
葡萄PLATZ家族基因鉴定及其在种子发育过程中的表达分析
2023-02-28 19:59向锐孙艺萌梁晨师校欣杜国强王莉
果树学报 2023年1期
向锐 孙艺萌 梁晨 师校欣 杜国强 王莉


关键词:葡萄;PLATZ;生物信息学;种子发育;表达分析
中图分类号:S663.1 文献标志码:A 文章编号:1009-9980(2023)01-0013-12
PLATZ(plant AT-rich sequence and zinc-binding protein)是一类新型的植物特异锌指类转录因子,具有PLATZ保守结构域(PF04640)。豌豆(Pisum sati-vum) PLATZ1是第一个被报道的PLATZ蛋白,是植物特异性、锌依赖DNA结合蛋白,可以非特异性地与富含A/T的碱基序列结合,发挥转录抑制作用,并具有C-X2-H-X11-C-X2-C-X(4-5)-C-X2-C-X(3-7)-H-X2-H(CHC4H2)和C-X2)-C-X(10-11))-C-X(10-11)-C這两个高度保守的蛋白锌指基序。
PLATZ家族鉴定分析已在拟南芥、水稻、玉米、芜青、小麦和油橄榄等物种中开展,并发现一系列在植物胁迫响应及生长发育调控方面具有重要功能的PLATZ转录因子。首先,在植物非生物胁迫响应方面,拟南芥AtPLA TZ1和AtPLA T22参与调控营养组织和种子的脱水抗性;在拟南芥中过表达棉花GhPLATZ/基因能显著增强萌发期和苗期的渗透胁迫耐受性。其次,在植物生长发育调控方面,异源表达水仙NtPLA TZ1基因能抑制烟草生长,沉默Nt-PLATZ1基因能促进烟草生长。玉米Fl3 (Zm-PLAT212)基因在玉米胚乳淀粉细胞中特异性表达,影响种子胚乳发育和养分储藏;水稻PLATZ转录因子GL6通过调控幼穗和谷粒的细胞增殖,决定籽粒长度和小穗;数;由此可见PLATZ转录因子在包括种子发育在内的多种生物学过程中有重要作用。
葡萄(Vitis vinifera L.)是世界上重要的果树作物之一,果实及其加工品具有极高的营养价值和经济价值。目前关于葡萄PLATZ转录因子家族的系统研究未见有报道,特别是参与种子发育的报道。笔者在本研究中利用生物信息学方法对葡萄PLATZ家族成员进行了鉴定,分析了基因结构、同线性关系、染色体定位、进化关系、蛋白质保守基序等,并对该家族成员在种子发育过程中与IAA处理下的表达模式进行了分析,为揭示葡萄PLATZ转录因子进化机制提供有用信息,为后续研究葡萄PLATZ家族基因的生物学功能奠定基础。
1材料和方法
1.1试验材料和处理
本试验于2021年4月-2021年10月在河北农业大学园艺学院生物技术实验室进行。试验材料为3个有核葡萄品种巨峰(Kyoho)、醉金香(Zui-jinxiang)、红地球(Red Globe)和2个无核葡萄品种火焰无核(Flame Seedless)、克瑞森无核(Crimson Seedless),均种植保存于河北保定河北农业大学(E115°29′,N38°51′)葡萄资源圃,年平均气温13.4℃,年平均日照2511 h,年平均降水量498.9 mm。分别对无核品种火焰无核、克瑞森无核及有核品种巨峰、醉金香于盛花后20、30、40、50 d收集胚珠。配制100μmol·L-1IAA溶液均匀喷施于红地球葡萄植株嫩叶,以无菌水喷施嫩叶为对照,于处理1、3、6h后收集嫩叶。每个处理设置3次生物学重复,收集的胚珠和叶片样品立即放入液氮中,并存于-80℃冰箱中备用。
1.2葡萄PLATZ家族成员鉴定
