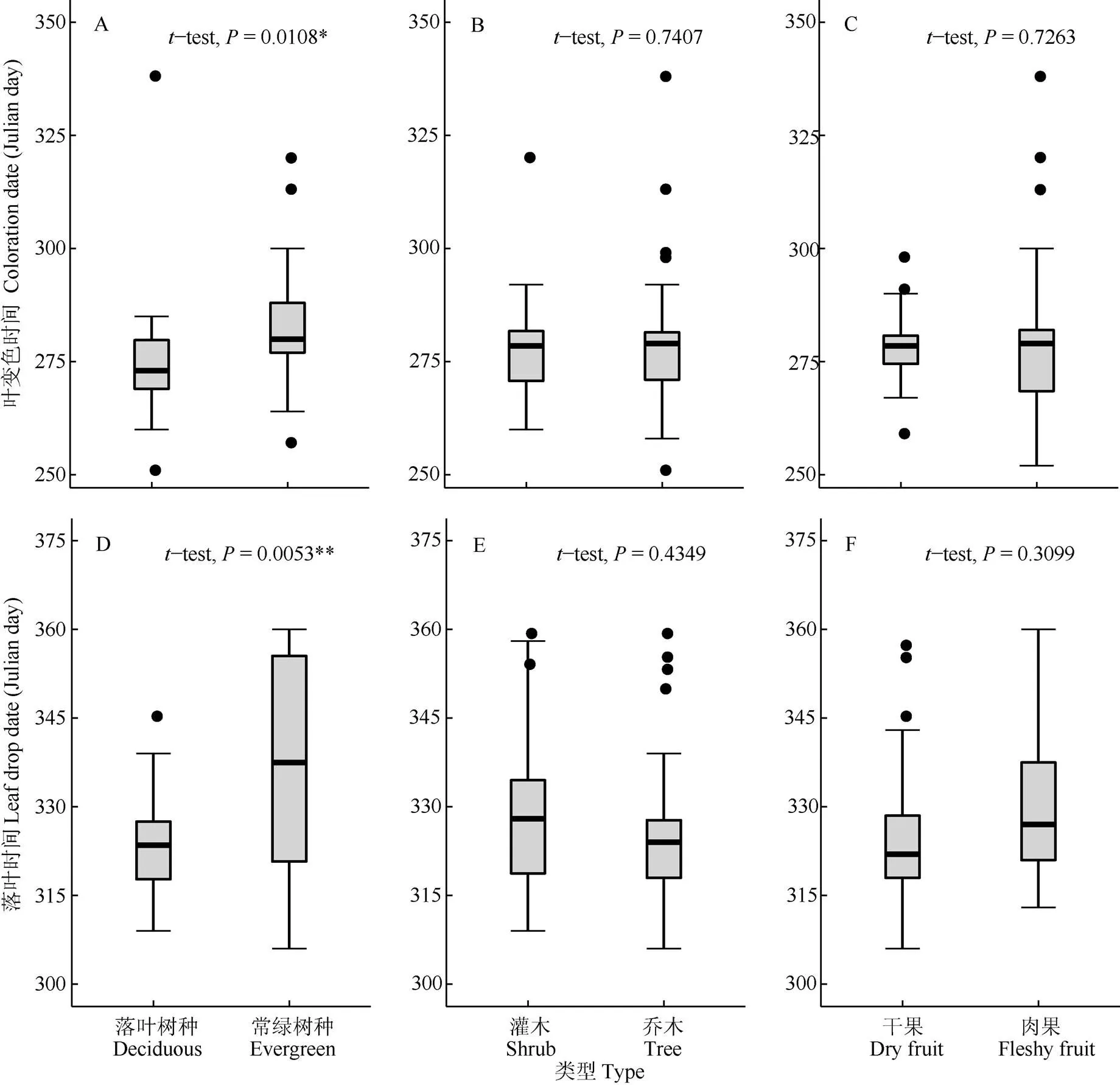
图2 不同功能类型植物的秋季叶物候差异
叶变色期和月平均温度以及月平均日照长度呈弱相关。在多年生树种中,温度和日照长度在树木叶衰老中起着关键作用[32]。橡树()、桦树()、山毛榉()等树木叶变色和温度相关[31]。Rosenthal等[33]报道光周期延长可以使落叶松针延迟变黄。此外,叶片变色或者落叶还与秋季温度迅速下降的变化日期和第一次霜冻的变化日期有关[31]。
古田山常绿阔叶林中落叶开始时间集中出现在10—12月,每月落叶植物物种数与月平均降水量、月平均温度呈弱相关和月平均日照长度都不相关。而前人[34–35]的研究表明植物落叶和温度成负相关,低温促进了大量的植物落叶,这与本研究结果不同。Bullock等[12]认为一些植物落叶受到日照时间的影响。然而,还有研究表明秋季物候事件几乎不受气温的影响[36],Vitasse等[37]报道欧亚槭()和欧梣()的叶衰老和温度没有显著关系,山毛榉和橡木()叶衰老日期的变异性并不是日照时间决定的。还有研究认为,降水可能不是物候的主要决定因素,树木本身的水分状况可能更加重要[38]。Williams- Linera[19]认为常绿树种的落叶与降雨量或者温度无关。在一些热带森林中,落叶和降水之间的相关性不显著[39]。法国圭亚那常绿林的常绿树种落叶具有季节性,但这种现象和降水以及干旱都不相关[39]。Xie等[40]的研究表明,只有在日均温为10 ℃时,落叶的开始时间和温度显著相关。树木衰老的影响因子依赖于物种,不同生理结构和生理机制的树种对环境变化表现出不同的应对策略[37],比如有的物种对温度敏感有的却不敏感[37]。现在关于秋季叶衰老决定因素的研究结论并不一致[30],对于触发叶衰老环境因子知之甚少[37]。此外,Xie等[40]认为,忽略地点间的区别可能会高估环境因素驱动的物候变化。
落叶期的开始和预先设定的3个环境因子都没有显著相关性,这可能是因为落叶期的开始是植物叶片组织采取了主动脱离的策略,以尽可能多的回收叶片中的资源。落叶作为树木衰老过程中的最后一步,其组织的脱离通常分为主动和被动两部分[41]。叶绿素的制造和维持对损害非常敏感,环境变化会直接影响植物的叶绿素细胞的数量和叶片营养物质的合成,当营养物质供应减少时,叶柄或叶柄基部的薄弱区开始发育,从而引起了植物落叶,叶片的主动脱落往往是伴随着叶变色进行的[41]。而叶片的被动脱落是指低温直接杀死了叶片细胞,从而引起的落叶,叶片一旦死亡,叶细胞拥有的所有资源都无法被树木重新吸收[41]。
