
爱帕琳肽13/血管紧张素受体AT1相关受体蛋白同源二聚体对人脐静脉内皮细胞行为的影响
2023-01-05 08:11:22王德秀李建设王太千肖长昊王学健
中国药理学通报 2023年1期
王德秀,李建设,宋 超,王太千,殷 月,肖长昊,王学健,鲁 洪,蔡 欣
(潍坊医学院1.基础医学院、2.临床医学院、3.生命科学与技术学院、4.药学院,山东 潍坊 261053)
近年来,随着生活水平的不断提高,动脉粥样硬化、高血压、冠心病、心衰等心血管疾病的发病率越来越高,严重威胁着人类的健康,探究这类疾病发病的病理生理机制以及寻找有效的药物成为当今的一项重大任务。血管重构(vascular remodeling,VR)是动脉血管内皮受损后,为适应内外环境变化而发生的结构和功能的改变,包括血管壁细胞的增生、肥大、凋亡、迁移、细胞外基质的产生及降解等细胞生物学变化[1]。研究发现VR既是心血管疾病恶化的重要病理基础,也是心血管疾病发生和发展的病因,因此,探究VR过程中血管内皮细胞的行为学已成为治疗心血管疾病的关键环节。
血管紧张素受体AT1相关受体蛋白(putative receptor protein related to AT1,APJ)是G蛋白偶联受体(G protein-coupled receptors,GPCRs)A类家族的成员之一,在心血管系统具有较高表达水平,爱帕琳肽(Apelin)是APJ的内源性配体,Apelin/APJ系统能促进细胞的增殖、迁移,还可促进血管生成,参与心血管功能,在心血管调节中具有降低血压、增强心脏收缩的作用,是心血管稳态中重要的调节因子[2]。近几年,APJ作为心血管相关疾病治疗靶点的潜力引起了越来越多的关注。事实上,临床前和最近的临床证据都表明,APJ是治疗肺动脉高压、动脉粥样硬化、心衰等心血管疾病的重要靶点[3-4]。本课题组前期研究发现,APJ除了能以单体的形式存在,还可以形成同源二聚体,在受体密度小于0.3颗粒/平方微米(particles/μm2)的情况下(静息状态),APJ同源二聚体占APJ总量的36%,Apelin-13能上调APJ表达,且同源二聚体的表达也随之升高[5],另外,APJ同源二聚体具有不同于单体的信号通路,能激活Gαi3和Gαq信号通路[6]。因此,本课题探讨APJ同源二聚体对人脐静脉内皮细胞( human umbilical vein endothelial cells,HUVECs)增殖、迁移和管腔形成能力的影响,该研究将有助于为临床的血管修复重建以及心血管疾病的治疗提供新靶点和思路。
1 材料与方法
1.1 实验细胞HUVECs和中国仓鼠卵巢细胞(Chinese hamster ovary cell, CHO)(中国科学院上海生命科学研究院细胞库)。
1.2 药品与试剂Apelin -13(美国Phoenix Pharmaceuticals Inc,批号:427374),DMEM基础培养基(美国Cytiva公司,批号:SH30243.01),胰酶EDTA消化液和青链霉素溶液(100×)(北京索莱宝公司,批号:T1300、P1400),Biocoat 356234 Matrigel基质胶(美国康宁公司,批号:356234),兔抗APJ抗体和小鼠抗β- actin抗体(美国Proteintech公司,批号:20341-1-AP、66009-1-Ig),Laser BioLab MALDI Calibration kit和MonoTip C18(日本Shimadzu公司,批号:TO-724R00、5010-21002 )。
1.3 仪器Herocell 180型细胞培养箱(美国Thermo公司),multiskan go酶标仪和FluorChem Q化学发光凝胶成像系统 (美国Protein Simple公司),AXIMA Assurance基质辅助激光解吸飞行时间质谱仪(日本Shimadzu公司),xCELLigence实时细胞分析仪(美国ACEA Biosciences公司)
1.4 方法
1.4.1细胞培养 HUVECs和CHO细胞置于10% FBS的DMEM培养基,于37 ℃,5% CO2的培养箱进行常规培养。
