
6个广西种源荷木光合及保水力的比较
2023-02-05 02:40:46杨丽萍谭长强
亚热带农业研究 2023年3期
张 燕, 杨丽萍, 杨 康, 谭长强
(1.南宁市南湖公园,广西 南宁 530000; 2.广西壮族自治区林业科学研究院,广西 南宁 530002;3.南宁乡村振兴集团有限公司,广西 南宁 530000)
荷木(Schimasuperba)又名木荷、荷树,属山茶科(Theaceae)常绿乔木。荷木四季长青,树冠浓密、枝叶繁茂,叶片厚革质且含水量大、含油脂少、燃点较高,萌芽力强,是生态和生物防火的主要树种。因其树干端直、木材白色,细致易加工,是良好的家具和工艺加工用材,也是建筑、器材、国防等特种用材树种[1-2]。国内外学者对荷木生理生态[3-6]、防火林带及性能[7-8]、遗传特性及栽培技术[9-13]等方面做了大量研究工作。在种源研究上,王淼等[9]根据造纸和建筑家居等不同用途,从湖南、浙江、江西、广东、福建的24个市(县)木荷种源中选出了5个优良种源。吕欣欣等[14]基于SNP分子标记技术,从DNA分子水平上揭示了广东、福建和江西的15个木荷种质资源的遗传多样性和变异规律。姚甲宝等[15]开展了不同光照强度对福建、浙江、江西等3个荷木种源幼苗的影响。目前,尚未见广西荷木种源研究的相关报道。
光合特性能够反映植物对环境的适应能力和生长潜力,光合作用气体交换参数是植物在环境变化中最敏感的适应性特征之一。光响应曲线反映了植物随光强变化的光合能力,可了解植物光反应过程,是判定植物受环境变化影响程度及光合作用能力的重要依据[16]。树木通过多种途径实现对干旱的适应。在干旱条件下,叶片通过减少水分丧失来缓解干旱胁迫。叶片保水力通常用来表示植物组织的抗脱水能力。张明生等[17]、赵爽等[18]分别对甘薯、板栗的抗旱性研究表明,离体叶片失水率可作为评价其抗旱性的生理指标之一。
植物与地理环境之间存在密切的关系。北回归线横穿广西,造成广西南北气候差异较大;且广西东南近海,使当地气候条件受季风影响较为明显。本研究以位于北回归线以北的灌阳县、恭城县、苍梧县,以及以南的博白县、兴业县、钦州市等6个广西种源荷木为试验材料,通过比较不同荷木种群叶片的光合特征、气孔密度及保水力等指标,揭示自然条件下不同种源荷木光合特征及叶片生理的差异,旨在为其优良种源选择及利用提供依据。
1 材料与方法
1.1 种源及试验地概况
1.1.1 种源 2012年分别于广西灌阳(灌阳县)、恭城(恭城县)、苍梧(苍梧县)、博白(博白县)、兴业(兴业县)、钦州(钦州市)采集成熟的荷木种子,带回广西林科院珍贵树种苗圃(108°56′E,22°56′N)进行播种和育苗。6个种源地2002—2012年气象概况见表1,相关数据由广西气象局提供。

表1 2002—2012年6个种源地气象概况
1.1.2 试验地概况及试验林营建 试验地位于南宁市郊广西林科院科研试验林区(108°56′E,22°56′N)。海拔约85 m,坡向朝南,中上坡,年平均气温21.7 ℃,≥10 ℃积温为7 200 ℃,极端最低温-1.5 ℃,极端最高温39.4 ℃,全年无霜期高达360 d,年均降雨量1 300 mm,年均相对湿度80%左右。土壤为砖红壤,土层厚度>1 m,pH值为5~6,肥力中等。试验林地总面积约2 hm2,采用列区种植,每个种源种植0.25~0.34 hm2。造林地前身为松树林地,采取钩机带状整地。2012年3月种植,株行距为2 m×3 m,种植穴:50 cm×60 cm×50 cm。定植前每穴施1 kg基肥(含15%N+6%P2O5+9%K2O+15%有机质)和200 g钙镁磷肥。定植后采用相同的抚育管理措施,前3年每年追肥200 g和抚育除草2次。种植后未进行间伐,保存率均在90%以上。