3.2 系统发育对植物秋季叶物候的影响
在古田山亚热带常绿阔叶林群落中所研究物种的叶变色和落叶物候均与系统发育没有显著关系,这表明叶变色和落叶时间在系统发育上并不保守,即所研究的物种亲缘关系的远近并不影响其叶变色和落叶物候期。这与温带的木本植物叶片衰老时间结果一致,表明进化关系不是决定叶变色和落叶的重要因素,因此在统计分析中,系统发育不受控制[14]。值得注意的是,系统发育信号对亚热带展叶物候的影响非常显著[18]。对北美、欧洲和东亚木本植物的研究也表明物种的展叶物候在一定程度上取决于其系统发育位置,近缘种比远缘种更相似[42]。Desnoues等[43]在对温带树种展叶物候的研究中表明系统发育亲缘关系的相对重要性随分类水平的降低而增加,因此,来自近缘属或同属内的两物种在共同环境条件下生长时,更有可能共享相似的展叶期。这可能是物种的系统发育历史中固有的发育或结构限制的结果[44],此外,植物生理特征可能决定物种对特定环境线索的反应,因此,亲缘关系近的物种可能具有相似的生理特征和敏感性[45]。目前对植物叶物候的研究表明展叶物候的系统发育是保守,而落叶和叶变色无论是温带还是亚热带常绿林都是不保守的,但是这一现象是否普遍,还有待更多案例的进一步佐证。
3.3 功能性状对植物秋季叶物候的影响
本研究中,植物的落叶类型对植物叶变色开始时间和落叶开始时间都有显著影响,落叶树种平均叶变色时间和落叶时间比常绿树种早。常绿和落叶这2种习性在本质上是因为叶结构特征不同导致的2种不同的生活史策略[46–47]。有研究表明,落叶物种的叶片与小枝的水平倾角低,节间距长,单位叶面积干重小,光截取能级显著大于常绿树种[46–47]。落叶树种叶寿命短,干旱季节或者寒冷季节来临前所有叶片都会凋落[46]。相反,常绿物种单位叶面积干重较大,其光合能力较弱,因此为了在时间上弥补碳同化量,常绿物种比落叶物种有更长的碳获取时间[47],叶寿命也就更长。因此常绿树种叶变色和落叶时间都比落叶树种晚。落叶植物的生命周期包括生长期和休眠期,这种休眠保护落叶植物免受极端冬季天气的影响,在秋冬温度较低,气候较为干燥的时候,树木不是花费精力来保护这些脆弱的器官,而是通过落叶来保护资源[48]。所以落叶植物有可能会对气候变化更为敏感,当环境变得不利于其生存,树木会提前进入落叶期进行自我保护。在一项对7种树种的调查中发现落叶树种比常绿树种展叶时间更早[49],因此落叶树种落叶早于常绿树种也可能是权衡碳氮代谢冲突的最佳植物策略[50]。
