
菜心BrHsfA3 基因克隆及其对高温胁迫的响应
2023-03-07 12:55庞强强孙晓东周曼蔡兴来张文王亚强
生物技术通报 2023年2期
庞强强 孙晓东 周曼 蔡兴来 张文 王亚强
(1. 海南省农业科学院蔬菜研究所,海口 571100;2. 海南省蔬菜生物学重点实验室,海口 571100;3. 海南省蔬菜学院士团队创新中心,海口 571100)
菜心(Brassica campestrisL. ssp.chinensisvar.utilisTsen et Lee),又名菜薹,是十字花科芸薹属白菜亚种的变种[1]。作为我国华南地区的特产蔬菜之一,其品质柔嫩,营养丰富,深受消费者的喜爱。菜心生长周期短,复种指数高,可周年生产供应,在应对蔬菜生产淡季上具有重要作用。菜心喜温但不耐高温,生长温度超过30℃就会产生菜薹纤细、产量降低和品质下降等问题[2-3]。因此,培育耐高温菜心品种是解决上述问题的根本措施。
热激转录因子(heat stress transcription factor,Hsf)是调节生物细胞内热激反应的一类重要因子,在高温胁迫下通过与热激元件(Hse)结合而激活下游基因的转录表达[4]。Hsf基因普遍存在于各种植物中,目前已发现甜瓜有22 个[5]、毛竹有41 个[6]、南瓜有36 个[7]、芥菜有60 个[8]、菜豆有30 个[9]Hsf基因。依据Hsf结构的不同,可将其分为A、B和C 三类家族,不同家族又分为多个亚族[10]。首个Hsf基因是从酵母中克隆得到[11],后来研究者从番茄中获得了第一个植物的Hsf基因[12]。HsfA3是热激转录因子A 类家族的成员,在正常条件下,HsfA3 属于组成型表达,分布在细胞质中;当受到非生物胁迫时,为诱导型表达,且作用在细胞核中[13-14]。HsfA3基因能够直接或间接调控植物的生长发育和逆境反应过程。唐锐敏等[15]研究发现,马铃薯StHsfA3在耐热过程中起到正调控作用,可能协同StHsp26-CP和StHsp70来增强过表达转基因株系的耐热性。拟南芥高温胁迫转录组的数据显示,HsfA3是唯一在各种高温胁迫条件下和各个高温胁迫时期都有稳定表达的两个热激相关基因之一[16],在拟南芥中过表达或异源过表达番茄的HsfA3都可提高其耐热性[17-18]。除了调控植物高温胁迫响应过程外,HsfA3还可参与植物的干旱[19]、低温[20]、盐[21]、氧化[22]等其他非生物胁迫应答。
近年来,本团队针对菜心耐热性开展了一系列研究工作[23-25],建立了菜心高温胁迫转录组数据库,并从中获得了1 个差异表达基因。在此基础上,本研究采用全基因合成法(PCR-based accurate synthesis, PAS)克隆了该差异表达基因的序列,并对其蛋白理化性质、蛋白结构、系统进化等进行生物信息学分析,同时利用RT-qPCR 技术分析该基因在高温胁迫下的表达模式,以期为菜心耐高温机理研究和耐热育种奠定基础。
1 材料与方法
1.1 材料
试验于2021年12月-2022年3月在海南省农业科学院蔬菜研究所蔬菜生物学重点实验室进行。供试材料为苗期高温胁迫鉴定及田间自然高温鉴定均表现为耐热的自交系CX1-7 和热敏的自交系CX7-3[23]。挑选饱满、大小一致的菜心种子播种于营养钵中,待植株长至3-4 片真叶时进行间苗,每个营养钵中留一株幼苗,每个自交系种植60 株。采用常规栽培管理方式培养至抽薹开花时,移入25℃/18℃(昼/夜)、12 h/12 h(昼/夜)、光照80%、湿度70%的人工气候箱中预培养3 d,之后将其中30 株移入另一人工气候箱内进行高温胁迫处理,处理温度为37℃/27℃(昼/夜),除温度环境外,其他环境条件均与预培养时保持一致。分别取高温处理0 h(CK)和6 h(T6)的菜心根、叶、薹和花,用液氮冷冻后置于-80℃冰箱保存备用,试验均设3 次重复。
