
胚胎期炎症暴露对中老年海马脑源性神经营养因子基因表达和认知功能的影响
2023-02-24 00:42张折折罗宝灵陈贵海
蚌埠医学院学报 2023年1期
刘 雪,张折折,罗宝灵,陈贵海,梁 冰
衰老相关性空间学习和记忆能力减退是常见的生物学现象,且易受到许多因素的加速,但发病机制尚不完全明确。现已证明海马介导空间学习和记忆,且容易受衰老的影响[1]。海马突触可塑性的改变是衰老相关性学习记忆减退的基础[2]。研究[3]表明神经营养因子,如脑源性神经营养因子(brain-derived neurotrophic factor,BDNF)是突触可塑性的关键调控因子,参与了记忆的形成,而BDNF基因的表达会随着年龄的增长而下降,进而涉及老年期认知损害[4]。证据[5-6]表明胚胎期的不良因素暴露如炎症、铅、营养不良可加速年龄相关性认知损害。我们前期的研究[7]表明,母鼠孕晚期暴露脂多糖(LPS)诱导的炎症会加重子代小鼠中老年期学习和记忆损害。然而,胚胎期炎症暴露是否影响中年期海马BDNF基因表达尚不清楚。
1 材料与方法
1.1 实验动物与处理方法 SPF级2月龄CD-1系小白鼠,雌鼠20只,雄鼠10只,购于北京维通利华责任有限公司。所有小鼠均在标准实验室条件下饲养[室温为(23±1)℃,湿度为55%±5%,明暗周期为12 h,可随意饮食和水]。适应2周后,将雌雄鼠以2∶1的比例合笼,次日清晨检查到阴栓者为受孕第0天。在母鼠怀孕第15~17天腹腔注射LPS(50 μg/kg)或等量的0.9%氯化钠溶液,在出生后第21天,将子鼠与母鼠分开,每笼3~4只同性别子鼠饲养。对应子鼠分别为CON组和LPS组,每组40只,雌雄各半,分别饲养至3月龄、15月龄时,完成行为学实验,随后每组随机抽取6只进行Western blotting和实时定量PCR实验。其中有任何运动障碍、脱毛或可见肿瘤的动物在实验前都被排除在外。所有实验遵从安徽医科大学动物伦理委员会要求(No.LLSC20160165)。
1.2 行为学测试 采用Morris水迷宫实验检测小鼠的学习和记忆功能,该行为学测试在直径150 cm、深度30 cm的黑色圆形不锈钢水池中进行,池中充满水(25 ℃),水深25 cm,分为四个等量的象限(Ⅰ、Ⅱ、Ⅲ和Ⅳ),圆柱形逃生平台(直径10 cm)隐藏在第一象限中心的水下1 cm处,水迷宫周围有一层不透明的白色窗帘,在距水面1 m的位置悬挂3个不同的黑色标志物,来帮助小鼠获取位置信息。该实验分为两个阶段:定位航行期(学习期)和空间探索期(记忆期)。将小鼠从不同象限且面向侧壁入水,允许其自由游动60 s,若找到逃生平台,则允许该小鼠在平台上停留30 s,然后开始下一次实验。未能在60 s内找到平台的小鼠从水池中被救出,并在下一次实验开始前在平台上放置30 s。每天4次,每次间隔15 min,连续7 d。待第7天学习期结束后,开始进行记忆期实验,撤去平台,将小鼠从平台相反的象限开始,自由探索60 s。Any-maze软件记录小鼠找到平台的游泳路程(代表学习能力)以及小鼠在靶象限(平台所在象限)内的游泳路程占总路程的百分比(代表记忆能力)。