同时,本研究中,植物的生活型对植物叶变色和落叶并没有显著影响,这与在弗雷明汉植物园、渥太华植物园、北京植物园和莫顿植物园的研究结果一致,树木、灌木和藤本植物的叶片衰老日期没有显著差异[18]。在对植物的展叶研究中,林下树种和树苗为了更好地利用冠层关闭前的可用光而更早展叶[6–7],灌木、乔木和藤本植物间的差异显著,特别是灌木平均比乔木早10 d[18]。同时,植物的果实类型和叶变色期、落叶期都不相关,而杨晓东等[15]对新疆乌鲁木齐开花物候的研究表明,与干果相比,肉质果实开花更早,这可能是因为开花和结果都属于繁殖物候,所以果实类型对开花物候的影响比叶衰老物候更大。
本研究通过记录叶变色开始日期和落叶开始日期探究亚热带木本植物秋季物候格局及其驱动因子,认为植物落叶和叶变色发生时间主要集中在秋冬季节,并且气候因子、植物功能性状等内外因素都一定程度上会对常绿阔叶林木本植物的秋季物候造成影响,其中植物落叶受环境的影响较叶变色更为显著。就内在因素而言,植物功能型较植物间系统发育关系对其秋季物候有更明显的影响。本次研究有助于我们了解亚热带植物秋季物候的影响因素,亚热带植物叶衰老系统发育不保守也有待更多的研究。植物的叶衰老影响了许多生态系统过程,包括碳、水和养分循环,了解特定地点和整个物种范围内物种叶片衰老时间的变化及其影响因素,为探讨植物对环境的适应机制具有重要的意义。
[1] ZHU K Z, WAN M W. A productive science: Phenology [J]. Pop Sci, 1963(1): 6–8. [竺可桢, 宛敏渭. 一门丰产的科学——物候学[J]. 科学大众, 1963(1): 6–8.]
[2] WAND Z J, ZHANG D Y, ZHUO L. Relationship of phenology inBunge and temperature variation in southern Junggar Basin [J]. Nonwood For Res, 2013, 31(1): 102–105. [汪智军, 张东亚, 卓立. 准噶尔盆地南缘文冠果物候与气温变化的关系[J]. 经济林研究, 2013, 31(1): 102–105. doi: 10.14067/j.cnki.1003-8981. 2013.01.010.]
