
大熊猫栖息地森林群落乔木层与林下冷箭竹关系研究
2023-02-13 12:31秦炜锐宋心强付明霞刘景怡张远彬周材权
西华师范大学学报(自然科学版) 2023年1期
秦炜锐,潘 翰,程 勇,宋心强,付明霞,刘景怡,张远彬,周材权,杨 彪
(1.西华师范大学 a.栗子坪大熊猫生态与保护四川省野外科学观测研究站,b.生态研究院,四川 南充 637009;2.北京市企业家环保基金会,北京 100020;3.荥经县大相岭自然保护区管理局,四川 雅安 625000;4.中国科学院、水利部成都山地灾害与环境研究所,成都 610041)
大熊猫(Ailuropodamelanoleuca)是全球生物多样性保护的旗舰物种与伞护物种。过去几十年大熊猫保护取得了举世瞩目的成就,但该物种仍然面临繁殖力弱、存活率低、栖息地破碎化等问题[1]。根据全国第四次大熊猫调查的结果,仍有30%的大熊猫栖息地质量有问题[2]。由于自然隔离和人为干扰,大熊猫栖息地被隔离成33个斑块,面积小于10 000 hm2的斑块有9个,栖息地破碎化将给野生大熊猫种群的生存、繁衍带来威胁[3]。大相岭山系有大熊猫38只[2],是六大山系中种群数量最少的[4],该区域栖息地面积小、破碎化程度高、人类活动干扰严重,亟待加强保护[4]。伴随大熊猫国家公园的建立,栖息地恢复工作是未来大熊猫国家公园保护管理工作的重点。
大熊猫是森林类型的物种,竹类资源、森林起源以及遮蔽条件是影响大熊猫生境利用的主要因素[5]。森林中的乔木为大熊猫提供躲避、休息以及巢穴,林下竹子作为大熊猫主要的食物来源,是大熊猫生存的基础。已有研究表明,森林群落乔木层郁闭度对林下植物的生长起重要作用[6-9],乔木是森林群落恢复过程中重要的生物因子[10],乔木层通过对光的透射、反射和吸收,直接影响林下光照的强度和分布,而光照对于绝大多数森林树种而言,是决定更新个体能否生存和生长的关键[11]。乔木层郁闭度对竹子的高度、基径和密度等具有重要的影响[7,12]。因此,研究乔木层结构特征与林下竹子生长指标之间的关系是栖息地恢复的基础,对提升大熊猫栖息地质量、保护大熊猫及全球生物多样性具有重要的作用。
基于此,本研究选择大相岭山系的大熊猫栖息地作为研究对象,针对大熊猫栖息地森林群落乔木层结构特征与林下冷箭竹(Bashaniafaberi)生长指标开展调查,定量分析两者之间的关系,并建立大熊猫栖息地森林群落乔木层特征与林下冷箭竹生长关系的模型,以期为该区域大熊猫栖息地恢复提供量化依据。
1 研究方法
1.1 研究区概况
