
优质肉鸡苏禽3号母系经济性状遗传参数的世代变化规律
2023-02-15 01:44王钱保黄华云刘宏祥李春苗黄正洋黎寿丰赵振华
江苏农业科学 2023年1期
王钱保, 黄华云, 刘宏祥, 李春苗, 黄正洋, 黎寿丰, 赵振华
(江苏省家禽科学研究所,江苏扬州 225003)
苏禽3号母系L系是江苏省家禽科学研究所自主培育黄羽优质肉鸡新品系,该品系具有体型适中、胴体美观、肉质风味佳、抗逆性强等优点,但也存在诸如生长发育不均、饲料报酬偏低、性成熟启动不一导致上市日龄参差不齐、产蛋高峰期短等缺点。肉鸡体质量、性成熟的启动至开产性状一直被认为是肉鸡育种工作中最重要的经济性状,而优质肉鸡有别于一般快大型肉鸡,其肉质及开产后的高峰产蛋数、维持时间及蛋品质也被划入成重要的经济性状。苏禽3号L系多项指标生长发育潜力的不足使得养殖成本大大增加,不仅严重降低了优质肉鸡的经济价值和市场竞争力,同时也阻滞了其后续开发和利用。因此,为了全面提升苏禽3号L系优质肉鸡的经济性能,最佳的措施之一便是依靠精准、可靠的遗传参数估计数量性状变异,合理制定最适宜的生产育种方案。
作为肉鸡生产中重要的经济性状,生长、上市日龄(性成熟启动)、肉质等已被大量学者进行细致研究,其中,基于传统系谱的最佳线性无偏预测(BULP)、基因组最佳线性无偏预测(GBULP)及一系列其他动物统计分析模型方法对优质肉鸡不同生长阶段体质量、胴体性状和肉质等遗传力及彼此间遗传关系进行估计较为丰富,但这些研究得出的结果却不尽相同,遗传力与遗传相关估值存在较大差异[1]。现有文献报导,优质肉鸡前期体质量与后期产蛋数遗传力统计范围在0.32~0.57与0.10~0.47间不等,如此大的数值跨度也导致遗传相关性的方向并不统一[2-3],正向[4]或反向[5]推论也使得遗传评估工作变得极为困难,致使育种工作者难以选择合适指数参数进行有效且针对性的育种规划。优质肉鸡的多项经济性状由众多基因综合调控[6-7],品种、环境、营养等因素也能影响表观修饰进而调控基因的表型效应[8],此外,统计方法和标准的不同也会影响各性状遗传力的比较与加权[9]。因此,遗传基础的复杂性加上统计方法的多样性决定了研究结果的异同性。
本研究综合参考优质肉鸡经济性状遗传复杂性,通过参照、发掘、利用前人研究结果,以优质肉鸡苏禽3号S3品系2018—2021年度育种核心群 1 603 羽一世代个体为对象,记录8、12、16、20、33、43周龄体质量、开产体质量、开产蛋质量及33周龄和43周龄产蛋数,估测苏禽3号L系各经济性状遗传参数并分析性状间的遗传关系,以期为优质肉鸡经济性状的选育奠定理论基础。
1 材料及方法
1.1 试验材料