通过Pfam数据库(http://pfam.xfam.org/search#tabview=tabl)获取PLATZ家族蛋白保守结构域(PF04640)的隐马尔可夫(HMM)文件,利用SPDEhmmersearch功能(HMMER 3.0)获得候选蛋白,以所有候选蛋白序列作为查询索引,使用国家生物技术信息中心(http://www.ncbi.nlm.nih.gov/)BLASTp程序默认设置下搜索葡萄基因组同源蛋白,将候选NCBI同源蛋白利用葡萄基因组数据库(http://www.genoscope.cn.fr)和葡萄基因组CRIBI生物技术网站(http://genome s.cribi.unipd.it/)进一步进行同源比对筛选,并利用SMART (http://smart.embl-heidelberg.de)和Pfam数据库(http://pfam.xfam.org/search#tabview=tabl)人为核对去除结构域不完整的蛋白,获得最终鉴定的PLATZ成员,并依据基因染色体位置命名。利用Ex PASy数据库(https://web.expasy.org/protparam/)中的Prot Param功能对PLATZ蛋白质的等电点、分子质量、不稳定系数、脂肪系数等基本信息进行分析。使用PSORT在线网站(https://wolfpsort.hgc.jp/)进行葡萄PLATZ家族成员亚细胞定位预测。
1.3 PLATZ家族同线性分析
从植物基因组复制数据库(http://chibba.agtec.uga.edu/duplication/)下载葡萄PLATZ自身基因组间及葡萄与拟南芥基因组间的同线区域,并用Cir-cos 0.63 (http://circos.ca/)绘图。
1.4葡萄PLATZ家族基因结构、蛋白保守基序及系统进化树分析
在葡萄基因组数据库(http://www.genoscope.cns.fr)获得葡萄基因组序列、CDS序列及蛋白序列。使用网站GSDS 2.0 (http://gsds.gao-lab.org/in-dex.php)开展外显子-内含子分析。利用在线网站MEME (http://meme-suite.org/)开展葡萄PLATZ家族蛋白保守基序分析,参数如下:高级选项默认,最大基序数7。使用SMART和Pfam数据库对MEME基序进行注释。利用TBtools软件绘制motifs图。利用MEGAX软件对葡萄、拟南芥、水稻、玉米、番茄和大豆共6个物种的PLATZ蛋白序列开展Clust-al W多重序列比对,采用邻接法(Neighbor-joining Tree)构建系统进化树,bootstraps检验(n=1000)。
1.5葡萄PLATZ家族基因啟动子顺式作用元件分析
使用SPDE软件获取VvPLA TZs上游2000 bp启动子序列,利用在线网站PlantCARE(http://bioin-formatics.psb.ugent.be/webtools/plantcare/html/)预测启动子区域顺式作用元件种类及数量。
1.6 VvPLATZ基因有核无核葡萄子代种子发育转录组分析
通过已发表转录组数据(登陆号,PRJ-NA338255),分析VvPLA TZ基因在有核葡萄红地球和无核葡萄森田尼无核杂交分离有核、无核子代种子发育的3个时期(盛花后24、30、39 d)的表达模式,所用材料、时期与已发表文献[19]-致,用TB-tools软件绘制基因表达热图。
1.7总RNA提取与qRT-PCR表达分析
RNA提取使用全能型植物RNA提取试剂盒(CWBIO,Beijing,China),并采用NanoDrop分光光度计(Thermo Fisher Scientific,Shanghai,China)测定RNA的OD值,琼脂糖凝胶电泳检测RNA样品质量。利用ScriptRTase (Trans Gen Biotech, Beijing,China)去除gDNA,使用UEIris Ⅱ RT-PCR System for First-Strand cDNA Synthesis试剂盒(US Ever-bright Inc.)合成cDNA。