
淹水胁迫对星花玉兰及其相关品种叶片光合特性的影响
2022-12-16 11:35陈伯祥杜习武胡永红
植物资源与环境学报 2022年6期
陈伯祥, 杜习武,, 秦 俊,3, 曾 丽, 胡永红,3, 叶 康,3,①
(1. 上海辰山植物园, 上海 201602; 2. 上海交通大学农业与生物学院, 上海 200240;3. 上海城市树木生态应用工程技术研究中心, 上海 200020)
水分是影响植物生长发育的重要非生物因子之一。生境中水分过多会抑制植物生长,并对植物多项生理生化指标产生较大影响[1-3]。梅雨季节,长三角地区常发生水涝灾害。随着城镇化的快速推进,城市不透水地面比例不断增大,地表径流增加,易造成城市内涝[4]。
玉兰属(YulaniaSpach)植物是中国重要的园林绿化树种,栽培历史悠久[5,6]。星花玉兰〔Y.stellata(Maxim.) N. H. Xia〕为玉兰属低矮灌木,具有树冠饱满、分枝密集、花呈星芒状且花量大等特点,观赏价值和园林应用价值均较高[7]。星花玉兰及其相关品种的叶片在地势低洼、排水不畅的环境中常常萎蔫变黄,甚至干枯脱落,导致其在相应环境园林中的推广应用受限。
叶片光合作用水平对植物生长、产量及抗逆性等具有重要影响,可作为判定植物生长和抗逆性能力的重要生理指标[8]。鉴于此,本文对星花玉兰及其相关品种‘菊花’(‘Chrysanthemiflora’)和‘贝蒂’(‘Betty’)3年生嫁接苗叶片光合特性指标在淹水胁迫14 d内的变化进行了比较,以期为星花玉兰及其相关品种在低洼地的种植提供参考依据。
1 材料和方法
1.1 材料
所有研究对象均为以望春玉兰〔Y.biondii(Pamp.) D. L. Fu〕为砧木的3年生嫁接苗,接穗分别是星花玉兰及其相关品种‘菊花’和‘贝蒂’。其中,星花玉兰平均株高166.8 cm、平均地径22.4 mm;‘菊花’为从星花玉兰实生苗中选育的品种,平均株高117.8 cm、平均地径21.1 mm;‘贝蒂’为星花玉兰与紫玉兰〔Y.liliflora(Desr.) D. L. Fu〕的杂交品种,平均株高131.5 cm、平均地径23.1 mm。供试植株均健康、无病虫害,且种植在上口径53.5 cm、底径35.0 cm、高36.0 cm的塑料花盆中,栽培基质为V(园土)∶V(草炭)∶V(介质土)=5∶2∶3的混合基质。
1.2 方法
1.2.1 实验设计 采用双套盆法[9],于2020年5月27日在上海辰山植物园试验地开始淹水实验。处理组淹水深度始终保持在土面上方3 cm;对照组正常浇水,即表层土壤不干不浇水,浇水则浇透。每种植物处理组和对照组各20株样株。
1.2.2 光合特性指标测定 在淹水处理1、3、7和14 d分别测定叶片光合特性指标。在检测日9:00—11:00,使用LI-6400光合仪(美国LI-COR公司)测定植株中部侧枝上第3枚成熟叶片的净光合速率、气孔导度、胞间CO2浓度和蒸腾速率。采用标准LI-COR叶室,在红蓝光源(6400-02 LED光源)下进行检测,测定时光照强度为1 400 μmol·m-2·s-1,空气流速为500 μmol·s-1。每种植物处理组和对照组各选5株样株检测,每枚叶片重复测定2次。
1.3 数据分析
采用EXCEL 2010软件对相关数据进行统计分析;采用SPSS 22.0软件进行差异显著性分析,其中不同淹水时间的差异分析采用Duncan’s新复极差法,对照组和淹水组间的差异分析采用独立样本t检验。
2 结果和分析
对淹水14 d内星花玉兰及其相关品种叶片的光合特性指标变化进行比较,结果(表1)表明:随着淹水时间的延长,对照组(正常浇水)星花玉兰、‘菊花’和‘贝蒂’的叶片净光合速率变化相对较小,而淹水组3种植物的叶片净光合速率变化相对较大且逐渐降低。与对照组相比,3种植物的叶片净光合速率均显著(P<0.05)下降,且降幅随淹水时间延长逐渐增大;在淹水1、3、7和14 d,星花玉兰的叶片净光合速率分别下降了29.0%、48.1%、92.5%和100.9%,‘菊花’的叶片净光合速率分别下降了26.8%、51.4%、94.0%和99.1%,‘贝蒂’叶片净光合速率分别下降了17.3%、31.6%、88.6%和94.6%。