1.4.2Western blot实验 弃掉培养基,预冷PBS洗涤细胞,加入含有PMSF的RIPA裂解液,枪头吹打,收集细胞到EP管中,4 ℃离心,取上清,一部分样品用于BCA法测定蛋白浓度,剩余样品加入4×上样缓冲液,煮沸10 min,分装进行Western blot。样品经电泳和转膜后,用脱脂奶粉封闭1 h,兔抗APJ(1 ∶1 000)于4 ℃冰箱内孵育过夜后,洗膜,HRP 标记的山羊抗兔多克隆抗体( 1 ∶2 000)于室温孵育1 h,洗膜后加入ECL化学发光工作液,采用超灵敏化学发光成像仪成像,随后用超纯水和再生液处理膜,进行内参β-actin的表达,抗体分别是鼠抗β-actin(1 ∶1 000)和山羊抗鼠单克隆抗体( 1 ∶10 000)。根据得到的条带对实验结果进行定量分析。
1.4.3基质辅助激光解吸飞行时间质谱(Matrix-Assisted Laser Desorption/Ionization Time of Fligh Mass Spectrometry, MALDI-TOF MS)分析 HUVECs长到90%左右将蛋白提出,分别加入5 μL兔抗APJ抗体和50 μL protein A/G beads 4 ℃孵育过夜,3 000×g离心去除上清,TBS 溶液清洗3次,0.1 mol·L-1glycine-HCl 洗脱抗原后,3 000×g离心,取上清为待分析蛋白样品。所得待分析蛋白样品经 MonoTip C18脱盐后上样。样品检测结果用 Laser BioLab MALDI Calibration kit 进行校正。
1.4.4Apelin-13引起50%最大效应的浓度 (EC50)的检测 在E-Plate16的孔中加入100 μL培养基后,进行基线检测(cell index<0.063),随后向孔中再加入100 μL HUVECs悬液,10 000个细胞/孔,在培养箱中放置 30 min 后,放在实时细胞分析仪(Real-Time Cell Analyzers,RTCA)上,24 h后,分别用0、0.01、0.05、0.1、0.5、1、5和 10 μmol·L-1的 Apelin-13 处理HUVECs,结果用 RTCA Software 2.0进行分析。
1.4.5细胞分组 课题组前期研究发现,APJ同源二聚体在Apelin-13的刺激下表达增多,且APJ 的跨膜区1(transmembrane domain 1,TM1)能阻断APJ同源二聚体的形成[5],基于此,本研究通过添加Apelin-13作为APJ同源二聚体组,添加 TM1作为APJ单体组。将HUVECs分为空白对照组、APJ同源二聚体组和APJ单体组,分别用 PBS、Apelin-13(100 nmol·L-1)+PBS和Apelin-13(100 nmol·L-1)+TM1(4 μmol·L-1)处理,APJ单体组中TM1在37 ℃,孵育1 h后再加入Apelin-13。
1.4.6CCK-8实验 HUVECs消化后按1×105个/孔接种于96孔板中。待细胞长至贴壁后,加入10 μL/孔的CCK-8溶液,根据分组条件对各组细胞进行不同刺激,37 ℃,1 h,用酶标仪(450 nm)测定各孔的吸光度(OD) 。
1.4.7细胞划痕实验 细胞培养于6孔板,待长到80%左右,去除培养基,用10 μL枪头在孔中间划一道竖线,PBS洗两遍去除划下来的细胞,然后根据分组条件对各组细胞进行不同刺激,并放入培养箱内继续培养,分别于2、4、8、16和24 h后拍照观察细胞迁移的状态,进行统计学分析。
1.4.8细胞成管实验 实验前24 h将高压好的100 μL枪头放入 -20 ℃冰箱预冷,基质胶放入4 ℃ 冰箱液化备用。用预冷枪头向置于冰上的96孔板中加入80 μL/孔的基质胶,随后放入培养箱中固化基质胶1 h。各组细胞用胰酶进行消化,按照2×104个细胞/孔的密度种在基质胶上,并将96孔板放于37 ℃、5 % CO2培养箱中继续孵育4 h后,显微镜下观察HUVECs的成管情况并拍照计算形成管腔数,做统计学分析。