2021年7月2日,试验选取江苏省家禽科学研究所培育品系苏禽3号优质肉鸡L系核心育种群近一世代(2020—2021年度)2 100羽母鸡家系资料,鸡只被饲养在江苏省家禽科学研究所邵伯基地试验禽场内。这些个体来自60个半同胞家系,该世代在2020年5月25日入孵,6月15日出雏,每个个体有完整的系谱记录。试验地点为江苏省家禽科学研究所肉鸡育种与生产科室养殖基地。育雏期均网上平养,育成期后10周龄上笼,统一执行优质肉鸡一般免疫程序和光照处理,全期自由采食饮水,营养需求按优质肉鸡常规管理,基础饲粮组成及营养水平见表1。

表1 基础饲粮组成及营养水平(风干基础)
1.2 数据采集处理和统计分析


表2 苏禽3号L系各性状的缩略词表
2 结果与分析
2.1 优质肉鸡苏禽3号L系各经济性状的统计描述
优质肉鸡苏禽3号L系不同时期体质量性状见表3。由表3可知,0~12 周龄的变异系数超过5%,其中,0周变异系数相对较小,此时公母鸡体质量变异系数在6%以下,4周龄后逐渐加大,16周龄后公母鸡分别达到7.84%、7.64%后逐渐降低,待开产后平稳减缓,后期33~43周后继续降低一定数值后趋于平稳,表明苏禽3号优质肉鸡L系前期体质量增长幅度较大,生长不均,相较于后期较为平稳的生长发育状况仍有很大的提升空间。16周龄公鸡体质量比母鸡高了近30%,因此,性别对苏禽3号优质肉鸡L系体质量性状的影响较大,提示公鸡比母鸡具有更强的生长潜力。
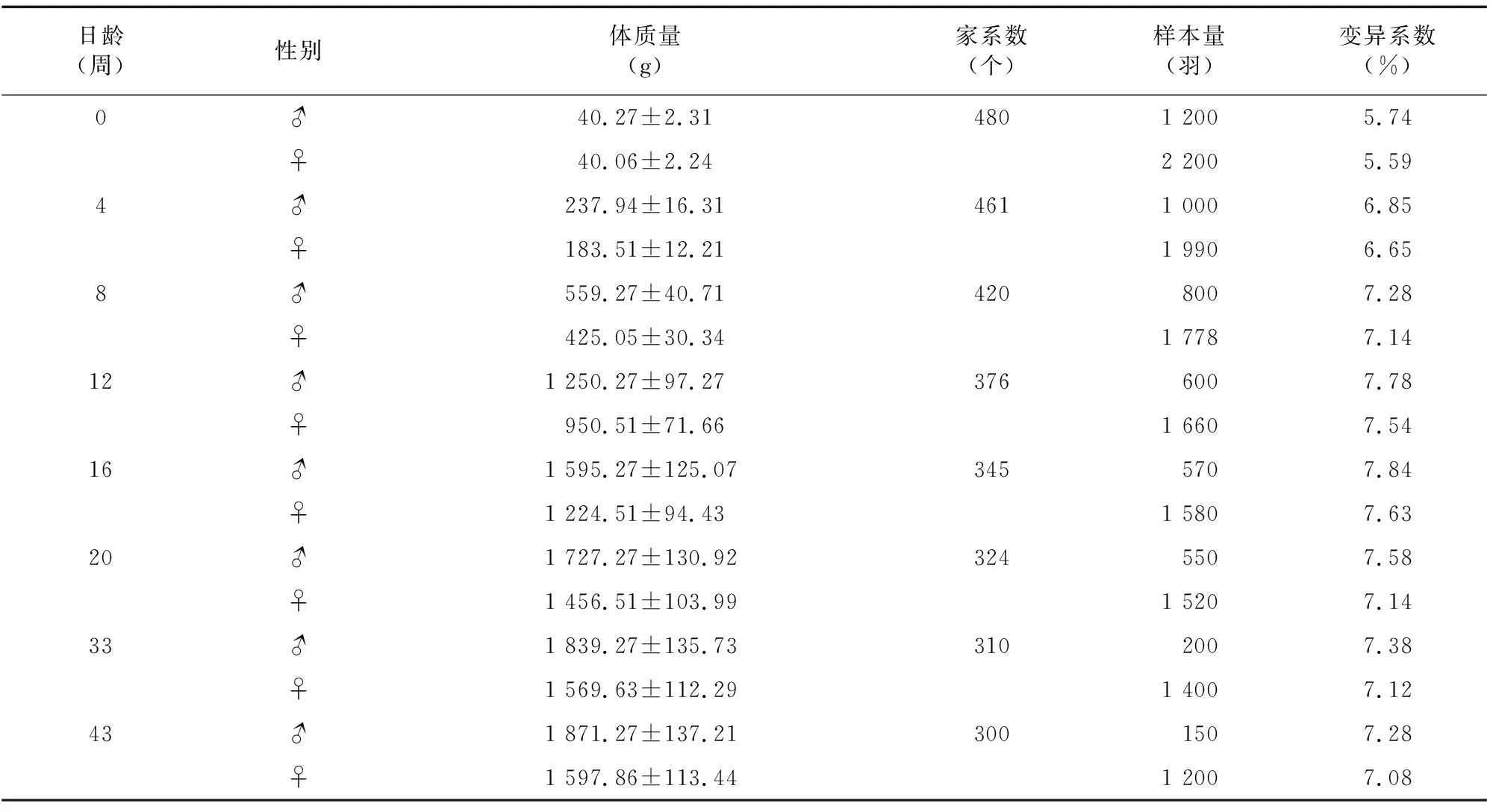
表3 苏禽3号L系不同时期体质量性状统计描述
优质肉鸡苏禽3号L系开产和产蛋性状见表4。由表4可知,除开产蛋质量外,优质肉鸡苏禽3号L系各指标的变异系数超过7%,其中,33周产蛋数和43周产蛋数变异系数较大,超过10%,表明鸡群开产时体质量、日龄不齐整,前期体质量和高峰期、产蛋后期产蛋数的整齐一致性较差,仍有很大的选育提升空间。

表4 苏禽3号开产和产蛋性状统计描述
2.2 优质肉鸡苏禽3号L系各经济性状遗传力估计及遗传与表型相关
优质肉鸡苏禽3号L系各经济性状遗传力估计及遗传与表型相关见表5。由表5可知,0、4、8、12、16、20周体质量遗传力为0.364 0~0.554 9,开产体质量遗传力为0.440 8,均属中高遗传力;33、43周体质量遗传力为0.143 0、0.157 1,属低遗传力,说明L系产蛋中后期饲养管理和环境因素对体质量的影响作用更大。开产蛋质量的遗传力为0.154 8,33周龄产蛋数和43周龄产蛋数的遗传力分别为 0.137 8、0.126 7,均属低遗传力。各时间点体质量均表现出正遗传相关性,遗传相关系数在0.142 4~0.938 5间,随体质量测定间隔的延长而减弱。