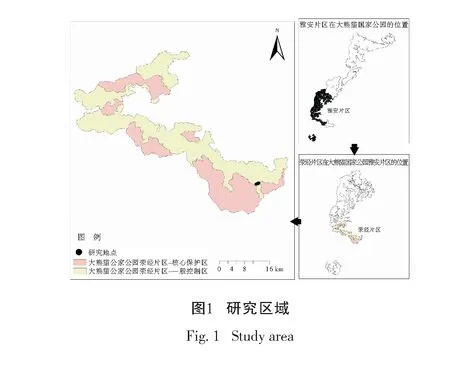
样地设置在大相岭大熊猫野化放归基地周边原生森林生态系统林下分布有冷箭竹的区域(图1)。该区域位于大熊猫国家公园荥经片区,海拔2 363~2 429 m,气温15.0~17.6℃,年降水量1 200~1 800 mm,属亚热带高山山地气候。植被类型为以冷杉(Abiesfabri)、槭树(Acerspp.)等为主要树种的针阔混交林,林下以冷箭竹为主,是理想的大熊猫栖息地。
1.2 野外调查
调查于2021年5月进行。设置4条样线,在样线上随机布设20 m×20 m的乔木样方共98个,记录样方的海拔(m)、坡度(°)、坡向(°)、坡位(背坡、山肩、山顶、麓坡、趾坡)、坡型(均匀坡、复合坡、凹坡、凸坡)、植被起源(原始林和次生林),以及乔木层郁闭度、乔木平均胸径、乔木平均高度等。每个乔木样方内分别按“品”字型获取3个1 m×1 m的冷箭竹样方,共计294个冷箭竹样方,计数每个样方中冷箭竹发笋量和竹子总株数,测其基径和高度。
1.3 数据处理方法
首先对野外调查所得的294个冷箭竹样方进行初步筛选,剔除6个数据不完整的样方,最终保留288个样方数据进行建模。将乔木郁闭度划分为4个级别:0~0.25,0.26~0.50,0.51~0.75,0.76~1.00;将乔木平均高度划分为3个级别:≤10 m,11~20 m,21~30 m;将乔木株数划分为5个级别:1~10株,11~20株,21~30株,31~40株,>40株;将乔木平均胸径划分为3个级别:粗(>70 cm),中(35~75 cm),细(<35 cm)。
利用R软件进行单因素方差分析,以检验乔木层结构特征对冷箭竹生长指标的影响,然后进行逐步回归分析,选取显著性最高的因子建立估测模型,根据模型评价指标,选择最优回归模型。由于冷箭竹平均高度和平均基径是连续型变量,因而考虑环境因子的随机效应,构建线性混合效应模型[13]:Yij=a0+a1Xij+bi+eij,其中,Yij表示第i类的第j个观测的反应变量,a0表示固定截距,a1表示固定斜率,Xij表示固定效应,bi表示随机效应(只影响截距),eij表示残差;冷箭竹无性系种群数量(包括发笋量和株数)是离散型变量,因而选择泊松分布形式,考虑环境因子(海拔、坡度、坡向、坡位、坡型、植被起源)的随机效应,构建广义线性混合模型[14-16]:g(μij)=ηij=Xijβ+Zijμi,其中,g(·)表示联结函数,μij表示条件均数(考虑了海拔、坡度、坡向、坡位、坡型、植被起源的随机效应),ηij表示条件线性预测值,Xij表示解释变量,β表示固定效应参数矢量,Zij表示与随机效应相关的解释变量,μi表示随机效应。
