
多功能木聚糖水解酶的研究及其在食品领域的潜在应用
2023-03-09 04:27王钰璐于海燕辛凤姣
食品与生物技术学报 2023年1期
韩 雪,王钰璐,于海燕,辛凤姣
(中国农业科学院 农产品加工研究所,北京 100193)
木质纤维素是一种重要的可再生生物质资源,因其清洁性及储量丰富的特点已成为潜在的规模化生产生物燃料及高附加值生物基产品的原料[1]。但是,天然木质纤维素的化学成分及结构都十分复杂,需利用多种糖类水解酶共同作用,才能对其进行有效的生物降解[2]。木质纤维素生物质由3 种主要成分组成,即纤维素(质量分数40%~60%)、半纤维素(质量分数20%~40%)和木质素(质量分数10%~25%)等[3]。半纤维素是一组非纤维素异质多糖的总称,由戊糖(木糖和阿拉伯糖)、己糖(甘露糖、葡萄糖和半乳糖)和糖酸(甲基葡糖醛酸)等组成[4]。根据其主链中存在的主要单糖进行分类,包括木聚糖、木葡聚糖、甘露聚糖、葡甘聚糖以及β-葡聚糖等[5]。
木聚糖是半纤维素中含量最丰富的成分,相对而言利用率较低,因此其作为可持续产品开发的重要生物资源而备受关注[6]。木聚糖广泛存在于秸秆、麦麸、玉米芯、甘蔗渣、果蔬等农副产品加工残余物中。木聚糖酶解产生木糖、木寡糖、阿拉伯糖、阿魏酸等多种生理活性成分,广泛应用于功能性食品、饮料、烘焙等食品行业中。并且,以木糖等酶解产物为前体可进一步合成木糖醇、阿拉伯糖醇、香兰素等食品添加剂[7]。木聚糖大多为异质结构,由β-D-吡喃木糖基组成主链,侧链含有不同类型的取代基。因此,木聚糖高效降解往往需要多种水解酶的协同作用。其中,内切木聚糖酶通过随机切割木聚糖内部的木糖糖苷键来攻击主链结构,其产物木寡糖被β-木糖苷酶进一步水解生成木糖单体。此外,木聚糖复杂的侧链组成需要额外的侧链水解酶,如α-L-阿拉伯呋喃糖苷酶、乙酰木聚糖酯酶等。然而,木聚糖水解过程中酶的种类和用量的高需求及降解方式的低效率导致多种酶投放成本较高,这也是木聚糖高效生物转化的主要瓶颈[8]。多功能酶在一条多肽链中同时具备多种活性,具有“一酶多能”的特点。木聚糖酶解体系中多功能酶的应用是降低成本的有效策略,此外,多功能水解酶在降解木聚糖复杂底物方面具有潜在的协同作用,因此具有广阔的应用前景[9]。作者将首先介绍木聚糖的结构组成及水解酶系,进而主要从酶的结构与功能方面系统综述多功能木聚糖水解酶的研究进展及其在食品领域中潜在的应用,最后展望该领域的发展前景。
1 木聚糖的结构组成及水解酶系
1.1 木聚糖的结构组成
木聚糖是一种复杂的异质性多糖,根据木聚糖侧链糖残基的类型不同分为:同型木聚糖、葡糖醛酸木聚糖、阿拉伯木聚糖、阿拉伯葡糖醛酸木聚糖和葡糖醛酸阿拉伯木聚糖(见图1)[10]。同型木聚糖是一种由木糖残基组成的非支化木聚糖,根据糖苷键的不同分为β-1,4-木聚糖、β-1,3-木聚糖、混合型β-1,3/1,4-木聚糖,广泛存在于绿藻等植物中[11];葡糖醛酸木聚糖是含有α-1,2 连接的葡糖醛酸基和4-O-甲基葡糖醛酸基取代的木聚糖,主要存在于双子叶植物中[12];阿拉伯木聚糖是指D-木糖残基的C2/C3 位连有L-阿拉伯呋喃糖取代基的β-1,4-木聚糖,主要存在于谷物中[13];其他两种类型的木聚糖,即阿拉伯葡糖醛酸木聚糖和葡糖醛酸阿拉伯木聚糖,含有甲基葡糖醛酸残基和L-阿拉伯呋喃糖残基,分别连接在主链β-1,4-木糖残基的C2 和C3位置,这两个术语有时可以互换,但L-阿拉伯呋喃糖与糖醛酸的比例较低的聚合物常被称为葡糖醛酸阿拉伯木聚糖[14]。此外,木聚糖中的L-阿拉伯呋喃糖基单元可以进一步在C5 位被羟基肉桂酸酯(即阿魏酸和对香豆酸)取代[13]。

图1 木聚糖的结构组成Fig.1 Structural composition of xylan
1.2 木聚糖水解酶系
木聚糖完全水解需要多种酶协同发挥作用,包括主链水解酶β-1,4-内切木聚糖酶、β-D-木糖苷酶、还原端外切木聚糖酶,以及侧链水解酶α-L-阿拉伯呋喃糖苷酶、α-D-葡糖醛酸酶、乙酰木聚糖酯酶、阿魏酸酯酶等。由于木聚糖水解酶成员的分类、催化位点、作用机制等特征已有相关文献进行总结[6,15-17],在此不逐一赘述。木聚糖水解酶系存在于两个家族:糖苷水解酶(GH)家族和碳水化合物酯酶(CE)家族(见表1)[16,18-24]。