开产体质量与20周龄、33周龄体质量均呈现出较强的遗传相关性(0.898 4和0.709 9),与4周龄、16周龄遗传相关性较弱(0.369 8和0.376 8)。各体质量间的表型呈较高的相关性,略低于遗传相关,同时变化规律与遗传相关相似。33周龄产蛋数和43周龄产蛋数的遗传相关达0.923 4,表明产蛋性状调控基因作用较为恒定。两者对应的表型相关也达0.836 7,表明L系在今后产蛋数为目标的选种过程中,可选择33周龄产蛋数作为目标性状,尽快缩短育种年限,加快育种总体选育进程。产蛋数和体质量间的遗传及表型相关均较低,表明产蛋数和体质量在选育过程中应区分开独自选育,同时合理选择遗传相关性较高的性状从而更好地提高育种效率。

表5 苏禽3号不同性状遗传力及遗传相关
3 讨论
近些年,家禽群体质量等经济性状的遗传进展在实际育种方案实施过程中备受关注,完整且系统的系谱记录是各经济性状获得精准遗传参数进而取得重大遗传进展的前提[10]。优质肉鸡占据了鸡肉消费的半壁江山,根据美国农业部数据推测2022年中国黄羽优质肉鸡的消费水平仍保持旺盛上升趋势。本研究针对多个不同时期体质量、产蛋数及开产性状遗传力及各性状间的遗传相关进行估计,阐明了苏禽3号群体遗传特性进而为后期的育种工作提供科学技术指导。
3.1 优质肉鸡苏禽3号各经济性状遗传力
肉鸡的体质量、开产及产蛋性状作为肉鸡育种工作中最重要的经济性状,肉鸡体质量性状遗传力受品种[11]、生长日龄[12]及饲养环境[13]等因素影响,此外,统计方法的不同也会影响遗传力的精准评估[1,14]。优质肉鸡的生长曲线表明,前期生长缓慢,出现拐点一般为6~8周龄左右,略迟于一些地方鸡,而比国外快大型肉鸡生长拐点明显更迟[15],16周龄后生长逐渐变缓,性成熟开始启动[16]。郭军等研究表明,鸡的体质量属于中等遗传力性状,随着日龄生长趋势在不断变化,不同阶段的遗传力也随之变化[17]。Singh等研究发现,火鸡随着日龄的增加遗传力随之增加[13],而Saatei等在探究日本鹌鹑的体质量遗传特性时发现,体质量遗传力随着年龄增长而降低,表明遗传力随年龄增加母体效应的影响混杂在加性效应中,从而提高了遗传力[18]。Tongsiri等发现,泰国地方鸡中前期遗传力随着周龄增长,但16周后逐渐降低,至24周时趋于稳定[5]。本研究发现,苏禽3号优质肉鸡L系体质量遗传力0和4周龄体质量时为0.554 9和0.523 7,属中高遗传力,由此可见早期母体效应对体质量遗传力影响较大,随着年龄的增长遗传力逐渐降低,很有可能是因为母体效应的逐渐降低;这与Saatei等的研究结果[18]一致,与Singh等的研究结果[13]表现出相反的变化趋势,原因可能是在不同品种中其生长性能差异较大导致遗传方差有所不同,同时也表明随着日龄增长,环境效应对体质量的影响逐渐加大,提示个体表型选择应在早期进行,才可取得良好的选育效果;对于20周龄以上的体质量性状选育应充分考虑环境对表型性状的影响,个体育种值、家系选育可能成为更好的选育方案。