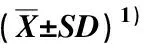
表1 淹水胁迫下星花玉兰及其相关品种叶片光合特性指标的变化
随着淹水时间的延长,对照组3种植物的叶片气孔导度变化相对较小,而淹水组3种植物的叶片气孔导度变化相对较大且逐渐下降。与对照组相比,3种植物的叶片气孔导度均显著下降,且降幅随淹水时间延长逐渐增大;在淹水1、3、7和14 d,星花玉兰的叶片气孔导度分别下降了19.2%、50.6%、71.0%和84.7%,‘菊花’的叶片气孔导度分别下降了27.4%、57.2%、81.4%和95.6%;‘贝蒂’的叶片气孔导度分别下降了20.7%、34.0%、76.4%和90.5%。
随着淹水时间的延长,对照组3种植物的叶片胞间CO2浓度变化相对较小,而淹水组3种植物的叶片胞间CO2浓度变化相对较大且逐渐升高。与对照组相比,3种植物的叶片胞间CO2浓度均升高,且升幅随淹水时间延长逐渐增大;在淹水1、3、7和14 d,星花玉兰的叶片胞间CO2浓度分别升高了9.6%、16.4%、85.8%和135.0%,‘菊花’的叶片胞间CO2浓度分别升高了2.5%、4.1%、59.7%和75.4%,‘贝蒂’的叶片胞间CO2浓度分别升高了2.1%、10.1%、42.0%和74.1%。
随着淹水时间的延长,对照组3种植物的叶片蒸腾速率变化相对较小,而淹水组3种植物的叶片蒸腾速率变化相对较大且逐渐下降。与对照组相比,3种植物的叶片蒸腾速率均下降,且降幅随淹水时间延长逐渐增大;在淹水1、3、7和14 d,星花玉兰的叶片蒸腾速率分别下降了5.3%、23.4%、61.4%和80.6%,‘菊花’的叶片蒸腾速率分别下降了7.5%、30.2%、65.4%和86.2%,‘贝蒂’的叶片蒸腾速率分别下降了8.0%、18.0%、67.8%和83.0%。
3 讨论和结论
淹水环境对植物生长造成胁迫的主要原因是生境出现低氧胁迫[10],植物体在低氧环境下的水分输送效率下降,致使植株处于生理性干旱状态[11]。星花玉兰、‘菊花’和‘贝蒂’在受到淹水胁迫后叶片的净光合速率均较对照组(正常浇水)显著下降,并随着淹水时间延长逐渐下降。在淹水7 d,3种植物淹水组的叶片净光合速率均较对照组降低了约90%,说明淹水7 d时供试3种植物的叶片光合系统严重受损。比较而言,在淹水1、3、7和14 d,‘贝蒂’的叶片净光合速率较对照的降幅最低,表明‘贝蒂’可能具有更强的耐水淹能力,3种植物的具体耐水淹能力有待进一步研究。在淹水胁迫下,星花玉兰、‘菊花’和‘贝蒂’叶片的气孔导度和蒸腾速率均低于对照组,并随着淹水时间延长逐渐下降,表明3种植物叶片上的气孔随淹水时间延长逐渐关闭,并伴随着蒸腾速率逐渐降低,以此提高叶片的保水能力。另外,气孔导度下降导致细胞间CO2的扩散阻力增大[12],净光合速率下降导致光合作用消耗的CO2量减少,最终导致胞间CO2大量堆积。综上所述,淹水胁迫对供试3种植物叶片的光合特性均有一定的影响,比较而言,‘贝蒂’受到的影响最小。
猜你喜欢
东北林业大学学报(2022年4期)2022-05-16
农业工程学报(2022年3期)2022-04-16
湖北农业科学(2021年13期)2021-08-04
新疆农业科技(2021年2期)2021-07-03
生态学报(2021年6期)2021-04-25
农业工程学报(2021年3期)2021-04-15
节水灌溉(2020年9期)2020-09-16
中学生物学(2020年12期)2020-03-30
作文与考试·小学高年级版(2019年8期)2019-04-29
微型小说选刊(2014年25期)2014-11-17