用于qRT-PCR反应的特异性引物均用Primer 5软件设计,其中VvPLAT24和Vv PLA T213的cDNA序列高度相似,因此共用一对引物,引物序列见表1,Actin(NC_012010)为内参基因。反应体系20μL:上下游引物各1μL,cDNA模板1μL,2×Fast Super EvaGreen qPCR Mastermix 10μL,ddH3O 7μL。设置3次生物学重复和3次技术重复。qRT-PCR设备为LightCycler96(Roche,Shanghai,China),反应程序:95℃120 s;95℃5 s;59℃30 s,45个循环。依据2-ΔΔCT法计算基因的相对表达量,并用Excel 2019进行整理及制图,运用SPSS软件进行显著性分析。
2结果与分析
2.1葡萄PLATZ家族成员鉴定、亚细胞定位预测及同线性分析
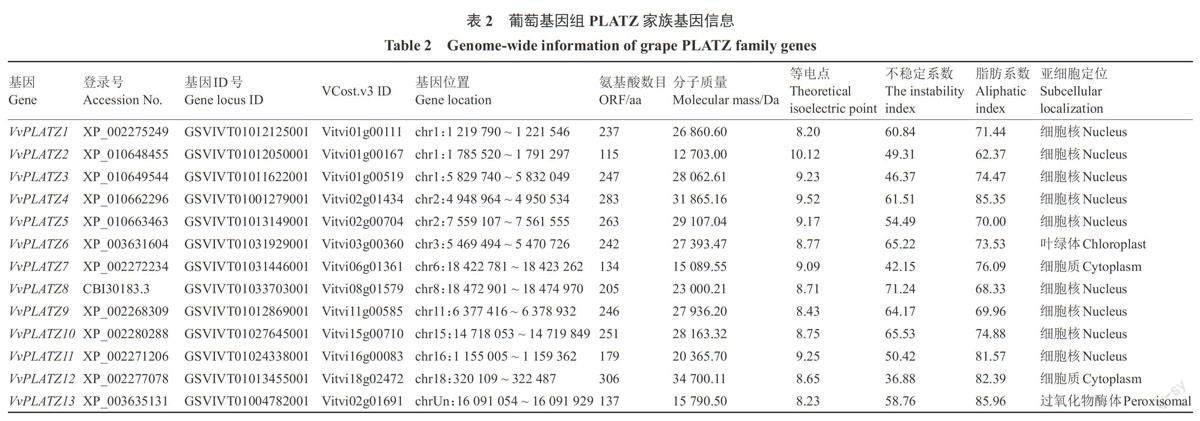
从葡萄全基因组中鉴定出13个PLATZ家族成员,分别命名为VvPLA TZ1~VvPLAT213。由表2可知,葡萄VvPLA TZs基因不均匀地分布在9条染色体上,3个基因在1号染色体,9个基因分别位于2、3、6、8、11、15、16、18号染色体,VvPLAT213基因未知。家族成员氨基酸序列长度为115( VvPLAT22)~306(VvPLAT212)aa,分子质量最大为34 700.11 Da(VvPLAT212),最小为12 703.00 Da(VvPLA T22),等电点为8.20(VvPLAT21)~10.12(VvPLAT22),不稳定系数为36.88(VvPLAT212)~71.24(VvPLAT28),脂肪系数为62.37(VvPLAT22)~85.96(VvPLA T213)。亚细胞定位预测结果表明,葡萄PLATZ家族13个成员中有9个定位于细胞核,2个定位于细胞质,1个定位于叶绿体,1个定位于过氧化物酶体。
根据葡萄自身基因组及葡萄与拟南芥PLATZ家族同线性分析(图1),发现葡萄基因组内Vv PLAT25~VvPLA TZ11和VvPLA T27~VvPLAT28两组并联重复,未发现串联重复。葡萄与拟南芥基因组之间PLATZ家族同线性分析发现,VvPLA T23与AT2G01818存在同线性区域。
2.2葡萄PLATZ家族基因结构及蛋白保守基序分析
为了更好地了解基因进化过程,对PLATZ家族成员的基因结构信息进行分析。系统进化树分析(图2-A)表明,A组PLATZ基因均含有5个外显子,表现出一定结构保守性,其他组PLATZ同组基因外显子一内含子结构具有多样性,例如B组PLATZ基因外显子数目为2~7个不等(图2-B);C组PLATZ基因外显子数目为3~6个不等,D组的VvPLA T22和VvPLA T23分别含有3个和5个外显子。蛋白保守基序分析(图2-C)表明,葡萄PLATZ家族整体上在蛋白保守基序分布模式和数量上呈现较强保守性,均含有PLATZ保守结构域。
2.3葡萄PLATZ家族系统进化分析
为了研究葡萄与其他物种PLATZ家族成员的进化关系,将源于葡萄(13个)、拟南芥(12个)、番茄(21个)、大豆(30个)、水稻(15个)和玉米(15个)的PLATZ蛋白序列开展系统进化树分析(图3),研究发现,来自不同物种的106个PLATZ家族成员可分为5个亚族(A~E),亚族A、B、C、D分别包含2、3、6和2个葡萄PLATZ成员,亚族E不包含任何葡萄PLATZ成员,表现出一定的进化遗失。与单子叶植物水稻与玉米相比,大部分葡萄PLATZ家族成员与双子叶植物大豆、番茄和拟南芥PLATZ亲缘关系更近,例如亚族A的VvPLAT25和SIPLAT219,亚族B的VvPLAT212和AtPLAT25、VvPLAT28和SI-PLAT220,以及亚族D的VvPLAT22和GmPLAT25具有较高的序列相似性,而单子叶植物水稻与玉米PLATZ成员相似性较高。这一现象与单子叶植物与双子叶植物进化上的亲缘关系一致,与单子叶植物相比,双子叶植物之间起源于更近的祖先。