1.3 组织制备 Morris水迷宫任务完成后第15天,将小鼠颈椎脱臼处死迅速取脑,冰上取海马组织后放置-80 ℃冰箱冷冻保存。
1.4 Western blotting分析 将海马组织解冻后,将其充分研磨,加入适当的RIPA细胞裂解液,高速离心,裂解组织,收集上清液。往上清液中按1∶4比例加入SDS-PAGE 蛋白上样缓冲液。水浴(100 ℃)加热10 min,使组织充分变性,待样品冷却至室温后,直接加到SDS-PAGE胶加样孔内,浓缩胶100 V,时间为30 min;分离胶电压100 V,时间为60 min。恒流转模后,继续漂洗5 min,加入Western 封闭液,室温条件下封闭2 h,加入一抗兔抗-BDNF(1∶1 000,ab108319,美国Abcam公司),4 ℃孵育过夜后使用PBST洗涤。加入二抗羊抗兔IgG (1∶20 000,Zsbio,ZB2301),室温孵育2 h后继续PBST洗涤,随后用发光试剂盒来检测蛋白。使用Image J 软件对蛋白条带的结果进行分析,计算BDNF的相对表达量。
1.5 实时定量PCR 称取海马组织50 mg,剪碎,液态研磨,加入1 mL TRIzol进行裂解,提取RNA组织,并检测其纯度。将RNA经RT反应,逆转录生成cDNA,取cDNA作为荧光定量的模板。反应体系如下,5 μL 2× SYBR Green Mixture、1 μL Forward primer(10 μmol/L)、1 μL Reverse primer (10 μmol/L)、1 μL cDNA、2 μL RNase Free water。反应条件如下,Hold(预变性):95 ℃、60 s、单循环;PCR反应:95 ℃、20 s,60 ℃、60 s,共40循环。本次实验所使用的计算方法为2-△△Ct。引物序列见表1。
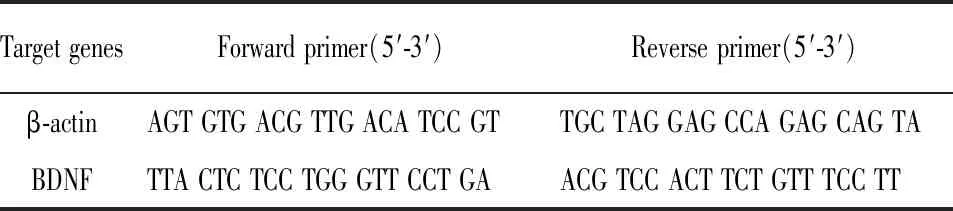
表1 引物序列
1.6 统计学方法 所有数据均使用SPSS25进行统计分析,所有资料均符合正态分布。Morris水迷宫成绩用重复测定方差分析及双因素方差分析进行统计。其中,组间数据分析采用LSD检验。海马BDNF基因表达水平与Morris水迷宫结果之间的相关性用Pearson 相关进行分析。
2 结果
2.1 学习期 CON组小鼠游泳路程随时间的增加而逐渐降低[F(6,216) = 35.968,P<0.01]。15月龄比3月龄CON组游泳路程明显延长[F(1,36)=10.949,P<0.01],但性别及性别与年龄的交互效应均无统计学意义(P>0.05)(见图1A)。在3月和15月龄,LPS的处理效应均显著[F(1,36)=6.016,P<0.01;F(1,36)=9.925,P<0.01],即在两个年龄段,LPS组的游泳路程均明显长于CON组,但性别及性别与处理的交互效应均无统计学意义(P>0.05)(见图1B、C)。