[3] AHAS R. Long-term phyto-, ornitho- and ichthyophenological time- series analyses in Estonia [J]. Int J Biometeorol, 1999, 42(3): 119–123. doi: 10.1007/s004840050094.
[4] Cleland E E, Chuine I, Menzel A, et al. Shifting plant phenol- logy in response to global change [J]. Trends Ecol Evol, 2007, 22(7): 357–365. doi: 10.1016/j.tree.2007.04.003.
[5] Polgar C A, Primack R B. Translated by DU Y J. Leaf out phenology in temperate forests [J]. Biodiv Sci, 2013, 21(1): 111–116. [POLGAR C A, PRIMACK R B. 杜彦君. 译. 温带森林展叶物候学 [J]. 生物多样性, 2013, 21(1): 111–116. doi: 10.3724/SP.J.1003.2013. 06187.
[6] AUGSPURGER C K, BARTLETT E A. Differences in leaf phenology between juvenile and adult trees in a temperate deciduous forest [J]. Tree Physiol, 2003, 23(8): 517–525. doi: 10.1093/treephys/23.8.517.
[7] SEIWA K. Changes in leaf phenology are dependent on tree height in, a deciduous broad-leaved tree [J]. Ann Bot, 1999, 83(4): 355–361. doi: 10.1006/anbo.1998.0831.
[8] LIN Y N, LIN X. Phenological observation of five Liana medicinal plants [J]. Territ Nat Resour Study, 1997(2): 63–64. [林月男, 林湘. 五种藤本药用植物物候观察[J]. 国土与自然资源研究, 1997(2): 63– 64. doi: 10.16202/j.cnki.tnrs.1997.02.015.]
[9] ZANI D, CROWTHER T W, MO L D, et al. Increased growing-season productivity drives earlier autumn leaf senescence in temperate trees [J]. Science, 2020, 370(6520): 1066–1071. doi: 10.1126/science.abd 8911.
[10] FRIEDMAN J M, ROELLE J E, CADE B S. Genetic and environ- mental influences on leaf phenology and cold hardiness of native and introduced riparian trees [J]. Int J Biometeorol, 2011, 55(6): 775–787. doi: 10.1007/s00484-011-0494-6.
[11] Nanda A, Suresh H S, Krishnamurthy Y L. Phenology of a tropical dry deciduous forest of Bhadra Wildlife Sanctuary, southern India [J]. Ecol Process, 2014, 3(1): 1. doi: 10.1186/2192-1709-3-1.
[12] BULLOCK S H, SOLIS-MAGALLANES J A. Phenology of canopy trees of a tropical deciduous forest in Mexico [J]. Biotropica, 1990, 22(1): 22–35. doi: 10.2307/2388716.
[13] WU C Y, WANG X Y, WANG H J, et al. Contrasting responses of autumn-leaf senescence to daytime and night-time warming [J]. Nat Clim Change, 2018, 8(12): 1092–1096. doi: 10.1038/s41558-018-034 6-z.
[14] Panchen Z A, Primack R B, Gallinat A S, et al. Substantial variation in leaf senescence times among 1360 temperate woody plant species: Implications for phenology and ecosystem processes [J]. Ann Bot, 2015, 116(6): 865–873. doi: 10.1093/aob/mcv015.
[15] WANG Y, YANG X D, ALI A, et al. Flowering phenology shifts in response to functional traits, growth form, and phylogeny of woody species in a desert area [J]. Front Plant Sci, 2020, 11: 536. doi: 10. 3389/fpls.2020.00536.
[16] DU Y J, MI X C, LIU X J, et al. Seed dispersal phenology and dispersal syndromes in a subtropical broad-leaved forest of China [J]. For Ecol Manage, 2009, 258(7): 1147–1152. doi: 10.1016/j.foreco. 2009.06.004.