2 结 果
2.1 冷箭竹及乔木层调查结果
结果显示(表1):每个冷箭竹样方内冷箭竹总株数为(28.88±10.37)株,发笋量为(6.56±5.40)株,平均高度为(83.63±29.92) cm,平均基径为(5.89±1.76) mm;每个乔木样方内乔木层郁闭度为(0.42±0.26),株数为(11.14±8.15)株,平均高度为(12.32±7.05) m,平均胸径为(28.63±22.40) cm。

表1 冷箭竹及乔木层调查结果(平均值±标准差)
2.2 乔木层结构特征与冷箭竹生长指标的关系
单因素方差分析的结果表明(表2),不同乔木层郁闭度下冷箭竹平均高度差异极显著(F3,284=13.28,P<0.01),冷箭竹株数差异显著(F3,284=3.72,P=0.012<0.05);不同乔木株数下的冷箭竹株数和平均高度差异极显著(F3,284=13.02,P=<0.01;F3,284=7.98,P<0.01);不同乔木平均高度下的冷箭竹平均高度和平均基径差异极显著(F2,285=13.26,P<0.01;F2,285=9.8,P<0.01),冷箭竹发笋量差异显著(F2,285=3.77,P<0.05);不同乔木平均胸径下的冷箭竹株数、平均高度和平均基径差异极显著(F2,285=4.92,P<0.01;F2,285=15.63,P<0.01;F2,285=12.23,P<0.01;),冷箭竹发笋量差异显著(F2,285=3.50,P=0.032<0.05)。
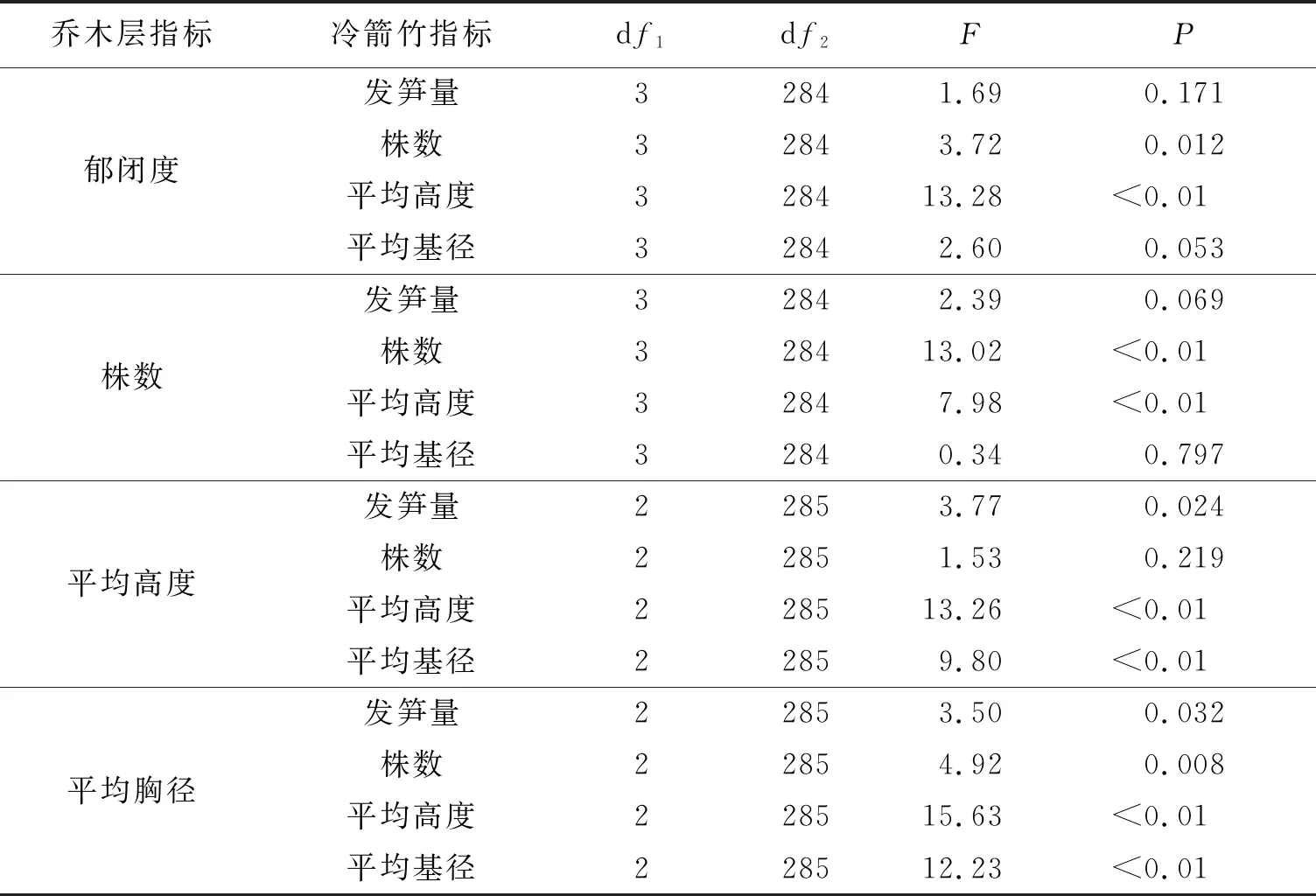
表2 乔木层结构特征不同组别的冷箭竹生长指标方差分析
2.3 模型构建
根据方差分析结果,挑选对冷箭竹生长指标有显著影响的因子作为模型的固定效应。因此,可将乔木平均高度和平均胸径2个因子保留在冷箭竹发笋量模型中;将乔木郁闭度、株数和平均胸径3个因子保留在冷箭竹株数模型中;将乔木郁闭度、株数、平均高度和平均胸径4个因子保留在冷箭竹平均高度模型中;将乔木平均高度和平均胸径2个因子保留在冷箭竹平均基径模型中。