2.4葡萄PLATZ家族基因启动子顺式作用元件分析
不同类型的顺式作用调控元件在基因启动子区域的分布模式一定程度上能揭示基因潜在的生物学功能和调控差异。通过对葡萄PLATZ家族基因启动子区顺式作用元件分析(图4),共鉴定出24种元件,按功能分为4类,激素响应类元件(茉莉酸甲酯、脱落酸、生长素、水杨酸、赤霉素)、光响应类元件、胁迫响应类元件(防御和胁迫、厌氧、干旱、低温、伤口反应)、植物发育调控类元件(胚乳表达、分生组织表达、种子特异性调控、昼夜节律控制、玉米醇溶蛋白代谢)。
除VvPLA T26基因外,其余基因均有激素響应元件,包括茉莉酸甲酯响应元件(CGTCA-motif,TGACG-motif)、脱落酸响应元件(ABRE)、生长素响应元件(TGA-element、AuxRR-core)、水杨酸响应元件(TCA-element)和赤霉素响应元件(GARE-mo-tif、P-box、TATC-box)(图4)。茉莉酸甲酯响应元件最为丰富(24个),脱落酸响应元件次之(23个),含有最多激素响应元件的基因为VvPLA T28(图4),推测大多数VvPLA TZ基因在激素调控途径中具有重要作用。
除VvPLATZ11和VvPLAT25以外,大多数葡萄PLATZ基因均含有胁迫响应顺式元件,如厌氧诱导元件(ARE)、低温响应元件(LTR)、干旱诱导元件(MBS)、伤口反应元件(WUN-motif)和防御与胁迫响应元件(TC-rich repeats)。此外,植物生长发育相关元件(CAT-box、circadian、GCN4_ motif、02-site及RY-elements)的存在表明PLATZ基因很可能参与葡萄生长发育。
2.5葡萄PLATZ家族基因在种子发育过程中的表达模式分析
利用已发表转录组数据,开展VvPLATZs基因在杂交分离有核、无核子代种子发育表达模式分析(图5-A),发现除VvPLAT22、VvPLAT25、VvPLATZ11、VvPLAT212外,其他VvPLATZ基因在有核、无核子代种子发育过程中RPKM表达量均较低或未检测出。与有核子代相比,VvPLAT25、VvPLATZ11及VvPLAT212在无核子代种子发育过程中均显著上调表达;而VvPLAT22在有核子代种子较无核子代上调表达。基于转录组数据,笔者在本研究中进一步选取8个基因在两个有核葡萄(巨峰、醉金香)和两个无核葡萄(火焰无核、克瑞森无核)中开展种子发育qRT-PCR分析,以验证基因表达模式在多个品种中的广谱性。结果表明(图5-B),VvPLAT22和VvPLAT26在有核品种种子发育过程中表达量显著高于无核品种,推测其可能在种子发育过程中发挥作用。与有核品种相比,VvPLAT25在两个无核葡萄品种种子发育过程中表达量均显著高于有核品种,暗示其可能与胚珠败育有关。值得注意的是,VvPLAT22和VvPLAT25基因呈现的差异表达模式几乎涵盖整个发育时期,且差异变化倍数较大。VvPLAT24/13、VvPLATZ11和VvPLAT212在无核品种种子发育前期(20、30 d)表达量均显著高于有核品种。以上qRT-PCR结果与转录组结果基本一致。
2.6 IAA处理后葡萄PLATZ家族基因表达模式分析
通过开展葡萄PLATZ家族基因在IAA处理下的表达模式分析(图6),发现8个VvPLATZs均响应IAA处理,VvPLA T24/13在IAA处理后th响应且表达水平显著大幅度上调,VvPLAT22、VvPLA T26和VvPLATZ10在IAA处理后6h表达水平显著下调,VvPLAT25和VvPLA T212在IAA处理后3和6h表达水平显著上调,且上调倍数较大。因此,推测VvPLAT24/13、VvPLA T25和VvPLA T212基因可能在IAA信号调节途径中起重要作用。