[17] Legendre P, Mi X C, Ren H B, et al. Partitioning beta diversity in a subtropical broad-leaved forest of china [J]. Ecology, 2009, 90(3): 663–674. doi: 10.1890/07-1880.1.
[18] Panchen Z A, Primack R B, Nordt B, et al. Leaf out times of temperate woody plants are related to phylogeny, deciduousness, growth habit and wood anatomy [J]. New Phytol, 2014, 203(4): 1208– 1219. doi: 10.1111/nph.12892.
[19] Williams-Linera G. Phenology of deciduous and broadleaved- evergreen tree species in a Mexican tropical lower montane forest [J]. Glob Ecol Biogeogr Lett, 1997, 6(2): 115–127. doi: 10.2307/2997568.
[20] HARRIS I, OSBORN T J, JONES P, et al. Version 4 of the CRU TS monthly high-resolution gridded multivariate climate dataset [J]. Sci Data, 2020, 7(1): 109. doi: 10.1038/s41597-020-0453-3.
[21] HU X L, CHANG Z Y, DU Y J. Effects of pollination mode and fruit type on reproductive phenology of woody plants [J]. Guihaia, 2017, 37(3): 315–321. [胡小丽, 常朝阳, 杜彦君. 传粉方式和果实类型对木本植物繁殖物候的影响[J]. 广西植物, 2017, 37(3): 315–321. doi: 10.11931/guihaia.gxzw201605001.]
[22] Zhu Y, Zhao G F, Zhang L W, et al. Community composition and structure of Gutianshan forest dynamic plot in a mid-subtropical evergreen broad-leaved forest, east China [J]. Chin J Plant Ecol, 2008, 32(2): 262–273. [祝燕, 赵谷风, 张俪文, 等. 古田山中亚热带常绿阔叶林动态监测样地——群落组成与结构[J]. 植物生态学报, 2008, 32(2): 262–273. doi: 10.3773/j.issn.1005-264x.2008.02.004.]
[23] HU X L, ZHANG Y J H, MI X C, et al. Influence of climate, phylo- geny, and functional traits on flowering phenology in a subtropical evergreen broad-leaved forest, east China [J]. Biodiv Sci, 2015, 23(5): 601–609. [胡小丽, 张杨家豪, 米湘成, 等. 浙江古田山亚热带常绿阔叶林开花物候: 气候因素、系统发育关系和功能性状的影响[J]. 生物多样性, 2015, 23(5): 601–609. doi: 10.17520/biods.2015083.
[24] Kress W J, Erickson D L, Jones F A, et al. Plant DNA barcodes and a community phylogeny of a tropical forest dynamics plot in panama [J]. Proc Natl Acad Sci USA, 2009, 106(44): 18621–18626. doi: 10.1073/pnas.0909820106.
[25] Edgar R C. MUSCLE: Multiple sequence alignment with high accu- racy and high throughput [J]. Nucl Acids Res, 2004, 32(5): 1792–1797. doi: 10.1093/nar/gkh340.
[26] Zhang J L, Mi X C, Pei N C. Phylotools: Phylogenetic tools for ecologists. R package version 0.0 [CP]. 2010, 7: 201019.
[27] DAVIES T J, WOLKOVICH E M, KRAFT N J B, et al. Phylogenetic conservatism in plant phenology [J]. J Ecol, 2013, 101(6): 1520–1530. doi: 10.1111/1365-2745.12154.
[28] AKAIKE H. Information theory and an extension of the maximum likelihood principle [M]// PARZEN E, TANABE K, KITAGAWA G. Selected Papers of Hirotugu Akaike. New York: Springer, 1998: 199– 213. doi: 10.1007/978-1-4612-1694-0_15.
[29] R Core Team. R: A Language and Environment for Statistical Computing [CP]. Vienna, Austria: R Foundation for Statistical Computing, 2021. https://www.R-project.org/.
[30] SEYFERT F. Eintritt phänologischer phasen und auslösende wetter- lagen [J]. Angew Meteorol, 1955, 2(6): 183–190.