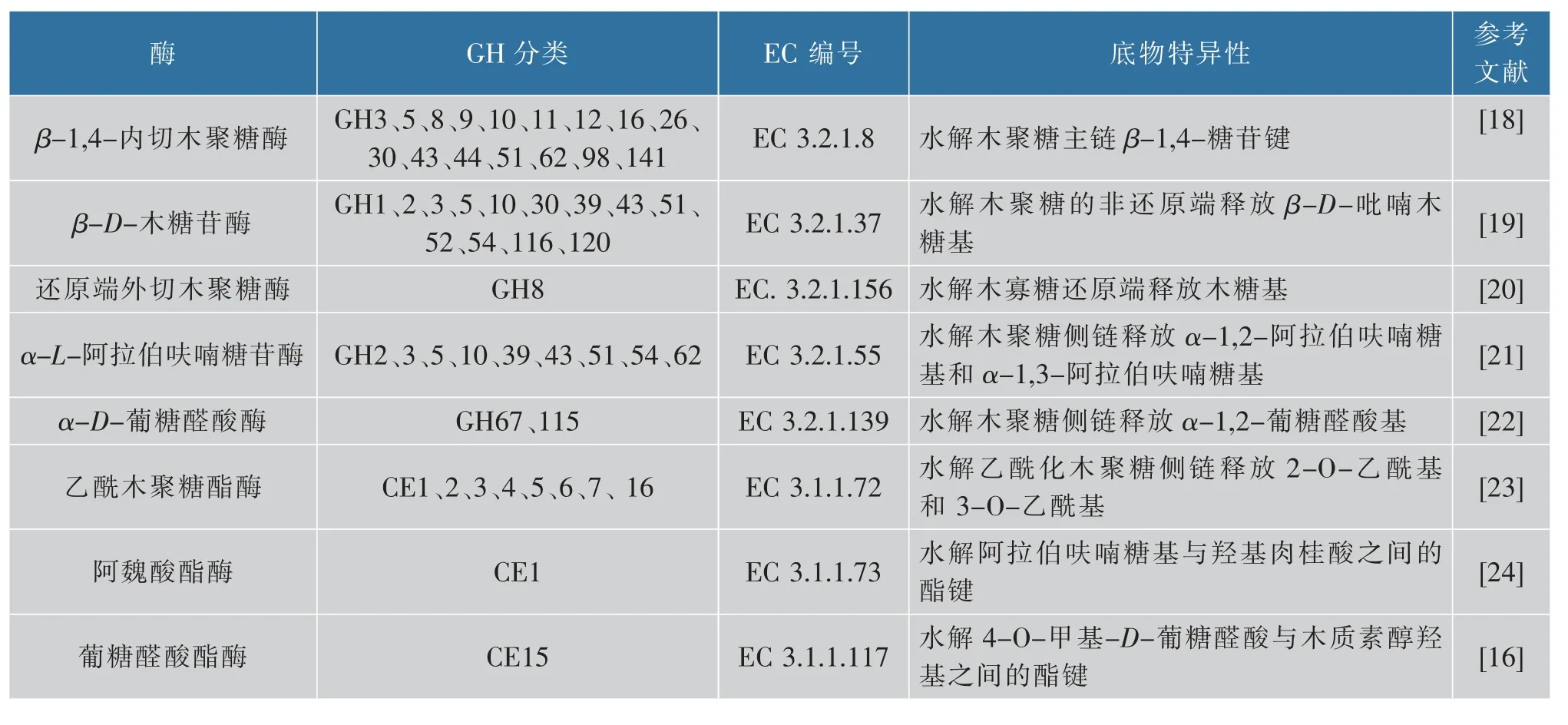
表1 木聚糖水解酶成员及其底物特异性Table 1 Xylanolytic enzyme members and their substrate specificity
2 多功能木聚糖水解酶
2.1 多功能木聚糖水解酶概述
多功能酶在酶蛋白的一条多肽链中同时具有两种或两种以上催化活性,可催化复杂的连续反应或多底物反应体系。在催化过程中,可能会有多个中间状态存在,这使得多功能酶表现出更高催化效率的特性。首先,第一个活性位点的产物可以主动或被动地直接通过底物通道转移到另一个活性中心,通道的“门控”作用有助于维持多个活性部位反应的精确同步,以最大限度地减少中间体的积累;其次,第一个反应的活性位点的状态可能影响第二个反应的活性位点的动力学特性,反之亦然[25-26]。同时,多功能酶往往具有良好的调控性,能够在不同催化条件下表现出不同的活性。多功能酶在进化上是由编码功能相关的基因融合而形成。推测复杂的多功能酶是由简单的单功能酶进化而来,催化活性相关的基因可以由共同的调控元件控制[27-28]。
木聚糖是一种复杂的异质性多糖,其多种侧链取代基的存在会形成空间位阻,使木聚糖难以被完全降解且酶解效率较低。因此,需要多酶协同催化来消除侧链的阻碍作用,使木聚糖主链更容易接近木聚糖酶的活性中心,进而提高木聚糖的降解效率[8,29]。然而,不同单酶的催化特性之间存在差异,且多酶催化体系在实际应用中往往面临着成本高、工艺复杂、重复利用率低等诸多挑战,因此,多功能酶对于低成本、高效木聚糖酶解体系的构建具有重要意义。根据结构域与催化功能的特点,木聚糖水解酶系主要分为两类:1)单催化结构域的多功能木聚糖水解酶,一条多肽链上仅有一个催化结构域,可通过一个或多个活性中心/位点催化不同的反应进而发挥多种作用,部分包含其他的辅助结构域(如碳水化合物结合模块[CBM]、纤维连接蛋白III[Fn3]等)对催化结构域的功能起调控作用;2)多催化结构域的多功能木聚糖水解酶,一条多肽链上存在多个催化结构域模块,各自具有不同的活性中心,不同结构域之间由柔性连接区(linker)或其他非催化结构域相连(见图2)。

图2 多功能木聚糖水解酶分类Fig.2 Classification of multifunctional xylanolytic enzyme
2.2 单催化结构域的多功能木聚糖水解酶
这一类多功能木聚糖水解酶仅含有一个独立的催化结构域催化不同的底物反应。根据其活性位点的位置,进一步可细分为单活性位点型、多活性位点型多功能木聚糖水解酶。
2.2.1 单活性位点型多功能木聚糖水解酶 单活性位点型多功能木聚糖水解酶在单一结构域中具有底物识别的泛杂性(promiscuity),通过相同的活性中心残基催化不同的底物。根据CAZy 数据库分析,具有多种底物特异性的木聚糖水解酶家族主要包括GH3、5、7、9、10、11、13、30、31、43、44、48 等。例如,GH30 家族的多功能木聚糖水解酶来自亚家族7,包括内切葡糖醛酸木聚糖酶和外切木二糖水解酶。该类酶的双功能分子机制已得到解析。Nakamichi 等报道解纤维素蓝状菌(Talaromyces cellulolyticus)来源的TcXyn30B 能够有效地将葡糖醛酸木聚糖转化为低聚木糖,进一步在低聚木糖的非还原端切割释放木糖单体[30-31]。与GH30 亚家族8来源的单功能葡糖醛酸木聚糖酶结构相比,TcXyn30B(PDB:6IUJ)中保守的Arg46 用于识别葡糖醛酸基侧链,对于内切木聚糖酶的活性十分关键;而活性中心附近的额外插入片段β2~α2 柔性环(loop)上的Asn93 用于识别低聚木糖的非还原端,对于外切木二糖水解酶活性很重要(见图3)。类似地,Nikolaivits 等解析 了嗜热 毁丝霉 菌(Thermothelomyces thermophila)来源的TtXyn30A及与产物复合体(PDB:7O0E)的晶体结构,该酶含有GH30 家族典型的TIM 桶型催化结构域和一个由9 条β 链组成的较小结构域(β9 结构域)[32]。