2.2 记忆期 15月龄CON组小鼠靶象限游泳路程百分比明显低于3月龄CON组[F(1,36) =16.950,P< 0.01],但两个年龄段性别差异均无统计学意义(P>0.05)。在3月龄和15月龄,LPS组均比CON组小鼠靶象限游泳路程百分比均明显降低[F(1,36)=8.139,P<0.01;F(1,36) =17.480,P<0.01],各组性别效应无统计学意义(P>0.05)(见图1D)。

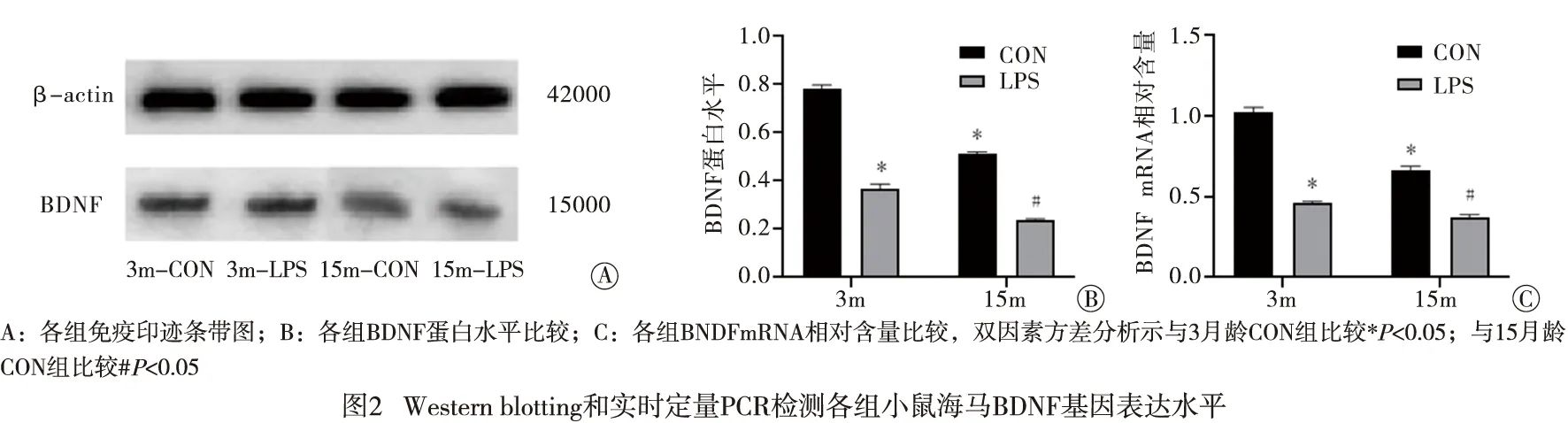
2.3 海马BDNF基因表达水平 各组免疫印迹条带图见图2A。在CON组中,15月龄较3月龄小鼠海马BDNF蛋白和mRNA相对含量均明显降低[F(1,20) = 203.300,P<0.01;F(1,20)=86.430,P<0.01],但性别效应无统计学意义(P>0.05)。在3月龄和15月龄,LPS组较CON组小鼠海马BDNF蛋白[F(1,23)=233.764,P<0.01;F(1,23)=706.340,P<0.01]和mRNA[F(1,23)=335.422,P<0.01;F(1,23)=93.868,P<0.01]相对含量均显著降低,各组性别效应无统计学意义(P>0.05)(见图2B、C)。
2.4 小鼠空间学习记忆能力与海马BDNF基因表达水平的相关性 Pearson分析表明,15月龄CON组和LPS组小鼠海马BDNF蛋白水平与平均游泳路程呈负相关关系(r=-0.649,P<0.05;r=-0.871,P<0.01),与靶象限游泳路程百分比呈正相关关系(r=0.780,P<0.01;r=0.840,P<0.01);小鼠海马BDNF mRNA相对含量与平均游泳路程呈负相关关系(r=-0.891,P<0.01;r=-0.582,P<0.05),与靶象限游泳路程百分比呈正相关关系(r=0.883,P<0.01;r=0.712,P<0.01)。3月龄海马BDNF蛋白和 mRNA相对含量与平均游泳路程和靶象限游泳路程百分比均无明显相关关系(P>0.05)(见表2)。
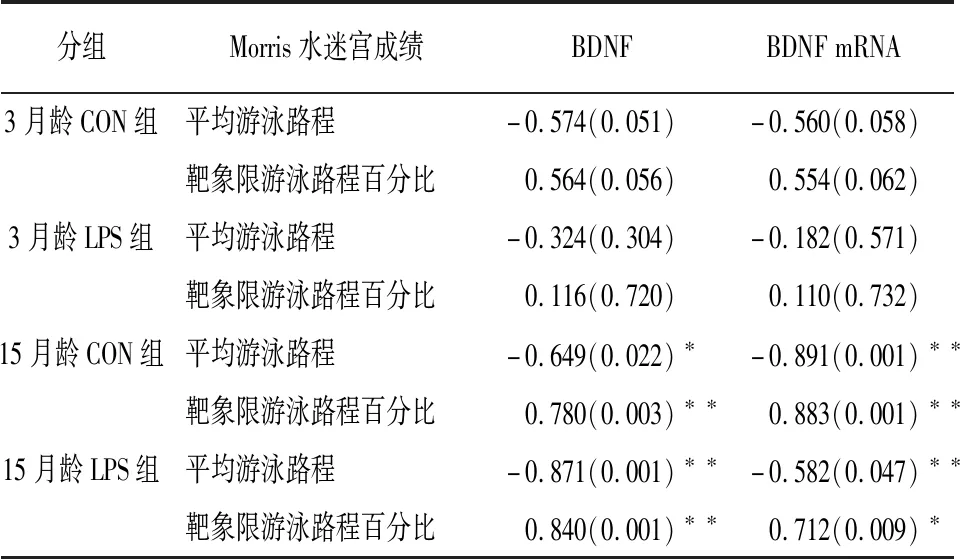
表2 海马BDNF基因表达水平与Morris水迷宫成绩的相关性(r)
3 讨论
衰老是伴随认知能力下降的正常生理过程,是老年相关疾病如阿尔茨海默病发展的主要危险因素[8]。虽然在阐明与年龄相关的神经病理学和影响认知下降的因素之间的复杂关系方面取得了相当大的进展,但导致认知功能障碍的具体机制目前并不明确。根据成人疾病的胎儿起源假说[9],大脑发育的关键窗口期易受应激的影响,并可能导致永久性的不良后果。其中,孕产妇感染是常见的应激源,可对胎儿的发育产生潜在的危害。动物研究[10]表明,母体感染可引起胎儿全身炎症反应,表现为胎儿或宫内环境中促炎症细胞因子的升高,进而影响胎儿的认知功能。