1.2 方法
1.2.1 总RNA 提取及cDNA 合成 参考Trizol 试剂说明书(北京全式金生物技术有限公司)分别提取菜心根、叶、薹和花的总RNA。利用cDNA 合成试剂盒(北京全式金生物技术有限公司)去除基因组DNA 并反转录合成cDNA。
1.2.2 基因克隆及生物信息学分析 从菜心高温胁迫转录组数据库中获取基因片段后委托武汉天一辉远生物科技有限公司进行全基因合成,得到基因全长序列。采用ORF finder(https://www.ncbi.nlm.nih.gov/orffinder/)预测氨基酸序列。采用Expasy 服务器中的Protparam 工具(https://web.expasy.org/protparam/)和ProtScale 工具(https://web.expasy.org/ protscale/)预测蛋白的基本理化性质。采用NetPhos 在线网站(https://services.healthtech.dtu.dk/service.php?NetPhos-3.1)分析蛋白的磷酸化位点;采用SignalP-5.0 在线网站(https://services.healthtech.dtu. dk/service.php?SignalP-5.0)预测蛋白的信号肽。采用NCBI-CDS 在线网站(https://www.ncbi.nlm. nih.gov/Structure/cdd/wrpsb.cgi)分析基因编码蛋白质的结构域。使用Heatster 在线工具(https://applbio.biologie.uni-frankfurt.de/hsf/ heatster/hsf_visualization_tool.php)对保守结构域进行注释。采用SOPMA 在线软件(https://npsa-prabi.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma. html)预测蛋白质的二级结构。采用Swiss-Model 在线网站(https://swissmodel.expasy.org/)预测蛋白质的三级结构。将克隆获得的片段在NCBI 在线网站(https://www.ncbi.nlm.nih.gov/)用Blastp 进行同源序列搜索,选取和下载同源性高、可靠性强的蛋白序列,采用MEGA6.0 软件中的邻接法(neighbor- joining method)构建系统进化树,重复计算次数设为1 000。
1.2.3 基因表达特性分析 分别将获得的菜心根、叶、薹、花的cDNA 稀释至100 ng/μL 作为RT-qPCR模板。以BrActin基因为内参基因[25],根据菜心BrHsfA3基因序列设计特异性引物,引物由上海生工生物工程有限公司合成,序列信息见表1。参考北京全式金生物技术有限公司的TransStart® TopGreen qPCR SuperMix 试剂盒说明书进行荧光定量分析,目的基因的表达量用2-△△Ct法来计算,每个基因的表达反应重复3 次。

表1 基因克隆和表达分析引物Table 1 Primers used for gene cloning and expression analysis
1.2.4 数据统计及分析 采用SPSS22.0 软件对数据进行统计分析,单因素分析采用Duncan 法,采用Excel2007 进行作图。