3讨论
PLATZ家族基因系统鉴定已在多种植物中开展,不同物种鉴定到的成员数各不相同。前人在拟南芥、水稻、小麦、番茄、油橄榄和芜青中分别鉴定出12、15、49、21、29和24个PLATZ基因,笔者在葡萄全基因组中共鉴定出13个PLATZ家族基因,推测PLATZ家族基因在植物进化上具有多样性。系统进化树分析发现,6个物种的PLATZ蛋白分为5个亚族,而亚族E不包含任何葡萄PLATZ成员,表现出一定的进化缺失,其余亚族双子叶植物的PLATZ蛋白同源性更高,且VvPLATZ和SIPLATZ蛋白的亲缘关系更密切,推测PLATZ基因在两个物种分化前有共同的双子叶祖先。
在进化上,基因复制事件后重复基因之间存在的功能保守和功能分化基本上取决于3种进化命运:缺失、亚功能化和新功能化。共线性分析结果表明,葡萄PLATZ家族存在两组并联重复基因,无串联重复基因,这与Azim等在芜青PLATZ基因家族的研究结果一致,表明并联重复事件在葡萄PLATZ家族成员扩增过程中起主要作用。并联重复基因VvPLAT25-VvPLATZ11表现出结构和功能的保守性:结构上,这对并联重复基因均含5个外显子,蛋白保守基序分布保持一致,启动子顺式作用元件均仅含光响应元件GTl-motif且无胁迫相关的元件;功能上,并联重复基因VvPLAT25~VvPLATZ11表达模式相似,均在无核品种种子发育过程中较有核品种显著高表达,且IAA处理后3h均显著高表达。
PLATZ基因在胚乳发育等生物学过程中具有重要作用。前人分别在玉米、棉花和水稻中发现PLATZ基因对胚乳发育、养分储存、种子萌发、种子成熟等具有重要作用。笔者在本研究中发现VvPLAT22和VvPLAT26在有核葡萄种子发育过程中表达量显著高于无核葡萄,推测这些基因与种子发育有关,而VvPLAT25在无核葡萄种子发育过程中表达量显著高于有核葡萄,且VvPLAT25具有胚乳表达(GCN4_motif)和种子特异调控(RY-element)相关顺式作用元件,推测该基因与胚珠败育有关。目前鲜见果树PLATZ基因响应激素处理的研究报道,本研究发现大部分葡萄PLA TZ基因响应IAA处理,且VvPLAT24/13、VvPLAT25及VvPLAT212响应幅度可观,推测葡萄PLA TZ家族基因可能通过影响IAA途径调控种子发育过程。本研究首次报道了葡萄PLATZ基因响应IAA处理,填补了此方面研究的空白。笔者在本研究中筛选出一些有核和无核葡萄种子发育差异表达基因,为后续无核分子育种提供基因资源,但这些候选基因的具体生物学功能及调控机制仍需要进一步研究。
4结论
在葡萄全基因组范围内共鉴定到13个PLATZ基因,通过生物信息学分析,将其分为4组,同组家族成员保守基序数目及类别具有保守性,基因结构分析表明除A组外葡萄PLATZ基因在进化上具有多样性。基因表达分析显示PLATZ基因在有核、无核葡萄种子发育不同时期表达模式存在差异且显著响应IAA处理,推测其在葡萄种子发育过程中发挥作用。本研究加深了对葡萄PLATZ基因的认知,为进一步开展葡萄PLATZ基因功能研究奠定了理论基础,有利于为葡萄无核分子育种提供优质基因资源。
猜你喜欢
阅读(低年级)(2020年11期)2020-12-28
女报(2020年10期)2020-11-23
中国教育信息化·基础教育(2016年10期)2016-12-20
今传媒(2016年11期)2016-12-19
电脑知识与技术(2016年27期)2016-12-15
中国科技纵横(2016年17期)2016-11-30
中国中药杂志(2016年20期)2016-11-19
江苏农业科学(2015年5期)2015-10-20
江苏农业科学(2015年9期)2015-10-20
江苏农业科学(2015年8期)2015-09-10