产蛋性能作为家禽生产中的一个重要性状,其遗传参数估计结果均较低[19]。相比于蛋鸡,优质肉鸡的经济价值在于肉,体质量生长及上市的胴体性能是其关注的重点。但有别于一般肉鸡,优质肉鸡母鸡性成熟启动直至开产后,其蛋品质、高峰期产蛋数及维持天数依然具有可观的经济价值,如何最大化利用优质肉鸡的产蛋性能成为今后选育的重点。郭军等研究表明,遗传背景对产蛋数遗传力影响有限,推测群体特异性对产蛋数遗传力有重要影响。此外,产蛋数遗传力随周龄变化而变化,产蛋后期遗传力较低[20],推测可能是因为产蛋后期生理机能趋于退化,遗传调控功能减弱[21]。党李苹等研究发现,略阳乌鸡31、35、40周龄产蛋数遗传力在0.25~0.27间,变化趋势亦不明显,表明周龄对产蛋数量性状基因位置效应影响较小,产蛋数性状受遗传调控在进化上较为保守,在时间上较为恒定[22]。本研究发现苏禽3号33、43周龄产蛋数遗传力分别为0.137 8和0.126 7,后期遗传力略低于前期,两者均属于低遗传力。开产性能作为家禽另一个重要的经济性状,受前期的生长发育和性成熟启动影响,同时也影响早期和高峰期产蛋数,本研究估计的开产日龄遗传力为0.147 6,稍低于如皋黄鸡的0.22和白耳黄鸡的0.27[23-24],作为早熟性状在开展个体选择时未能起到很好的选育效果。可能原因是本研究群体持续对产蛋数开展定向选择,促进了有利基因的纯合,遗传方差的降低导致性状遗传力变小。开产日龄和2个时期产蛋数的低遗传力表明,这些性状在很大程度上受饲养管理和环境等因素所制约,L系作为当下屠宰型优质肉鸡消费生力军,如何进一步提升其经济性能,对于低遗传力性状直接进行个体选择效果不佳,还须通过家系选择、家系内选择等方法,兼顾配套系杂交等模式,充分利用杂种优势有望取得良好的选育效果。
3.2 优质肉鸡苏禽3号各经济性状间的遗传与表型相关
Tongsiri等对泰国地方鸡8、12、16、20周龄及开产5个时期体质量间的遗传相关进行了估计,范围在0.55~0.99间[5];Niknafs等评估了伊朗本地土鸡8周龄、12周龄、开产3个时间点体质量间的遗传相关,范围在0.36~0.91之间,以上研究体质量间的遗传相关性均随着周龄间隔的增大而降低[25]。在本研究对苏禽3号从0至43周龄及开产时共10个时间段体质量间的遗传相关进行估计,发现遗传相关系数在0.142 4~0.938 5间,与上述报道变化规律基本一致,表明体质量间的正向遗传相关性对体质量早期选择起着关键作用,根据早期体质量对后期体质量进行选择具有可行性,但选择的准确性随着时间的跨度延长而降低,因此,在今后育种选择过程中,建议合理选择与之遗传相关性较高的早期体质量进行选择从而提高育种效率。