2 结果
2.1 菜心BrHsfA3基因克隆
根据前期转录组获得的菜心c27244.graph_c0 基因片段,通过全基因合成方法,获得了ORF 序列长度为906 bp 的基因片段,编码301 个氨基酸,含起始密码子ATG 和终止密码子TAA(图1)。在拟南芥基因组网站进行BLAST 比对,发现其与拟南芥中AtHsfA3基因最为相似,结合转录组注释结果,将其命名为BrHsfA3。

图1 BrHsfA3 的ORF 序列及氨基酸序列Fig. 1 ORF sequences and amino acid sequences of BrHsfA3
2.2 BrHsfA3蛋白理化性质分析
采用Proparam 在线网站分析了BrHsfA3基因编码蛋白质的理化性质,该蛋白分子式为C1488H2307N415O475S16,原子总数为4 701,相对分子量为34.12 kD,理论等电点为5.00,脂肪系数为55.42,平均疏水性为-0.827,不稳定系数为56.06。ProtScale 分析结果表明,BrHsfA3 蛋白质存在明显的疏水区和亲水区,其中在第5 位最高,为1.989;第44 位最低,为-3.022;表明BrHsfA3 蛋白为亲水性蛋白(图2)。NetPhos 磷酸化修饰预测分析表明,BrHsfA3 蛋白存在丝氨酸、苏氨酸和酪氨酸3 个磷酸化位点(图3),其中丝氨酸磷酸化位点21 个,苏氨酸磷酸化位点20 个,酪氨酸磷酸化位点3 个。利用SignalP-5.0 预测BrHsfA3 信号肽发现,该蛋白无信号肽(图4),为非分泌蛋白。

图2 BrHsfA3 蛋白疏水性/亲水性预测Fig. 2 Prediction of the hydrophobic/hydrophilic of BrHsfA3 protein
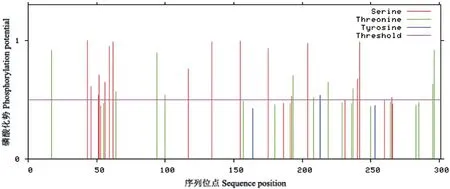
图3 BrHsfA3 蛋白氨基酸翻译后磷酸化修饰预测Fig. 3 Prediction of phosphorylation site after amino acid translation of BrHsfA3 protein

图4 BrHsfA3 蛋白信号肽预测Fig. 4 Prediction of the signal peptide prediction of BrHsfA3 protein
2.3 BrHsfA3蛋白结构预测分析
采用NCBI-CDS 在线网站分析BrHsfA3基因编码蛋白质的结构域发现,该蛋白有1 个特殊位点,属于Hsf_DNA-bind 超级家族,具有1 个Hsf 保守结构域(图5-A)。在靠近N 端第11-51 位含有氨基酸残基形成的DNA 结合结构域(DBD),第69-137位氨基酸残基形成的寡聚化结构域(OD),第146-156 位氨基酸残基形成的核定位信号(NLS),C 端第271-277 位氨基酸残基形成的转录激活结构域(AHA)(图5-B)。对BrHsfA3 蛋白的二级结构分析发现,BrHsfA3 蛋白二级结构中含有丰富的α-螺旋和无规则卷曲,其中参与形成α-螺旋的氨基酸有138 个,占总氨基酸的45.85%;142 个氨基酸参与形成无规则卷曲,占总氨基酸的47.18%;参与形成延伸链的氨基酸有12 个,占总氨基酸的3.99%;9个氨基酸参与形成β-转角,占总氨基酸的2.99%(图6)。利用Swiss-Model 工具得到了BrHsfA3 蛋白的三级结构,与二级结构预测结果相符,含有大量α-螺旋和无规则卷曲(图7)。