β9 结构域中的第一条β 链位于催化结构域的N 端,而其余8 条β 链位于C 端,表明该结构域对于催化结构域的稳定折叠十分关键。生化分析表明该结构域不能与所测定的多糖结合,其功能还有待阐明。与TcXyn30B 相比,TtXyn30A 中同样存在额外的插入片段,对于木二糖水解酶活性很重要,而保守的Arg34 尽管对酶活力很重要,但并不直接介导与葡糖醛酸基侧链的识别,这与其他已报道的GH30 木聚糖水解酶不同。该研究不仅拓展了GH30 双功能木聚糖水解酶的生物技术应用,还为研究单功能酶转变为多功能酶的分子决定因素提供了一个良好的范例。GH43 家族主要作用于木聚糖侧链,发挥β-木糖苷酶和(或)阿拉伯呋喃糖苷酶功能,部分成员具有多功能性。Liu 等报道了来源于短小芽孢杆菌(Bacillus pumilus TCCC 11350)的嗜冷酶rXYL,同时具有木糖苷酶和阿拉伯呋喃糖苷酶两种活性。序列比对显示rXYL 包含木糖苷酶保守的催化氨基酸Asp14、Asp127、Arg287。该酶在30 ℃活力最高,在4 ℃和0 ℃条件下分别维持其最大活力的26%和18%,且具有相对较强的木糖耐受性。此外,rXYL与内切木聚糖酶在低温下连续或同时降解山毛榉木聚糖显示出较高的协同效应(分别为11.14 倍和16.21 倍)[33]。然而,该酶识别木糖苷和阿拉伯呋喃糖苷底物的分子机制尚不清楚,有待进一步探索。Basit 等报道了嗜热毁丝霉菌(Thermothelomyces thermophila)来源的TtXy43,仅含有单独的GH43 家族催化结构域,但同时具有内切木聚糖酶、β-木糖苷酶、α-L-阿拉伯呋喃糖苷酶3 种活性,可以将木聚糖高效降解生成木糖单体[34]。具有的3 种活性共用相同的催化氨基酸Asp134 和Glu228,并且3 种活性在相似的催化条件下发挥功能。结构模拟和分子对接显示,TtXyn43 的活性位点形成狭长的底物结合口袋以容纳较长的木聚糖底物进入,Glu176、Asp85 和Asp38 对于底物识别起到关键作用。Rohman 等报道了喜热噬油芽孢杆菌(Geobacillus thermoleovorans IT-08)来源的Xyl,含有GH43 家族催化结构域和β-三明治辅助结构域,同时具有β-木聚糖酶和阿拉伯呋喃糖苷酶的活性,两种活性共用相同的催化氨基酸Asp14 和Glu177[35]。通过解析Xyl 单体(PDB:5Z5D)及其与产物复合体(PDB:5Z5I)结构发现,该酶的活性中心位于催化结构域漏斗状中心空腔,可以容纳木糖和阿拉伯糖两个糖基单元,分别位于底物结合口袋-1 和+1 亚位点。Asp121、His238、Arg269 对于识别阿拉伯糖基发挥重要的作用,保守的Glu198 对于识别木糖基起着重要的作用。此外,其他GH 家族的单催化结构域多功能木聚糖水解酶的作用机制也有一定的报道。例如,Tong 等报道了嗜热细菌(Dictyoglomus turgidum)来源的GH3 家族木糖苷酶/阿拉伯呋喃糖苷酶Dt-2286,序列比对得出其催化氨基酸为Asp276 和Glu507[36]。重组Dt-2286 在摇瓶大肠杆菌中β-木糖苷酶的表达量高达270 U/mL,该酶的最适反应条件为pH 5.0、98 ℃,且该酶具有较高的热稳定性和有机溶剂的耐受性,与内切木聚糖酶协同作用于木聚糖,可释放高达15.08 倍的还原糖。Bouraoui 等从突尼斯温泉细菌中分离出仅含有GH51 家族催化结构域多功能酶THSAbf,该酶对pNPAf 具有较高活力(kcat/Km=1 050 L/(mmol·s)),且可以水解桦木和山毛榉中的4-O-甲基葡糖醛酸木聚糖及甜菜来源的线性及分支阿拉伯聚糖[37]。阿拉伯呋喃糖苷酶和内切木聚糖酶的催化氨基酸相同(Glu177 和Glu296),且两种活性在60 ℃下和较宽的pH 范围(pH 4.0~7.0)内较为稳定。结构模拟和分子对接发现,THSAbf 具有较宽的活性位点裂缝用于发挥多种功能。此研究对简化及构建成本低、效率高的木聚糖水解酶复配体系具有重要的意义。

图3 TcXyn30B 的晶体结构Fig.3 Crystal structure of TcXyn30B