体质量与产蛋数性状间负相关研究结果较多,虽然对肉鸡的选育造成困扰,但是也恰好弥补了体质量对产蛋数性状的影响[26]。本研究产蛋数和体质量间的遗传相关均较低,相关系数不超过0.25,结果与党李苹研究的体质量与产蛋数的遗传相关属于弱的正相关结果[27]相似。推测仅有在生长强度超过一定阈值时,体质量与产蛋数性状间才呈现负遗传相关,低于这个阈值时生长与繁殖是相辅相成,健康的体质量生长会为后期产蛋性能的发挥提供坚实的体况基础。多数研究表明,开产、产蛋数性状与体质量等性状之间的遗传相关性较高[11,19]。开产日龄作为早熟性的一个经济指标和选种参照,不仅对优质肉鸡性成熟和上市具有重要意义,同时对优质肉鸡的选种方案规划有很高的指导性。蛋质量作为优质肉鸡育种上的一个重要指标,种蛋大小至关重要,偏小则蛋壳致密,蛋黄和蛋白等内容物比例不均,后期孵化效果差。王钱保等研究表明,开产蛋质量正向影响后期蛋质量,提高开产蛋质量的意义比提高中后期(33~43周龄)的蛋质量意义更大,因此选择开产时的蛋质量更为重要[28]。本研究中,苏禽3号L系开产性状与其他各性状间存在不同程度正负相关性,因此可加以充分利用,建议在群体当前的生长强度下,可同时对体质量性状、开产性状和产蛋数性状共同进行选育。
4 结论
苏禽3号优质肉鸡L系前期0~20周龄体质量属于中等遗传力,后期33、43周龄体质量遗传力降为低遗传力,表明遗传效应在L系生长调控方面起关键作用,对L系在12周龄以前个体体质量表型选择有望取得良好的选育效果,16周龄后应充分考虑对饲养管理、环境的综合控制。L系开产日龄和33、43周龄产蛋数遗传力较低,直接选育对提高产蛋数效果不理想,应加强持续产蛋力选育。体质量和开产、产蛋数表现出遗传和表型同趋势的正相关性,表明遗传和环境效应呈现一致性。各性状间的低相关性说明组合性状同步选育难度较大,而体质量间的高相关性提示合理选择高遗传相关性状可提高育种效率。
猜你喜欢
动物营养学报(2022年11期)2022-12-08
养猪(2022年4期)2022-08-17
特产研究(2021年6期)2021-12-08
21世纪(2019年12期)2019-12-18
河南农业科学(2019年9期)2019-09-24
小哥白尼(野生动物)(2018年9期)2018-12-08
湖南农业(2017年1期)2017-03-20
农村百事通(2016年6期)2016-05-14
湖北畜牧兽医(2015年11期)2016-01-11
山东农业科学(2014年1期)2015-03-09