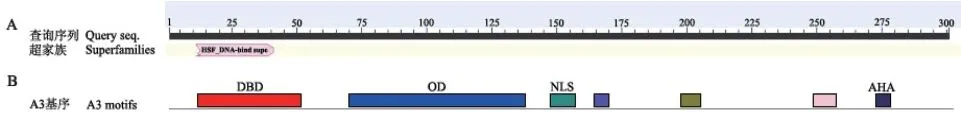
图5 BrHsfA3 蛋白的保守结构域及功能结构域Fig. 5 Conservative domains and functional domain of BrHsfA3 protein
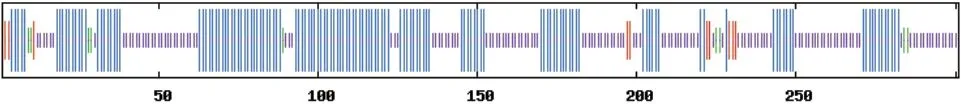
图6 BrHsfA3 蛋白二级结构预测Fig. 6 Prediction of BrHsfA3 protein secondary structure
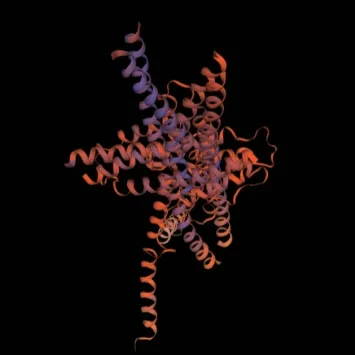
图7 BrHsfA3 蛋白三级结构预测Fig. 7 Prediction of the three-dimensional structure of BrHsfA3 protein
2.4 BrHsfA3基因系统进化分析
Blastp 同源性检索发现BrHsfA3 与22 种植物的HsfA3 蛋白相似性较高,这些植物为甘蓝型油菜(Brassica napus, KAH0907994.1)、 芜 菁(B. rapa,XP_009122679.1)、 花 椰 菜(B. oleraceavar.Oleracea, XP_013611350.1)、 萝 卜(Raphanus sativus,XP_018445490.1)、琴叶鼠耳芥(Arabidopsis lyrata subsp.Lyrata, XP_002873110.1)、 拟 南 芥(A.thaliana, NP_001318473.1)、 荠 菜(Capsella rubella,XP_006287844.1)、 亚 麻 荠(Camelina sativa,XP_019089984.1)、 棉 花(Gossypium hirsutum, XP_016691460.2)、白栎(Quercus lobata,XP_03092673 0.1)、三裂叶薯(Ipomoea triloba, XP_031131419.1)、木槿(Hibiscus syriacus,XP_039026314.1)、木薯(Manihot esculenta, XP_021619963.1)、花生(Arachis hypogaea, XP_025634986.1)、 苹 果(Malus domestica,XP_017191546.1)、狭叶羽扇豆(Lupinus angustifolius, XP_019426589.1)、 梨(Pyrusx bretschneideri,XP_009365327.1)、 葡 萄(Vitis vinifera, RVW482 07.1)、烟草(Nicotiana attenuata, XP_019250820.1)、牵牛花(Ipomoea nil, XP_019179577.1)、豇豆(Vigna unguiculata, XP_027936489.1)、 胡 杨(Populus euphratica, XP_011016541.1)。系统进化分析结果(图8)显示,BrHsfA3和甘蓝型油菜、芜菁、花椰菜、萝卜等十字花科作物处于同一分支,说明其亲缘关系较近。此外,苹果、梨、白栎、葡萄等聚为一支,烟草、三裂叶薯和牵牛花聚为一支,棉花、木槿、木薯和胡杨聚为一支,花生、狭叶羽扇豆和豇豆聚为一支,亚麻荠、荠菜、拟南芥和琴叶鼠耳芥聚为一支,表明HsfA3基因具有较高的保守性,所构建的系统发育树与其进化情况基本一致。