1.2 测定指标与方法
于2022年11月,每个种源随机取3个20 m×20 m样方,调查样方内林木胸径、树高生长量。依据胸径和树高数据(表2),从每个样方选取3株生长旺盛且无病虫害的平均木进行相关试验。于晴天9点,剪取每株平均木向阳且生长旺盛的枝条的第4~第5片成熟叶片,测定保水力、叶绿素含量及气孔密度,3片1个重复,每株3个重复。

表2 广西不同种源荷木生长状况
1.2.1 叶片保水力 采用离体叶片自然失水法测定失水率。分别于采后0.5、1.0、2.0、4.0、6.0、10.0、14.0、18.0、24.0 h进行称重,最后将叶片在80 ℃下烘干至恒重。根据所得数据,计算出每次称重时的叶片含水量及累积失水量。失水率/%=(累积失水量/叶片含水量)×100。
1.2.2 叶绿素含量 采用丙酮—乙醇混合提取法[19]。称取0.2~0.3 g叶片,去除粗大的中心叶脉,剪成碎片放入离心管中,用10 mL体积比为1∶1的丙酮与无水乙醇提取液避光浸泡1 d,取上清液,用全波长酶标仪测定663、645 nm下的光密度,计算叶绿素含量。
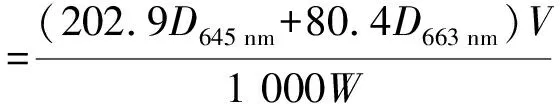
式中,D645 nm、D663 nm分别为相应波长下的光密度,V为提取液的体积,W为叶片鲜质量。
1.2.3 气孔密度 参考文献[20],采用指甲油涂抹法测定。用透明指甲油均匀涂抹叶片下表面待完全干燥后揭取下来,展平并制成气孔装片。在显微镜100倍的放大率下选择5~6个视野(总面积为3~5 mm2),观测每张叶片的气孔密度。
1.2.4 叶片气体交换参数及光响应特征 每株平均木取1枝向阳枝条,选取3片成熟叶测定。将测试叶片充分诱导活化后,利用Li-6400XT便携式光合分析仪测定气体交换参数,包括净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、胞间CO2浓度(intercellular CO2concentration,Ci)、蒸腾速率(transpiration rate,Tr)和水分利用效率(water use efficiency,RWUE)。
分别设定2 000、1 800、1 600、1 400、1 200、1 000、800、600、400、200、100、50、20、0 μmol·m-2·s-1共14个梯度光强测定光响应曲线。测定时控制叶片温度为(25±1) ℃,CO2为400 μmol·mol-1。采用直角双曲线修正模型[21]对净光合速率进行拟合,并计算表观量子利用效率(apparent quantum utilization efficiency,RAQY)、光补偿点(light compensation point,VLCP)、光饱和点(light saturation point,VLSP)、暗呼吸速率(dark breathing rate,Rd)、最大净光合速率(maximum net photosynthesis rate,Pmax)和RWUE等光合参数。
1.3 统计与分析