研究[7]发现,在孕期给予母鼠腹腔注射小剂量LPS,虽然不会导致胎儿的早产和死亡,且可正常发育,但中老年期会出现空间学习记忆功能的提前减退。本研究中,15月龄CON组比3月龄CON组学习期游泳路程延长(P<0.01),记忆期靶象限游泳路程百分比降低(P<0.01),且两个年龄段的炎症暴露组比对照组成绩均差。这些结果提示青年期和中老年期LPS组小鼠学习记忆能力减退。然而,我们关于青年期的结果似乎与已出现的证据[11]矛盾,这可能与本研究样本量大于已有的研究而使统计学把握度增加所致。
研究[12]表明,认知功能障碍与神经元的突触可塑性受损有关。BDNF作为调节突触可塑性和记忆形成的主要驱动因子,与酪氨酸激酶受体B结合,在认知形成过程中发挥关键作用[13-15]。本课题组及其他小组的研究表明,BDNF通过介导海马长时程增强作用来参与学习与记忆的过程[16-17],而上调海马BDNF mRNA转录水平可改善大鼠的记忆障碍[18]。此外,端脑受损BDNF突变小鼠表现出显著的空间学习缺陷[19]。人类退行性疾病中,包括阿尔茨海默病、帕金森病、亨廷顿病的认知功能障碍可部分归因于BDNF蛋白及mRNA转录水平的减少[20]。BDNF的表达水平易受产前和产后应激的影响。例如母子分离可导致大鼠成年后海马BDNF表达水平的降低,其影响主要局限于成熟形式的BDNF[21]。而在本研究中,胚胎期炎症暴露可使成年小鼠海马BDNF表达水平下降。另一项研究[22]显示,在母鼠怀孕期间给予温和性刺激可导致Lewis雄性大鼠成年期BDNF蛋白水平降低和mRNA转录的增加。这种差异可能是由不同品系的啮齿类动物和实验操作导致的。Pearson相关分析显示,15月龄CON组和LPS组小鼠海马BDNF表达水平与学习期游泳路程呈负相关关系,与记忆期靶象限游泳路程百分比呈正相关关系。因此我们推测海马BDNF表达水平降低可能与年龄相关性空间学习记忆减退有关。
综上,胚胎期炎症暴露可导致小鼠认知功能减退及海马BDNF基因表达水平降低。本研究的不足之处在于未采用转基因技术过表达或敲除BDNF,观察小鼠在青年期和中老年期认知功能改变情况。且到目前为止,我们仍不清楚BDNF基因表达情况对胚胎期炎症暴露引起的认知功能障碍的贡献程度。未来的研究将对BDNF基因参与认知功能调控的具体机制作进一步探索。
猜你喜欢
现代食品科技(2022年8期)2022-09-02
中学生数理化·高一版(2022年3期)2022-04-05
中学生数理化·七年级数学人教版(2022年3期)2022-03-16
初中生学习指导·提升版(2020年10期)2020-09-10
数学小灵通·3-4年级(2020年6期)2020-06-24
科学生活(2019年7期)2020-01-01
养殖与饲料(2019年10期)2019-02-25
小学生学习指导(高年级)(2019年6期)2019-01-11
发明与创新·小学生(2018年12期)2018-12-29
山东畜牧兽医(2018年3期)2018-04-26