[31] ESTRELLA N, MENZEL A. Responses of leaf colouring in four deciduous tree species to climate and weather in Germany [J]. Climate Res, 2006, 32(3): 253–267. doi: 10.3354/cr032253.
[32] LEE D W, O’KEEFE J, HOLBROOK N M, et al. Pigment dynamics and autumn leaf senescence in a New England deciduous forest, eastern USA [J]. Ecol Res, 2003, 18(6): 677–694. doi: 10.1111/j.1440- 1703.2003.00588.x.
[33] ROSENTHAL S I, CAMM E L. Photosynthetic decline and pigment loss during autumn foliar senescence in western larch () [J]. Tree Physiol, 1997, 17(12): 767–775. doi: 10.1093/tree phys/17.12.767.
[34] Wu Y, Liu W Y, Shen Y X, et al. A preliminary study on phenology of indigenous plants in karst mountainous region at the stone forest scenic spot in Yunnan [J]. Mount Res, 2006, 24(6): 647–653. [吴毅, 刘文耀, 沈有信, 等. 云南石林景区主要乡土植物物候特征的初步研究[J]. 山地学报, 2006, 24(6): 647–653. doi: 10.3969/j.issn.1008-2786. 2006.06.002.]
[35] Chen F J, Chen K H, Xie Y G, et al. Phenological patterns of main plants in Karst ecosystem in northwest Guizhou, China [J]. Mount Res, 2010, 28(6): 695–703. [陈发军, 陈坤浩, 谢永贵, 等. 黔西北喀斯特生态系统中主要植物物候格局[J]. 山地学报, 2010, 28(6): 695–703. doi: 10.16089/j.cnki. 1008-2786.2010.06.002.]
[36] SPARKS T H, MENZEL A. Observed changes in seasons: An over- view [J]. Int J Climatol, 2002, 22(14): 1715–1725. doi: 10.1002/joc. 821.
[37] VITASSE Y, PORTÉ A J, KREMER A, et al. Responses of canopy duration to temperature changes in four temperate tree species: Rela- tive contributions of spring and autumn leaf phenology [J]. Oecologia, 2009, 161(1): 187–198. doi: 10.1007/s00442-009-1363-4.
[38] BORCHERT R. Soil and stem water storage determine phenology and distribution of tropical dry forest trees [J]. Ecology, 1994, 75(5): 1437– 1449. doi: 10.2307/1937467.
[39] LOUBRY D. Phenology of deciduous trees in a French-Guianan forest (5 degrees latitude north): Case of a determinism with endogenous and exogenous components [J]. Can J Bot, 1994, 72(12): 1843–1857.
[40] XIE Y Y, WANG X J, WILSON A M, et al. Predicting autumn phenol- logy: How deciduous tree species respond to weather stressors [J]. Agric For Meteorol, 2018, 250/251: 127–137. doi: 10.1016/j.agrformet. 2017.12.259.
[41] Coder K D. Leaf senescence & abscission [M/OL]. Athens: University of Georgia, (2008-9-12). https://hdl.handle.net/10724/36467.
[42] Zohner C M, Benito B M, Fridley J D, et al. Spring predict- tability explains different leaf-out strategies in the woody floras of North America, Europe and East Asia [J]. Ecol Lett, 2017, 20(4): 452– 460. doi: 10.1111/ele.12746.
[43] Desnoues E, de Carvalho J F, Zohner C M, et al. The relative roles of local climate adaptation and phylogeny in determining leaf-out timing of temperate tree species [J]. For Ecosyst, 2017, 4(1): 26. doi: 10.1186/s40663-017-0113-z.
[44] Lechowicz M J. Why do temperate deciduous trees leaf out at different times? Adaptation and ecology of forest communities [J]. Am Nat, 1984, 124(6): 821–842. doi: 10.1086/284319.
[45] Harvey P H, Pagel M D. The Comparative Method in Evolutionary Biology [M]. Oxford: Oxford University Press, 1991: 1–248.