Hameleers 等从海狸粪便基因组中鉴定到双功能乙酰木聚糖酯酶/阿魏酸酯酶BD-FAE,仅包含单独的CE 结构域,被归类于一个新的具有广泛底物特异性的酯酶家族[38]。该酶不仅水解桦木来源的乙酰葡糖醛酸木聚糖(AcGX)释放乙酸,发挥乙酰木聚糖酯酶功能;还能从玉米纤维来源的乙酰基和阿魏酰基高度取代的低聚木糖(AcFaXOS)中释放阿魏酸,发挥阿魏酸酯酶功能。BD-FAE 的晶体结构(PDB:6TKX)显示其采取典型的α/β 水解酶折叠构象,活性中心包含保守的催化三联体Ser128、Asp237、His269。活性位点附近存在一个暴露于溶剂中的较浅凹槽,在空间上可以与高度取代的较大底物结合。此外,Dilokpimol 等研究了10 种真菌来源的新型CE1 酶,其中,CE1 亚家族2 的瘤孢棒囊孢壳(Corynascus sepedonium)Fxe1和嗜热毛壳菌(Chaetomium thermophilum)FxeA 以及CE1 亚家族5 的球毛壳菌(Chaetomium globosum)Fxe1 具有阿魏酸酯酶和乙酰木聚糖酯酶双重活性,能够从小麦阿拉伯木聚糖中同等程度地释放乙酸和阿魏酸[39]。结构模拟显示催化位点附近的氨基酸变化可能决定其不同的底物识别特异性:具有严格乙酰木聚糖酯酶活性CE1 亚家族1 中保守的Trp,在具有双功能的CE1 亚家族2 中被较小侧链的残基(Ala、Pro、Ser、Thr 等)取代,允许其在催化位点结合较大的底物阿魏酸;而与同家族的阿魏酸酯酶相比,CE1 亚家族5 的CgFxe1 存在独特的Thr226 可能与乙酰基形成相互作用,从而促进其双重活性。
此外,还有一些单活性位点单催化结构域水解酶不仅降解木聚糖,还能催化其他多糖底物水解。例如,Wang 等从木聚糖降解菌株芽孢杆菌(Bacillus sp.KW1)中鉴定出GH10 家族耐热酶XynA,该酶不仅可以水解木聚糖类底物,而且对微晶纤维素、羧甲基纤维素(CMC)、对硝基苯纤维二糖苷(pNPC)、对硝基苯基吡喃葡萄糖苷(pNPG)等纤维素类底物具有水解活性[40]。Phakeenuya 等报道了来源于解凝乳类芽孢杆菌(Paenibacillus curdlanolyticus B-6)的GH9 家族的 多功能 酶PcMulGH9,具有广泛水解纤维素、甘露聚糖、木聚糖中β-1,4 糖苷键的能力[41]。Liu 等报道了枯草芽孢杆菌(Bacillus subtilis BS-5)来源的GH13 家族α-淀粉酶/内切葡聚糖酶/内切木聚糖酶BSGH13[42],以及Han 等报道了嗜热毛壳菌(Chaetomium thermophilum)来源的GH7 家族具有纤维二糖水解酶和木聚糖酶活性的耐热酶Ctcel7 等[43]。此类多功能酶具有多种复杂底物的降解能力,对其适应条件和作用机理的研究为工业化应用奠定基础。
2.2.2 多活性位点型多功能木聚糖水解酶 多活性位点型多功能木聚糖水解酶在单一催化结构域中含有不同的催化中心,各自催化不同的反应,具有“兼职”(moonlighting)酶的特征。与单活性位点型多功能木聚糖水解酶相比,这一类酶的研究较少。Cao 等报道的热解纤维素菌(Caldicellulosiruptor lactoaceticus)来源的乙酰酯酶/木糖苷酶CLH10 是一类特殊的双功能木聚糖水解酶[44]。在一级序列上,CLH10 的酯酶与糖苷酶保守序列片段呈混合型分布,不能被归类于某一家族。结构研究显示其具有独立的木糖苷酶和乙酰酯酶活性位点,分居催化结构域的两侧,互不干扰。CLH10 结构整体(PDB:6A6O)呈现典型的酯酶构象,具有保守的催化三联体,在其背侧的柔性环偶然进化形成一个糖苷酶活性中心,从而实现对乙酰酯和木糖苷底物的识别与催化。该研究显示位于蛋白质表面的环状区可以作为蛋白质工程潜在位点,用于设计和产生具有新型多功能酶催化剂(见图4)。此外,Shahid 等从嗜酸纤维素分解菌(Acidothermus cellulolyticus 11B)中挖掘到GH10 家族的内切木聚糖酶/阿拉伯呋喃糖苷酶/乙酰木聚糖酯酶XynB,能够水解多种木聚糖天然底物产生木糖、木二糖、阿拉伯糖、乙酸[45]。该酶包含一个GH10 催化结构域和两个纤维素结合结构域CBM2、CBM3,与不溶性木聚糖相比,两个CBM 的存在使其对胶体微晶纤维素具有更高的结合力。该酶在70 ℃和pH 6.0 时具有最佳活性,且在高达70 ℃时保持稳定。这些特性使其具有极大的工业化应用潜力。然而,该酶通过单一GH10 结构域催化糖苷键和酯键水解的分子机制尚不清楚,亟待深入探索。