采用赤池信息准则(AIC)指标来比较不同模型间的模拟效果。AIC值越小,表明模拟效果越好。结果表明,所挑选的主要因子模拟效果均好于没有显著性影响的因子的模型拟合效果。使用不同的有显著性的因子作为固定效应,其混合模型的精度不同。在冷箭竹平均高度模型中,以乔木郁闭度作为固定效应的模型AIC值最小,拟合精度最高;在冷箭竹平均基径模型中,以乔木平均高度作为固定效应的模型AIC值最小,拟合精度最高;在冷箭竹发笋量模型中,以乔木平均胸径作为固定效应的模型AIC值最小,拟合精度最高;在冷箭竹株数模型中,以乔木株数作为固定效应的模型AIC值最小,拟合精度最高(表3)。
结果显示(表3),冷箭竹平均高度、平均基径、发笋量和株数模型的固定效应截距均达到极显著水平(P<0.01),说明乔木层结构特征对模型变化影响极显著。乔木株数和平均高度的固定效应系数对冷箭竹平均高度模型变化影响显著(P<0.05),乔木郁闭度和乔木平均胸径的固定效应系数对冷箭竹平均高度模型变化影响均极显著(P<0.01);乔木平均高度和平均胸径的固定效应系数对冷箭竹平均基径模型变化影响极显著(P<0.001);乔木平均高度的固定效应系数对冷箭竹发笋量模型变化影响显著(P<0.05),平均胸径对冷箭竹发笋量模型变化影响极显著(P<0.01);乔木株数的固定效应系数对冷箭竹发株数模型变化影响极显著(P<0.01)。
根据模型结果可知:随着乔木层郁闭度的增加,冷箭竹平均高度表现出减小趋势;随着乔木株数、平均高度和平均胸径的增加,冷箭竹平均高度均表现出增加趋势;随着乔木平均高度和乔木平均胸径增加,冷箭竹平均基径和发笋量均表现出增加趋势;随着乔木层郁闭度和株数的增加,冷箭竹株数均表现出减少趋势;随着乔木平均胸径的增加,冷箭竹株数表现出增加趋势(表3)。
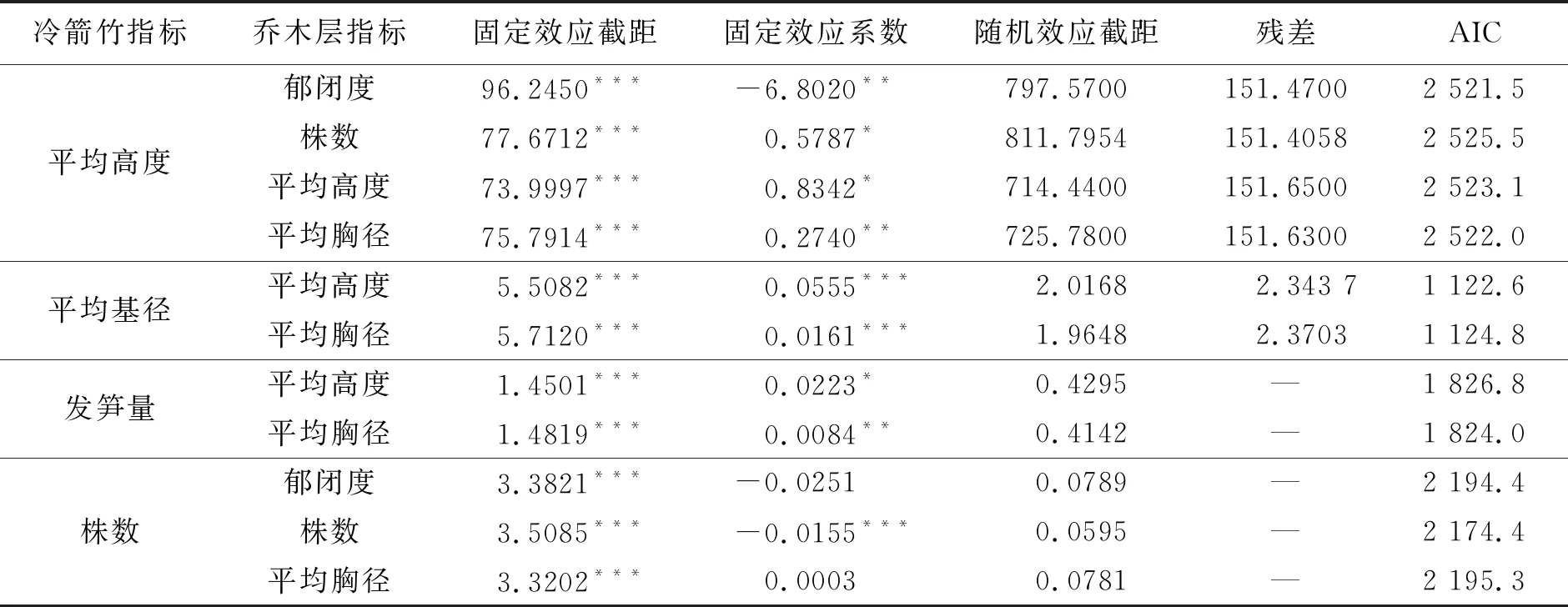
表3 冷箭竹生长指标模型拟合结果
3 讨 论
森林群落结构是在环境因子、群落物种间以及干扰因子等因素综合影响下形成的。本研究在大相岭大熊猫野化放归基地周边原生森林生态系统林下分布有冷箭竹的区域,利用样线法和样方法,开展森林群落乔木层与林下冷箭竹调查,并利用模型构建了大熊猫栖息地森林群落乔木层结构特征与林下冷箭竹生长指标的关系,这为大熊猫栖息地恢复提供了定量的数据支撑。
竹子的空间分布对于大熊猫栖息地的分布具有重要影响[17]。竹子密度、基径、高度和幼竹所占比例是衡量森林中竹子资源的重要指标[18],也是表征竹子生长状况的指标[19]。本研究结果表明,乔木平均高度和平均胸径对冷箭竹平均高度、平均基径和发笋量变化均影响显著,乔木株数对冷箭竹株数及平均高度影响显著。