图8 菜心与其他科属植物HsfA3 氨基酸序列的分子系统进化树系统树Fig. 8 Phylogenetic tree of HsfA3 amino acid sequence from flowering Chinese cabbage and other plants
2.5 BrHsfA3基因在高温胁迫下的表达模式分析
从菜心高温胁迫转录组数据库中提取了BrHsfA3基因在高温胁迫前后的FPKM 值,由图9 可知,在正常温度条件下,BrHsfA3在热敏和耐热自交系叶片中的表达量基本一致,高温处理6 h 时,BrHsfA3基因在耐热自交系CX1-7 叶片中的FPKM 值较对照明显增加,热敏自交系CX7-3 叶片中无显著变化。为进一步验证转录组数据的正确性,采用RT-qPCR 技术分析了BrHsfA3基因在高温胁迫下的表达特性(图10)。在正常温度条件下,BrHsfA3基因在菜心的根、叶、薹和花中均有表达,其中BrHsfA3基因在热敏自交系CX7-3 的根中相对表达量最高,其次是薹、叶,花中的相对表达量最低;BrHsfA3基因在耐热自交系CX1-7 的不同组织中的相对表达量由高到低依次为薹>叶>根>花。高温处理后,BrHsfA3基因在热敏自交系CX7-3 的根中表达量显著上调,在叶、薹和花中的表达量无显著差异;在耐热自交系CX1-7 叶和花中的BrHsfA3基因表达量受高温诱导显著上调,根和薹中的表达量无显著差异。单独来看,BrHsfA3基因在热敏自交系 CX7-3 和耐热自交系CX1-7 高温处理前后叶片中的RT-qPCR 结果与转录组中的表达情况类似,说明转录组数据是真实可靠的。
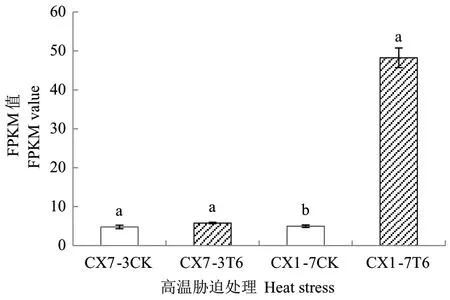
图9 BrHsfA3 基因在高温胁迫处理下的FPKM 值Fig. 9 FPKM value of BrHsfA3 gene under heat stress
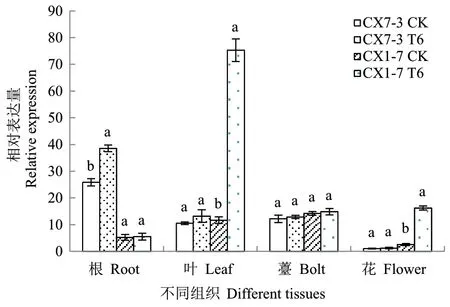
图10 BrHsfA3 基因在高温胁迫前后不同组织的相对表达量Fig. 10 Relative expressions of BrHsfA3 gene in different tissues under heat stress
3 讨论
菜心作为喜温蔬菜,在生长的各个阶段都容易遭受高温伤害,严重影响其生长发育,是造成产量和品质下降的主要非生物胁迫之一[26]。因此,阐明菜心高温胁迫响应机制,挖掘耐热基因,对于加速菜心耐热育种具有重要意义。越来越多的证据表明,热激转录因子在植物生长和非生物胁迫防御过程中具有重要作用[27-29]。
为了研究菜心耐高温胁迫的分子机制,本研究克隆得到了BrHsfA3基因。序列分析显示,BrHsfA3基因的ORF 长度为906 bp,编码301 个氨基酸。理化性质分析显示,BrHsfA3 蛋白相对分子量为34.12 kD,理论等电点为5.00,具有明显的疏水区和亲水区,为无信号肽的亲水性蛋白,存在丝氨酸、苏氨酸和酪氨酸3 个磷酸化位点。氨基酸序列分析表明,该蛋白序列具有典型的Hsf 家族保守结构域,在N端含有高度保守的DNA 结合结构域和寡聚化结构域,在氨基酸序列的中部含有核定位信号结构域,C 端含有转录激活结构域,缺少核输出信号结构域(NES),这种类似结构在辣椒CaHsfA3 中也有发现[30]。Hsf 蛋白结构的不同决定了各族转录因子行使的功能存在差异,A 类Hsfs主要负责热激基因的表达调控,B 类和C 类Hsfs 缺少AHA 基元不具备激活功能[31],因此推测BrHsfA3也可能具有转录活性和激活功能,这需要在后续研究中予以重点关注。BrHsfA3 蛋白二级和三级结构预测显示,无规则卷曲和α-螺旋占比较高,这与前人在马铃薯[15]上的研究结果一致。系统发育分析表明,BrHsfA3 氨基酸序列与甘蓝型油菜的亲缘关系最近,表明BrHsfA3可能与甘蓝型油菜BnaHsfA3具有相似的功能和作用机制。