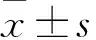
2 结果
2.1 APJ在HUVECs中的表达将HUVECs和CHO细胞提取蛋白后进行Western blot实验,根据APJ 的蛋白分子量发现(Fig 1),HUVECs中蛋白在43 kD处存在表达,与理论值相符,CHO无表达,表明HUVECs中存在APJ的表达。

Fig 1 Expression of APJ in HUVECs and CHO
2.2 APJ同源二聚体在HUVECs中的表达根据MALDI-TOF MS质谱扫描结果进行数据分析,如Fig 2所示,APJ不仅能以单体的形式存在,还能以同源二聚体形式在HUVECs中表达,且同源二聚体表达量是单体的0.22倍(P<0.01)。

Fig 2 Expression of APJ homodimer in
2.3 不同浓度的 Apelin-13 对HUVECs活力的影响为了探究APJ同源二聚体对HUVECs行为的影响,我们首先用RTCA检测引起 50%最大效应的浓度(EC50),如Fig 3A所示,分别用不同浓度的Apelin-13(0、0.01、0.05、0.1、0.5、1、5和10 μmol·L-1)处理HUVECs ,并用 RTCA Software 2.0 对结果进行分析,如Fig 3B所示,EC50为 2.26×10-8mol·L-1,且引起最大效应的浓度为1×10-6mol·L-1。

Fig 3 Effects of different concentrations of Apelin-13
2.4 APJ同源二聚体促进HUVECs增殖本实验利用CCK-8法检测对照组、APJ同源二聚体组和单体组的HUVECs增殖情况,如Fig 4所示,与PBS组相比,Apelin-13+PBS和Apelin-13+TM1组的HUVECs的增殖显著增加,16 h的增殖最快,同时,Apelin-13+PBS组HUVECs的增殖效果明显快于Apelin-13+TM1组,差异有统计学意义(P<0.05),表明APJ同源二聚体的促增殖能力强于APJ单体。

Fig 4 Effect of APJ homodimer on proliferation
2.5 APJ同源二聚体促进HUVECs迁移HUVECs划痕实验结果如Fig 5A所示,随着培养时间的不断延长,各组细胞向划痕裸区迁移,划痕距离逐渐减小。数据统计如Fig 5B所示,与PBS组相比,各时间点的Apelin-13+PBS组和Apelin-13+TM1组的迁移速度明显加快,其中Apelin-13+PBS组的迁移速度明显快于Apelin-13+TM1组,且16 h的迁移速度最快(P<0.01),表明APJ同源二聚体的促迁移能力高于APJ单体。

Fig 5 Effect of APJ homodimer on migration
2.6 APJ同源二聚体促进HUVECs成管镜下观察 HUVECs 成管情况,如Fig 6A所示,发现各组细胞在基质胶上培养4 h后均出现成管,激动剂处理时间越长,小管的密度越多,总管腔面积也越大。数据统计如Fig 6B所示,与PBS组相比,各时间点的Apelin-13+PBS组和Apelin-13+TM1组的细胞成管数目明显增多,16 h细胞成管数目最多,且Apelin-13+PBS组的成管数目明显多于Apelin-13+TM1组(P<0.05),表明APJ同源二聚体的促成管能力高于APJ单体。

Fig 6 Effect of APJ homodimer on tube formation
3 讨论
生理状态下,成体的血管内皮细胞更新率很低,但是在病理状态(如动脉粥样硬化、恶性肿瘤、糖尿病等)下可以发生血管新生,即血管内皮细胞脱离原来的静息状态,出现增殖、迁移并以出芽方式形成管状结构,管状结构的不断延伸逐渐与周围血管连接形成新生血管[7-8]。血管新生在肿瘤生长、血管生成、血管重构和炎症反应等许多生理病理过程中发挥着关键作用[9-10]。在视网膜血管系统中,内皮细胞迁移引起血管收缩,收缩的血管通过内皮细胞迁移的方式进行血管重塑,形成完整的血管网络;高糖诱导细胞焦亡分泌的白介素1β通过促进大鼠原代主动脉平滑肌细胞增殖迁移,加速动脉损伤后内膜的新生[11]。人参皂苷通过促进血管生成改善糖尿病大鼠的皮肤损伤[12],卵泡抑素样蛋白1通过增强肺动脉内皮细胞的增殖、黏附以及血管的形成进行肺血管重塑,从而预防和治疗肺动脉高压[13]。可见,内皮细胞正常的增殖、迁移和成管在VR和各类心血管疾病中发挥着非常重要的作用。