图4 CLH10 的晶体结构Fig.4 Crystal structure of CLH10
2.3 多催化结构域的多功能木聚糖水解酶
这一类多功能木聚糖水解酶典型特征为含有多个催化结构域模块,不同的催化结构域通过各自的催化位点执行不同的功能,不同催化结构域之间与其他结构域或柔性环构成的连接区相连。多催化结构域多功能木聚糖水解酶大多包含GH 和CE 催化结构域,可以同时水解糖苷键与酯键。Yang 等从产黄青霉(Penicillium chrysogenum P33)中挖掘到含CE1 和GH62 催化结构域的双功能乙酰木聚糖酯酶/阿拉伯呋喃糖苷酶PcAxe,两种酶在相似条件下发挥功能,最适pH 均为7.0,乙酰木聚糖酯酶和阿拉伯呋喃糖苷酶的最适温度分别为40 ℃和50 ℃,且在广泛pH 范围下稳定,并具有金属离子耐受性[46]。PcAxe 与重组木聚糖酶协同水解脱木质素玉米秸秆,协同度高达1.35,且用等量的PcAxe 代替商用纤维素酶,脱木质素玉米秸秆中葡萄糖的释放量提高了51%,显示出较高的纤维素水解效率,表明该酶具有良好的应用前景。类似地,Mroueh 等从解纤维梭菌(Ruminiclostridium cellulolyticum)中挖掘到阿拉伯呋喃糖苷酶/乙酰木聚糖酯酶(GH62-CE6-1240)[47]。该酶包含GH62 和CE6 双催化结构域,可催化不溶性小麦阿拉伯木聚糖的脱乙酰化和阿拉伯糖基的水解,且对阿拉伯聚糖(I 型鼠李糖半乳糖醛酸聚糖的一种成分)也有活性,表明其参与了果胶的降解。因此,GH62-CE6-1240 在植物细胞壁多糖的降解中起关键作用。Wang 等从高效降解玉米秸秆的细菌菌群EMSD5 中挖掘到双功能内切木聚糖酶/阿魏酸酯酶rXyn10A/Fae1A,可以同时作用于木聚糖的主链和侧链[48]。该酶包含GH10 和CE1 双催化结构域和CBM2 和CBM13 碳水化合物结合模块。两种酶的最适温度均为50 ℃,木聚糖酶和阿魏酸酯酶的最适pH 分别为6.0 和7.0,两种酶均在较宽的pH 范围内较为稳定。生化分析显示,仅含有CE1 催化结构域的截短体具有较低的阿魏酸酯酶活性,而同时含有CE1-GH10 催化结构域的截短体使阿魏酸的产量显著增加,但仅为全长蛋白质产阿魏酸总量的78%,表明处在同一多肽链上的不同催化结构域对阿魏酸的释放具有分子内协同效应。rXyn10A/Fae1A 能够从农业废弃物中产生高达2.78 mg/g 的阿魏酸和160.5 mg/g 的木糖/低聚木糖,显示出巨大的应用潜力。Kmezik 等从多毛拟杆菌(Bacteroides eggerthii)多糖利用位点(PUL)27 中鉴定到含双催化结构域CE15 和GH8 的葡糖醛酸酯酶/外切木聚糖酶BeCE15A-Rex8A,酶学分析发现,N 端的CE15 葡糖醛酸酯酶结构域对葡糖醛酸酯酶(GE)模式底物活性较差,可能与活性位点附近关键氨基酸的替换有关;而C 端GH8 外切木聚糖酶结构域可以从低聚木糖的还原端释放木糖单体,但对较长的木聚糖链没有活性。全长蛋白质和单独GH8 结构域分别与商用GH11 木聚糖酶协同催化玉米芯,结果发现木寡糖的含量均得到了明显的提升,但并未观察到两个结构域间的协同效应[49]。该研究报道的CE15-GH8 的结构域组成非常罕见,同时强调了针对PUL 的基因挖掘对于发现新酶类型与结构具有重要意义。Pereira 等从人体肠道结肠拟杆菌中挖掘到阿魏酸酯酶/乙酰酯酶Bi1040-FAE,能够从阿魏酰酯化的寡糖、去淀粉麦麸、不溶性小麦阿拉伯木聚糖中释放阿魏酸,也可以水解乙酰化燕麦木聚糖释放乙酸[50]。由于该酶的表达量较低,通过序列比对获得了多毛拟杆菌来源的BeGH43/FAE。该酶N 端为GH43 家族融合蛋白质序列,具有阿拉伯呋喃糖苷酶活性;C 端为Bi1040-FAE 同源蛋白质,具有乙酰酯酶和阿魏酸酯酶的活性。结构解析(PDB:6LMY)显示,BeGH43/FAE 的GH43 结构域具有典型的五叶片β-螺旋桨结构和β-三明治型结构,活性中心具有保守的催化氨基酸Asp149 和Asp38;酯酶结构域呈现保守的α/β 水解酶折叠构象,具有保守的催化三联体Ser634、His774、Asp742(见图5)。分子动力学模拟显示BeGH43/FAE 的酯酶结构域具有较大的底物结合口袋和更高的底物可及性,可以容纳较大的阿魏酸酯类底物。这类融合阿拉伯呋喃糖苷酶和酯酶活性的多结构域多功能酶可以提高复杂阿拉伯木聚糖的降解效率,有助于肠道菌降解膳食纤维释放阿魏酸。