已有研究表明,竹类生长和更新受乔木层郁闭度影响[20],乔木层郁闭度的大小是影响缺苞箭竹(Fargesiadenudat)更新的最主要因子[21],魏辅文和冯祚建[22]认为成竹、发笋量、老笋比例、死亡率与上层乔木郁闭度呈负相关,宋利霞等[23]认为华西箭竹(Fargesianitida)分株种群的长势随林冠郁闭度的增大而渐差,Li等[24]对箭竹的研究表明箭竹幼苗密度随着海拔高度和乔木郁闭度的增加呈现先增加后减小的趋势。而本研究中,冷箭竹株数随着乔木株数的增加而减少,这可以解释为随着乔木株数增加,乔木层郁闭度也增加,过分荫蔽的环境影响根系的发育,使竹子幼苗的根系生长脆弱,根系竞争能力差,影响幼苗对水分和养分的吸收,造成幼苗死亡[21],冷箭竹株数减少。随着乔木层郁闭度的增加,冷箭竹平均高度减少,这与宋利霞[25]和张聪等[26]对华西箭竹的研究结果一致,但是与解蕊等[27]对缺苞箭竹的研究结果不一致。这说明郁闭度过高不利于冷箭竹生长,也体现了不同克隆植物对应环境异质性策略的差异。华西箭竹在大林窗(400 m2)环境具有较强的生长优势,这暗示了冷箭竹较华西箭竹的耐阴性差,适生于郁闭度更小的环境中[27]。本研究结果表明,冷箭竹平均高度与乔木株数、乔木平均高度和平均胸径均呈正相关关系,这与Kang等[19]的研究结果不一致。这可能与森林乔木层的树种组成有关。
大熊猫国家公园荥经片区是大相岭山系大熊猫种群的核心分布和集中分布区,也是连接大相岭山系和邛崃山山系大熊猫种群基因交流的关键区。因此,对该区域大熊猫栖息地进行修复是恢复大相岭山系大熊猫栖息地的重要基础,也是保护、维持大熊猫种群数量的有效途径。在大熊猫栖息地恢复过程中,能自然恢复的应尽量自然恢复,不能自然恢复的可以进行人工恢复。大熊猫、森林和竹子三者协同进化[28],所以可将森林和竹子之间的关系作为大熊猫栖息地人工恢复的参考标准。通过研究了解大相岭山系大熊猫栖息地森林群落乔木层结构特征与林下冷箭竹的生长指标之间的关系,可以科学合理地采取经营措施,以确保合理的乔木层结构特征,促进大熊猫取食竹更新生长。
猜你喜欢
临床检验杂志(电子版)(2020年1期)2020-04-03
福建林业(2020年5期)2020-03-18
四川林业科技(2019年5期)2019-07-07
热带林业(2019年4期)2019-03-05
新农民(2019年9期)2019-02-19
安徽农学通报(2019年1期)2019-02-14
湖北林业科技(2019年6期)2019-01-06
森林工程(2018年3期)2018-06-26
湖北林业科技(2017年2期)2017-05-25
学苑创造·B版(2015年12期)2016-06-23