Apelin是第一个被发现的APJ的内源性配体,在心血管系统中发挥着重要作用,大量研究表明, Apelin/APJ系统能通过ERK/cyclinD1途径促进血管平滑肌细胞的增殖[14],在自发性高血压大鼠中通过激活自噬促进血管平滑肌细胞增殖[15]。Apelin/APJ系统还可促进H9c2心肌细胞和人主动脉血管平滑肌细胞的增殖[16-17]。另外,Apelin-13能够促进猴脉络膜/视网膜内皮细胞增殖、迁移和管腔的形成,加速视网膜新生血管的形成。本课题组前期通过生物物理和生物化学的方法(免疫共沉淀、荧光共振能量转移与生物发光共振能量转移等)已在体外检测到APJ同源二聚体的存在[5],本实验通过Western blot和MALDI-TOF MS在原生细胞中再次证实,APJ和APJ同源二聚体均能表达于HUVECs,且Apelin-13的EC50为 2.26×10-8mol·L-1,引起最大效应的浓度为1.0×10-6mol·L-1。随后我们利用CCK-8实验测定在1.0×10-7mol·L-1Apelin-13的刺激下,APJ同源二聚体对HUVECs增殖的影响,实验结果发现,Apelin-13+PBS组(APJ同源二聚体组)和Apelin-13+TM1组(APJ单体组)的HUVECs的增殖能力较PBS组显著增强,且Apelin-13+PBS组的增殖效果明显好于Apelin-13+TM1组,同时,发现Apelin-13处理16 h的增殖最快,说明APJ同源二聚体的促增殖能力强于APJ单体。利用细胞划痕法检测APJ同源二聚体对HUVECs迁移的影响,结果显示Apelin-13+PBS组细胞向划痕裸区迁移速度明显快于Apelin-13+TM1组,且16 h的迁移速度最快,表明APJ同源二聚体的促迁移能力高于APJ单体。通过人工基底膜(Matrigel)包被检测APJ同源二聚体对HUVECs成管能力的影响,结果发现细胞在基质胶中孵育后均能出现成管,随着激动剂刺激时间延长,小管的密度逐渐增多,总管腔面积也逐渐增大。各时间点的Apelin-13+PBS组和Apelin-13+TM1组的细胞成管数目明显多于PBS组,激动剂刺激16 h细胞成管数目最多,且Apelin-13+PBS组的成管数目明显多于Apelin-13+TM1组,上述结果表明APJ同源二聚体的促增殖、迁移和成管能力均高于APJ单体。
综上,本实验研究发现,APJ同源二聚体对HUVECs增殖、迁移和血管形成都有促进作用,优于单体,且激动剂处理16 h后的作用最为显著。我们将以此为基础,进一步探究APJ同源二聚体对VR的调节机制,为 APJ 参与的重要生理病理过程(动脉粥样硬化、高血压等)提供细胞内分子机制的实验依据,有助于更精确地了解相关疾病的发生并提供更有效的、特异的治疗靶点。
猜你喜欢
——紫 苏
河南农业(2024年1期)2024-01-19 01:56:54
华人时刊(2023年1期)2023-03-14 06:43:36
汉字汉语研究(2021年2期)2021-08-30 08:58:46
中国军转民(2017年7期)2017-12-19 13:30:00
河北书画研究(2016年3期)2016-04-28 08:55:35
中国卫生标准管理(2015年16期)2016-01-20 09:26:29
大连工业大学学报(2015年4期)2015-12-11 04:06:50
现代检验医学杂志(2015年2期)2015-02-06 02:01:09
现代检验医学杂志(2015年1期)2015-02-06 01:59:20
中国卫生(2014年10期)2014-11-12 13:10:24