[46] Givnish T J. Adaptive significance of evergreen vs. deciduous leaves: Solving the triple paradox [J]. Silv Fenn, 2002, 36(3): 703–743. doi: 10.14214/sf.535.
[47] Souza J P, Prado C H b a, Albino A L S, et al. Shoot-foliage relationships in deciduous, semideciduous, and evergreen cerrado tree species [J]. Braz J Plant Physiol, 2009, 21(1): 76–86. doi: 10.1590/ S1677-04202009000100009.
[48] Zhang L, Luo TX. Advances in ecological studies on leaf lifespan and associated leaf traits [J]. Chin J Plant Ecol2004, 28(6): 844–852. [张林, 罗天祥. 植物叶寿命及其相关叶性状的生态学研究进展[J]. 植物生态学报, 2004, 28(6): 844–852.]
[49] Davi H, Gillmann M, Ibanez T, et al. Diversity of leaf unfolding dynamics among tree species: New insights from a study along an altitudinal gradient [J]. Agric For Meteorol, 2011, 151(12): 1504–1513. doi: 10.1016/j.agrformet.2011.06.008.
[50] Ingvarsson P K, García M V, Hall D, et al. Clinal variation in, a candidate gene for day-length-induced growth cessation and bud set, across a latitudinal gradient in European aspen () [J]. Genetics, 2006, 172(3): 1845–1853. doi: 10.1534/genetics. 105.047522.
Studies on Affecting Factors of Leaf Senescence Phenology in a Subtropical Evergreen Broad-leaved Forest in Gutianshan, Zhejiang Province
SHOU Jiajun, XUE Qianhuai, WANG Xinyang, TANG Xinran, CHEN Qin, DU Yanjun*
(College of forestry, Hainan University, Haikou 570100, China)
In order to explore the influencing factors of leaf senescence phenology in subtropical evergreen broad-leaved forest, the phylogenetic relationships, functional traits of 72 woody plants and climate factors were studied in Gutianshan subtropical evergreen broad-leaved forest in Zhejiang Province from November 2018 to February 2021. The results showed that the leaf coloration period was from September to December, and the leaf drop period was from October to December. There was no significant correlation between the number of leaf drop species and monthly average temperature, monthly average precipitation and monthly average sunshine duration. The monthly number of leaf coloration species had weak correlation with monthly average temperature and monthly average sunshine duration. The deciduous property had a significant effect on leaf coloration and leaf drop stages. The phylogenetic relationship among species had no significant effect on leaf coloration and leaf drop stages. Therefore, both biotic and abiotic factors could affect leaf senescence for evergreen broad-leaved species, which had an important value for improving the prediction models of autumn phenology.
Leaf coloration phenology; Subtropical evergreen broad-leaved forest; Leaf drop; Functional trait; Phylogenetic conservatism
10.11926/jtsb.4582
2021-11-25
2022-03-11
海南省研究生创新科研课题(Qhys2021-120)资助
This work was supported by the Project for Innovative Research of Graduate Students in Hainan (Grant No. Qhys2021-120).
寿佳君(1996年生),女,硕士研究生,研究方向为植物物候。E-mail: maia7782@163.com
通讯作者 Corresponding author.E-mail: yanjundu1981@gmail.com
猜你喜欢
幼儿画刊(2023年12期)2024-01-15 07:06:12
热带作物学报(2022年6期)2022-07-08 06:45:10
亚热带农业研究(2019年2期)2019-09-05 09:39:06
星星·诗歌原创(2017年5期)2017-08-13 10:09:55
军营文化天地(2017年6期)2017-06-28 11:30:29
西南农业学报(2017年5期)2017-06-23 08:14:35
河北林业科技(2016年5期)2016-11-08 03:12:55
军营文化天地(2016年3期)2016-05-26 06:45:33
西南农业学报(2016年5期)2016-05-17 05:42:22
海峡姐妹(2016年1期)2016-02-27 15:15:12