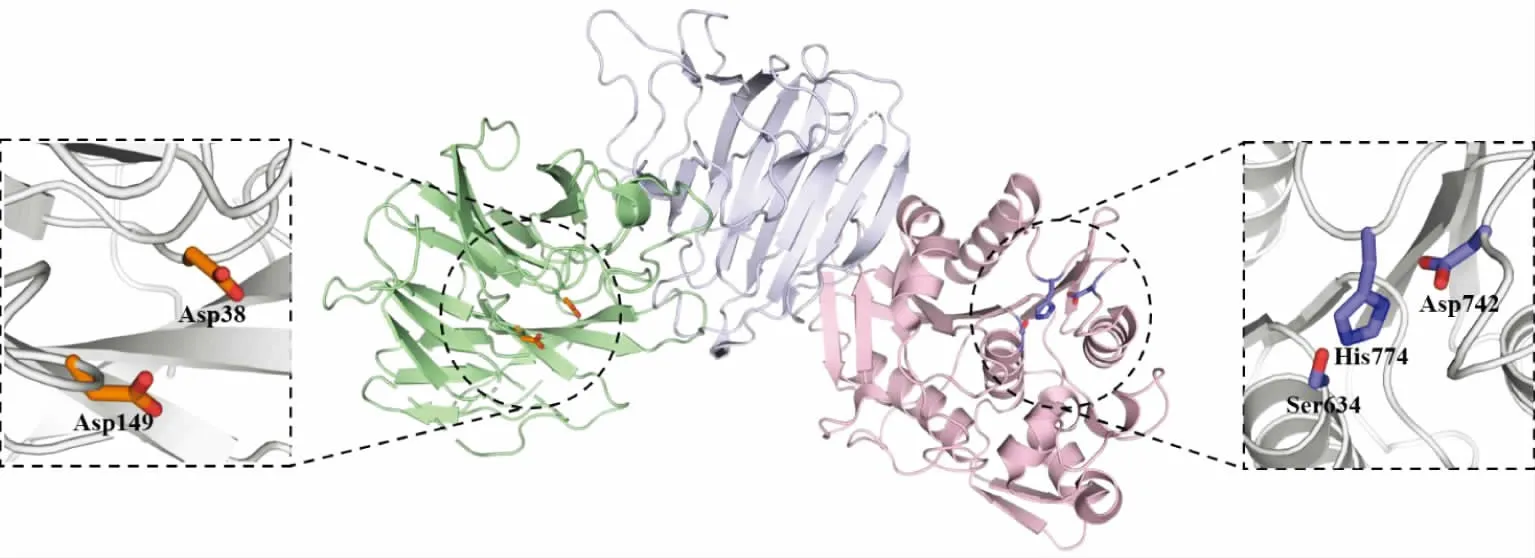
图5 BeGH43/FAE 的晶体结构Fig.5 Crystal structure of BeGH43/FAE
具有多个CE 催化结构域的木聚糖水解酶也有报道。Kmezik 等报道了营发酵单胞菌(Dysgonomonas mossii)来源的双功能乙酰酯酶/阿魏酸酯酶DmCE1B,含有两个CE1 催化结构域和中间的CBM48 结构域[51]。生化研究显示,包含N 端CE1和CBM48 的截短体(DmCE1B_nt)同时具有乙酰酯酶和阿魏酸酯酶的活性,与全长蛋白质活性相似;而C 端CE1 和CBM48 截短体(DmCE1B_ct)仅具有较低的阿魏酸酯酶活性。DmCE1B_ct(PDB:7B5V)的结构解析显示,其具有10 链组成的β-三明治型CBM48 结构域和酯酶典型的α/β 水解酶折叠结构,中间由一个较短的10 个氨基酸组成的连接区相连。酯酶保 守的催 化三联 体(Ser542、His638、Glu606)位于CE1 和CBM48 结构域之间的凹槽中,底物阿魏酸甲酯位于活性中心凹槽的边缘,与催化丝氨酸距离较远。然而,由于全长及具有双重活性的DmCE1B_nt 的结构尚未报道,该酶实现双功能的分子基础以及不同结构域对全长蛋白质的调控机制并不清楚,有待进一步探索。
此外,Kmezik 等也从卵形拟杆菌(Bacteroides ovatus)和约氏黄杆菌(Flavobacterium johnsoniae)中鉴定到两种具有相似结构域组成的酯酶,包含N 端的CE6 结构域和C 端的CE1 结构域[52]。其中CE6表现出乙酰酯酶活性,而CE1 表现出阿魏酸酯酶活性。这两种酶可以显著提高商用木聚糖酶水解玉米芯(接近2 倍)和日本山毛榉(超过20 倍)的能力。并且,与添加两个分离的单催化结构域相比,全长蛋白质协同商用木聚糖酶水解天然木聚糖底物的能力显著提升,表明两个酯酶结构域之间存在分子内协同作用,这种协同增强效应的分子机制尚待阐明。此类多功能酶是改善生物质有效降解的重要工具,在食品、饲料、造纸等工业生产中具有广阔的应用前景。
此外,还有一些研究着重分析催化结构域之间的连接区或其他结构域对酶催化功能的影响。例如,Krska 等从极端嗜热厌氧菌(Caldicellulosiruptor kristjanssonii)中挖掘到多模块嗜热双功能酶CkXyn10C-GE15A,该酶包含N 端的两个CBM22、内切木聚糖酶和葡糖醛酸酯酶催化结构域和中间的3 个CBM9,以及C 端的钙黏素结构域和两个SLH 结构域[53-54]。两种酶活性的最适反应条件存在差异,且两个催化结构域混合催化天然木聚糖底物时,并未发现明显的协同效应。5 个碳水化合物结合模块对不溶性多糖、可溶性多糖、低聚糖具有不同的底物偏好性,并对催化结构域的活性和热稳定性起促进作用。Holck 等从废水处理的污泥基因组中鉴定出一种含有3 个催化结构域的阿魏酸酯酶/阿拉伯呋喃糖苷酶/内切木聚糖酶CE1-GH62-GH10,该酶发挥3 种功能时的催化条件存在差异:以不溶性小麦阿拉伯木聚糖为底物测定的GH62、GH10 的最适温度为45 ℃,CE1 的最适温度为35 ℃;GH10的最适pH 范围较宽泛(pH 3.0~8.0),GH62 最适pH 7.0~9.0,CE1 最适pH 10.0~11.0[55]。值得注意的是,全长蛋白质与单催化结构域的等摩尔混合体系相比活性相当,但热稳定性有一定提高;GH62-GH10对底物的催化效率比单独的GH10 高2.5 倍,而CE1 需要在GH10 存在的情况下从底物中释放阿魏酸。几种多功能木聚糖水解酸特性见表2。