采用Excel对试验数据进行整理和统计,采用DPS 7.05软件进行方差分析和多重比较,采用Origin 2021做图。
2 结果与分析
2.1 6个广西种源荷木光合能力的比较
2.1.1 光响应特征 从表3可以看出,不同种源RAQY变化范围在0.069~0.089之间,平均值大小依次为:苍梧>恭城>灌阳≥博白≥兴业≥钦州,其中苍梧与钦州种源之间存在显著差异(P<0.05),其他种源间差异不显著。不同种源Pmax变化范围在11.14~14.51 μmol·m-2·s-1之间,大小依次为:灌阳>苍梧>恭城>钦州>博白>兴业,其中灌阳种源极显著高于其他种源,苍梧种源与恭城种源、博白种源与兴业种源之间无显著差异,苍梧及恭城种源与其他种源之间差异极显著。VLSP变化范围在1 072.87~1 635.45 μmol·m-2·s-1之间,大小依次为:灌阳>恭城>博白>苍梧>兴业>钦州,其中灌阳、恭城、博白种源之间差异不显著,极显著高于兴业、钦州种源;灌阳种源显著高于苍梧种源,恭城与博白、苍梧种源之间无显著差异。不同种源VLCP变化范围在11.54~24.63 μmol·m-2·s-1之间,大小依次为:恭城>灌阳>兴业>苍梧>博白>钦州,其中恭城种源极显著高于除灌阳外的其他种源,灌阳与兴业、苍梧种源两两之间无显著差异。Rd变化范围在0.76~1.84 μmol·m-2·s-1之间,大小依次为:恭城>苍梧>灌阳>兴业>博白>钦州,其中博白与兴业之间、苍梧与灌阳和兴业之间、恭城与苍梧种源之间差异不显著,恭城与苍梧以外的其他种源间差异显著。
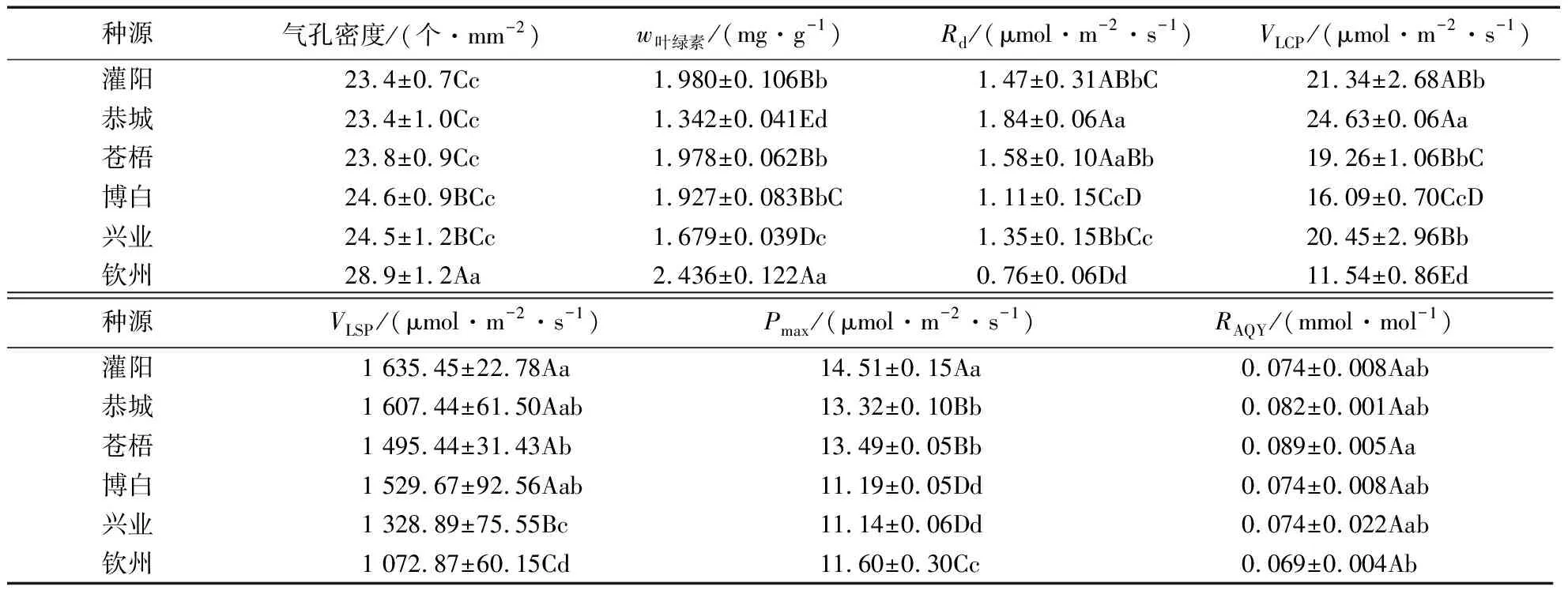
表3 6个广西种源荷木气孔及光合生理特征比较1)
2.1.2 叶绿素含量 不同种源叶绿素含量变化范围在1.342~2.436 mg·g-1之间,大小依次为:钦州>灌阳>苍梧>博白>兴业>恭城。其中,钦州种源极显著高于其他种源,恭城种源极显著低于其他种源,灌阳、苍梧、博白种源两两之间无显著差异。
2.1.3 气孔密度 不同种源荷木气孔密度变化范围在23.4~28.9个·mm-2之间,大小依次为:钦州>博白>兴业>苍梧>灌阳/恭城。其中,钦州种源极显著高于其他种源,博白、兴业、苍梧、灌阳、恭城种源两两之间无显著差异。
2.1.4 气孔交换特征 6个广西种源荷木叶片气孔交换特征见表4。从表4可见,Pn变化范围在10.6~15.0 μmol·m-2·s-1之间,大小依次为:灌阳>苍梧>博白>钦州>恭城>兴业,其中灌阳、苍梧显著高于其他种源,而其他种源间无显著差异。Gs变化范围在0.13~0.19 mol·m-2·s-1之间,大小依次为:灌阳>苍梧>钦州>恭城/博白>兴业,灌阳、苍梧、钦州种源显著高于恭城、博白、兴业种源,灌阳、苍梧、钦州种源两两之间差异显著。Ci变化范围在239.2~261.7 μL·L-1之间,大小依次为:钦州>灌阳>兴业>恭城>苍梧>博白,其中钦州种源显著高于恭城、苍梧、博白种源。Tr变化范围在2.34~3.04 mmol·m-2·s-1之间,大小依次为:灌阳>钦州>苍梧>博白>恭城>兴业,其中恭城、兴业种源显著低于其他种源。RWUE变化范围在3.91~4.94 mmol·mol-1,大小依次为:灌阳>苍梧>兴业>恭城>博白>钦州,其中灌阳种源显著高于恭城、博白、钦州种源。

表4 6个广西种源荷木叶片气孔交换特征比较1)
2.2 6个广西种源荷木叶片保水力的比较