在正常生长环境中,Hsfs几乎处于低活性或无活性状态,但当遭受不利于生长发育的情况时,Hsfs活性被迅速激发,特异性识别并与热激元件结合,从而激活体内的防御系统,以提高自身的抗性,减轻胁迫造成的伤害。由于每个植物热激转录因子的种类和结构不同,因此在胁迫响应机制和生长发育调控中的功能具有多样性[32-33]。HsfA3 是植物形成耐热性的重要因子之一[13,34]。已有研究表明,HsfA3 位于信号传递链的末端,拟南芥AtHsfA3是Dreb2A和Dreb2C的下游基因,其表达受它们的直接调控,从而提高植株耐热性[35]。为了预测BrHsfA3基因的生物功能,本研究结合菜心高温胁迫转录组数据库和RT-qPCR 技术,分析了BrHsfA3基因在高温下的表达模式,结果显示BrHsfA3基因的在菜心根、叶、薹和花中均有表达。在正常温度条件下,热敏自交系CX7-3 根中的表达量明显高于叶、薹和花,耐热自交系CX1-7 的薹和叶中表达量显著高于根、花。多数研究证明,HsfA3基因的表达对番茄[18]、多年生黑麦草[36]和甘蓝型油菜[37]的耐热性起正向调控作用,本研究中高温胁迫下耐热自交系叶、花和热敏自交系根中表达量均显著上调也直接证明了这一观点。另外,对比BrHsfA3在不同自交系间的表达量发现,除根中外,耐热自交系CX1-7 的叶、薹和花中在高温前后的表达量始终高于热敏自交系CX7-3,这与茄子SmHsfA3基因在叶片中的研究结果类似[38]。根中表达量则在热敏自交系中较高,推测产生的原因可能是由于Hsf基因介导的耐热调控机制非常复杂,BrHsfA3基因在不同耐热性植株的不同组织中对高温胁迫响应机制存在差异,耐热性的形成是由不同的Hsf基因共同作用的结果。
4 结论
本研究成功克隆得到菜心BrHsfA3基因,该基因ORF 全长为906 bp,编码301 个氨基酸。该蛋白相对分子量为34.12 kD,理论等电点为5.00,为无信号肽的亲水性蛋白,存在丝氨酸、苏氨酸和酪氨酸3 个磷酸化位点,具有1 个Hsf 保守结构域和典型的DBD、OD、NLS 和AHA 结构域,含有丰富的α-螺旋和无规则卷曲。BrHsfA3与甘蓝型油菜BnaHsfA3同源性最高。BrHsfA3基因在菜心不同耐热性植株及其不同组织中均有表达,在正常温度条件下,不同组织中耐热自交系CX1-7 的表达量由大到小依次为薹>叶>根>花,热敏自交系CX7-3 的表达量由大到小依次为根>叶>薹>花。高温胁迫处理后,耐热自交系CX1-7 的叶、花中和热敏自交系CX7-3 的根中表达量均显著上调,其余组织中该基因的表达不受高温影响。
猜你喜欢
源流(2022年11期)2023-01-29
广州大学学报(自然科学版)(2019年1期)2019-05-07
北京农学院学报(2019年1期)2019-02-22
河南农业科学(2017年4期)2017-04-12
西南农业学报(2016年5期)2016-05-17
饮食与健康·下旬刊(2016年9期)2016-05-14
西湖(2016年9期)2016-05-14
西南农业学报(2016年6期)2016-04-16
天津科技大学学报(2016年1期)2016-02-28
湖北师范大学学报(自然科学版)(2015年2期)2016-01-10