表2 多功能木聚糖水解酶的特性Table 2 Properties of multifunctional xylanolytic enzyme

续表2
3 多功能木聚糖水解酶在食品领域的潜在应用
随着人们对安全和健康食品需求的不断增加,研发功能性食品成为了关注的焦点。木聚糖是半纤维素的主要成分,存在于水果和蔬菜的初级细胞壁和膳食纤维中[56-57],对果汁澄清和面包加工产生影响,可以加入木聚糖水解酶进行改善,从而获得澄清度和营养成分更高的果汁和蓬松度、口感更好的面包。此外,酶解木聚糖可获得益生元低聚木糖,作为功能性食品添加剂,具有可降低血液胆固醇、增加钙吸收、抗氧化、维持胃肠道健康、降低结肠癌风险等功能。以下详细综述了木聚糖水解酶在食品领域的研究进展和发展前景。
3.1 果汁澄清
自从确定了果汁的营养价值以来,果汁已成为各年龄段人群中流行的健康饮品,制备优质、有益健康的果汁已成为商业食品行业的焦点。任何果汁的制备都涉及3 个步骤,包括提取、果汁澄清和稳定化。然而,商业果汁生产的主要障碍是半纤维素的存在导致果汁澄清度和产量低、果汁浓缩和巴氏杀菌困难以及商业市场可接受度低。因此,食品公司一直在大力研究果汁澄清的方法从而改善其外观以及延长储存时间[58]。早期果汁澄清仅依靠物理和化学方法或超滤技术去除半纤维素,但这些方法无法达到令人满意的澄清水平,进而有研究发现酶法是提高果汁澄清度的最有效方法。其中,在果汁中加入内切木聚糖酶可有效降解果汁中的多糖,从而提高果汁的澄清度和香气,延长包装产品的贮藏寿命。有研究者从日本曲霉(Aspergillus japonicus)中提纯了内切木聚糖酶(UFMS 48.136),并成功利用该酶对芒果汁、香蕉汁、橙汁、菠萝汁和蜜橘汁进行了澄清处理,其澄清度分别达到51.11%、9.99%、2.66%、6.11%和8.54%[59]。此外,α-L-阿拉伯呋喃糖苷酶也可用于果汁的澄清,彭程等将黑曲霉(Aspergillus niger)来源的α-L-阿拉伯呋喃糖苷酶加入柚子汁中,其澄清度提高了15.18%[60]。有研究者研究了α-L-阿拉伯呋喃糖苷酶和商用内切木聚糖酶的协同加入对苹果汁、葡萄汁、橙汁和桃汁的作用,结果发现,与两酶单独处理相比,两酶协同加入可提高果汁的产量和澄清度[61]。因此,木聚糖水解酶在果汁澄清中发挥着重要的作用,且两个单功能木聚糖水解酶在果汁澄清中具有协同作用,但加入两种酶成本较高,且不同酶的作用条件可能存在差异,操作困难。因此,在果汁加工中,多功能木聚糖水解酶的应用将是未来的发展趋势,具有广阔的应用前景。
3.2 烘焙
目前,食品公司投入了大量精力以开发高质量感官的健康产品。例如,在烘焙产品方面,使用酶代替化学品是改善面包黏度、质地和风味的有效途径。在烘焙加工中,全麦面粉中麸皮的存在会导致面筋蛋白稀释,从而降低面团和面包的一些关键质量属性,例如面包体积、颜色、质地和味道等。阿拉伯木聚糖是位于小麦胚乳中的非淀粉多糖,占面粉的0.4%~0.8%(质量分数)。虽然质量分数很低,但其具有高吸水性,是影响面团发酵和面包口感等的重要因素。内切木聚糖酶可随机催化阿拉伯木聚糖主链的木糖残基间的共价键,降低平均聚合度,从而改变面团的流变特性、面包的比容和面包屑的硬度等[62-63]。然而,阿拉伯木聚糖含有35%~40%的阿拉伯糖取代基,部分阿拉伯糖取代基含有酯键相连的阿魏酸,这些侧链取代基的存在会阻碍内切木聚糖酶对阿拉伯木聚糖主链的水解。因此,需要加入阿拉伯呋喃糖苷酶和阿魏酸酯酶从而提高阿拉伯木聚糖的水解效率,且释放出低热量的阿拉伯糖和对人体有益的阿魏酸。已报道嗜热子囊菌(Thermoascus aurantiacus)产生包含木聚糖酶、木糖苷酶和阿拉伯呋喃糖苷酶的酶混合物,这些酶混合物在长时间温育后的固态发酵过程中释放木糖和阿拉伯糖,从而使面包体积增加了22%,而松脆度和支链淀粉回生分别减少了25%和17%[64]。此外,Xue 等发现与单独加入木聚糖酶/阿拉伯呋喃糖苷酶相比,组合加入木聚糖酶和阿拉伯呋喃糖苷酶具有更强的阿拉伯木聚糖水解能力,且对面团流变性质和品质有协同作用[65]。
3.3 低聚木糖的制备