6个广西种源荷木叶片失水率见图1。从图1可见,6个种源荷木离体叶片在不同时段失水率不同,但随着离体时间的延长,失水率都呈现不断提高的趋势,同时不同种源间失水率差异也随着离体时间的延长而逐渐变大。钦州种源失水速率明显要高于其他种源。24 h后,6个种源荷木失水率在44.82%~76.80%之间,其中灌阳种源失水率最低,其次为恭城、苍梧,最高的为钦州种源。综上所述,钦州种源保水力较差,而灌阳种源保水力较强。

图1 6个荷木种源叶片失水率的比较Figure 1 Water loss rate of leaves of 6 S.superba provenances from Guangxi
2.3 叶片指标与种源地气候条件的相关分析
不同种源荷木叶片指标与种源地气候条件的相关分析见表5。荷木叶片叶绿素含量与VLCP呈极显著负相关,与Rd呈显著负相关;气孔密度与VLSP、VLCP、Rd呈极显著负相关;VLSP与VLCP、Rd呈显著正相关;VLCP与Rd呈极显著正相关。气孔密度与年总降雨量呈显著正相关,与经度呈极显著负相关;年总降雨量与Rd呈极显著负相关,与RAQY、Pmax、VLSP呈显著负相关;Pmax与年平均气温呈显著负相关;RAQY、VLSP、VLCP、Rd与经度呈显著正相关;Pmax与纬度呈极显著正相关,VLSP、Rd与纬度显著正相关。胸径与Pn、Tr呈显著正相关。树高与胸径、Pn、Tr呈极显著正相关。Pn与Gs呈极显著正相关,RWUE与经度呈极显著正相关。

表5 不同种源荷木叶片指标与种源地气候条件相关分析1)
3 讨论
本研究通过分析各光合指标与种源产地生态环境因子之间的相关关系表明,气孔密度与年总降雨量呈显著正相关,与经度呈极显著负相关;年总降雨量与RAQY、Pmax、VLSP呈显著负相关,与Rd呈极显著负相关;Pmax与年平均气温呈显著负相关;RAQY、VLSP、VLCP、Rd与经度呈显著正相关;纬度与Pmax呈极显著正相关,与VLSP、Rd呈显著正相关。以上表明,经度、纬度、年总降雨量、年平均气温等是引起荷木生理及光合特性产生丰富遗传变异的主要环境因素[22-23]。
一般而言,来源于热带或南亚热带的种源,在高温强光天气下仍有较强的光合能力,而起源于温带或北亚热带的种源在温和的环境条件下,光合能力较强[24]。申文辉等[25]通过研究格木的光合特征表明,南部种源Rd要低于北部种源。本研究中,广西灌阳、恭城、苍梧等桂北荷木种源VLSP高,VLCP也高,阳性特征明显,有较好的适应强光能力,并且具有较高的RAQY和Pmax,利用光能能力较强,光合能力强,但利用弱光能力较弱。桂北种源(灌阳、恭城、苍梧)叶片在自然脱水的过程中,叶片含水量下降的速度小于钦州、博白种源,具有更低的失水率与较强的保水力。因此,在建造防火林带的时候应选择阳光较充足的阳坡,这将更有利于桂北种源的生长。而在较恶劣干旱的山地造林,也可适当选择靠北的荷木种源作为防火林带树种。博白种源具有较高的VLSP和较低的VLCP,说明其对光照的适应能力较强,利用光照的范围较广。钦州种源VLSP和VLCP都较低,保水力最低,说明其利用弱光能力较强,在生产上适合阴坡或半阴坡的地形造林,更有利于增强防火效果。