低聚木糖是由多个木糖单元通过β-1,4-木糖苷键连接而成的寡聚糖,在结构上,低聚木糖根据构成其化学结构的单体数量以不同的形式存在,包括木二糖、木三糖、木四糖、木戊糖、木己糖等。根据物理化学特性,低聚木糖被归类为不易消化的低聚糖,在宽泛的温度范围(最高达100 ℃)和pH 条件(pH 2.5~8.0)下具有较好的稳定性,是糖尿病患者的功能性食品和膳食甜味剂的理想添加剂[66-68]。此外,低聚木糖可通过免疫刺激增加健康双歧杆菌(Bifidobacterium)的数量从而改善肠道功能[69],可能降低患结肠癌的风险,具有益生元作用[70-71]。作为食品成分,低聚木糖具有可接受的气味和低热量,并且不会导致龋齿的生成,可用于抗肥胖饮食[72]。目前,低聚木糖的生产主要采用化学法及酶法等。其中,化学法是在高温高压下使用酸水解或碱提取获得低聚木糖,该方法会产生大量的副产物,如糠醛和羟甲基糠醛等,其会降低低聚木糖的益生元特性[73]。然而酶法生产低聚木糖不会产生无用的副产物,且对环境无污染,是食品工业生产低聚木糖的首选方法[74]。Wang 等以200 mg 去淀粉麦麸为底物,通过加入双功能阿魏酸酯酶/木聚糖酶进行酶解,其低聚木糖(木糖/低聚木糖)产量可达18.2 mg[48]。Li 等通过加入内切木聚糖酶和β-木糖苷酶水解杨木锯末生产低聚木糖,结果发现当仅加入内切木聚糖酶或β-木糖苷酶时,低聚木糖的产率较低,而当两种酶协同加入水解时,低聚木糖的产率可高达93.9%[75]。这些研究表明,相比于单独使用木聚糖酶,采用多功能酶或多酶协同作用于木聚糖分子可以明显提高木聚糖主链的水解效率。随着人们对功能性食品需求的不断增加,酶法生产益生元低聚木糖具有很好的市场前景,后续采用低成本、高活性的多功能木聚糖水解酶生产低聚木糖是一种可持续发展的有效策略。
4 展 望
近年来,随着酶工程技术领域的发展及与食品生物领域的交叉融合,针对食品体系及特殊食品分子的酶催化体系构建成为酶工程研究领域的热点之一。与传统的单酶或多酶催化体系相比,兼具多种催化活性的多功能酶在酶解木聚糖等复杂底物时具有降低生产成本、提高催化效率、简化酶解工艺等明显优势。目前,针对天然多功能木聚糖水解酶的挖掘表达、酶学表征和催化机制解析已取得卓有成效的研究进展。然而,由于不同类型的多功能木聚糖水解酶的结构、与底物结合及催化的酶动力学机制较为复杂,对该类酶的分子调控机制仍知之甚少,尚未实现工业化应用。未来的研究可以从以下方面展开:1)通过木质纤维素降解优势菌群的多组学研究和生物信息数据挖掘技术,筛选和挖掘具有更好酶学特性的新型多功能木聚糖水解酶;2)通过序列比对、结构解析等研究手段,更加全面深入地探索不同类型多功能木聚糖水解酶催化分子机制,同时使用定向进化和祖先序列重建探索酶系统,进一步了解酶进化分子机制,特别是探究单酶同时发挥多种功能的分子基础;3)通过理性设计、定向进化等方法改变酶的分子结构以提高催化活性及热稳定性,如在活性中心引入新的残基可以改变酶与底物的静电相互作用,通过活性位点周围氨基酸的改造将天然单功能酶进化为多功能酶等;4)基于酶学机制的高效融合多功能木聚糖水解酶的人工构建与修饰,目前人工构建融合酶的过程中经常存在错误折叠、功能或结构干扰、蛋白质间连接器断裂等问题,后续可以尝试将更稳定的酶放在融合酶的前端从而克服上述问题,此外,使用翻译后修饰位点如N-乙酰化或糖基化位点可以提高酶的稳定性;5)结合人工智能技术精确指导特异性天然多功能酶筛选与精准设计,通过多学科的综合交叉研究,解决高效食品酶创制中的共性关键问题,进而定制更符合工业需要的多功能酶催化体系,推动相关产业的快速发展。
猜你喜欢
阅读(中年级)(2022年9期)2022-10-08
生物信息学(2022年1期)2022-04-01
红蜻蜓·低年级(2022年2期)2022-03-19
红蜻蜓·低年级(2021年12期)2021-12-19
天然产物研究与开发(2018年8期)2018-09-10
科技创新导报(2018年1期)2018-05-07
天然产物研究与开发(2018年4期)2018-05-07
石油化工应用(2018年3期)2018-03-24
分析化学(2017年12期)2017-12-25
广东饲料(2016年8期)2016-02-27