本研究中,Pmax与纬度呈极显著正相关,与赵晓焱等[26]研究结果相类似,与熊彩云等[27]则不同。赵晓焱等[26]研究表明,兴安落叶松(Larixgmelinii)光合能力随纬度升高有明显增加的趋势。熊彩云等[27]研究表明,随着纬度的增加,安徽、广东、江西等3个种源荷木的Pmax表现为南部种源高于北部种源。研究结果之间的差异可能由测量季节不同以及种植地理位置存在的气候条件不同所引起。熊彩云等[27]试验地位于其供试种源的中部,而本试验地位于供试种源地理位置的南部。赵勋[28]研究认为,温度是影响越南安息香(Styraxtonkinensis)光合能力的重要气候因子。曾伟等[29]、Friend et al[30]认为,亚热带常绿阔叶优势树种在不同季节具有不同的光合特性。本研究在12月份进行,此时南宁气温可能更适合北部种源荷木的生长,这可能是灌阳、恭城种源Pmax较高的原因。但在其他月份或季节,荷木的生理表现是否与12月份的表现相一致,尚待进一步验证。本研究中,荷木胸径仅与Pn、Tr呈显著正相关,树高仅与胸径、Pn、Tr呈极显著正相关,树高、胸径与其他指标之间无显著相关性。这说明荷木的生长受气孔交换特征的影响更多,而气孔交换特征受种植地气温、光合有效辐射和水汽压亏缺等环境因子的影响[31],说明种植地气候环境对荷木生长产生了较大影响。本研究中,Pn与Gs极显著正相关,与梁文斌等[32]对光皮树Pn与Gs的研究结果相一致,说明Gs可能是导致荷木种源Pn存在差异的重要因素。
在光合作用中,气孔是植物与周围环境气体交换的门户。刘亮等[33]研究表明,气孔密度可能与温度有关。张立荣等[34]对矮嵩草草甸4种植物研究表明,增温提高了矮嵩草(Kobresiahumilis)、垂穗披碱草(Elymusnutansnutans)、麻花艽(Gentianstraminea)的气孔密度,但降低了高山唐松草(Thalictrumalpinum)气孔密度。而Reddy et al[35]认为,温度升高可以提高叶片气孔密度。本研究表明,气孔密度与年均气温呈一定的正相关关系,6种种源气孔密度大小依次为:钦州>博白>兴业>苍梧>灌阳>恭城,呈现出桂南种源气孔密度比桂北种源大的趋势。在光合作用中,叶绿素起着能量接收和转换作用,叶绿素含量高的物种或种源的Pmax也高[36],但也有研究认为两者之间并没有显著的相关性[37]。本研究表明,叶绿素含量与Pmax之间无显著相关性(表2)。
4 结论
综上分析,桂北种源(灌阳、恭城)叶片含水量下降速度小于钦州、博白种源,具有更低的失水率和较强的保水力。博白种源对光照的适应能力较强,利用光照强度的范围较广,钦州种源利用弱光能力较强。总体上,桂南荷木种源比桂北种源气孔密度大。经度、纬度、年总降雨量、年平均气温等是引起荷木生理及光合特性产生丰富遗传变异的主要环境因素,但荷木生长主要受净光合速率的影响。
猜你喜欢
湘潮(上半月)(2021年4期)2021-07-20 08:05:28
果农之友(2018年6期)2018-10-27 11:02:40
铁道通信信号(2018年4期)2018-06-06 03:31:34
中国-东盟博览(旅游版)(2018年2期)2018-05-14 12:55:10
人大建设(2018年2期)2018-04-18 12:16:59
西江文艺(2017年15期)2017-09-10 15:41:18
西江文艺(2017年15期)2017-09-10 15:41:18
福建茶叶(2017年4期)2017-02-04 21:38:40
文史春秋(2016年1期)2016-12-01 05:41:53
农民科技培训(2016年4期)2016-04-20 13:32:48
