
茂兰喀斯特区68种不同功能(类)群植物比叶面积研究
2022-12-13 12:13赵文君崔迎春周汀侯贻菊周华吴鹏
贵州林业科技 2022年4期
赵文君,崔迎春,周汀,侯贻菊,周华,吴鹏
(1.贵州省林业科学研究院,贵州 贵阳 550005;2.贵州荔波喀斯特森林生态系统国家定位观测研究站,贵州 荔波 5584003;3.贵阳市林长制工作服务中心,贵州 贵阳 550002)
植物功能性状是连接植物与环境的桥梁,是植物在长期适应环境过程中形成的能最大程度利用各种外部资源的形态、生理和物候等属性[1]。叶片是植物获取资源的重要器官,对环境变化敏感且可塑性大[2],比叶面积被认为是最适合研究的植物功能性状,它能代表土壤的肥力,是叶干物质含量与叶厚度综合作用的结果[3]。比叶面积(Specific leaf area,SLA)指单位干重的鲜叶表面积,是物种层面的特征参数,是植物资源摄取策略的关键叶片性状之一[2]。比叶面积与植物相对生长速率、光合速率、叶片氮含量有关,可用来反映植物的碳获取策略,对植物的相对生长速率具有重要影响,是生理过程权衡的最佳指示者[4-5],它因遗传特性和环境条件不同而异,即使在相同生境内的同一物种,也会因植物个体生长阶段、生存策略和资源利用效率的不同而表现出差异性[6]。
喀斯特生态系统是一种特殊的、脆弱的生态系统类型,其上的森林植被无论在群落外貌、结构以及区系组成,还是在演替更新以及对环境的影响等方面都与常态地貌上的明显不同[7-8]。近年来,对喀斯特地区植物叶片功能性状的研究进行了初步探索,但与其他常态地貌区相比,研究资料仍相对较少。如习新强等[9]研究了黔中喀斯特山区植物的比叶面积和叶面积在灌丛、落叶阔叶林、落叶常绿混交林3个演替阶段的变化规律,盘远方等[4]研究了岩溶山灌丛植物叶功能性状和土壤因子对坡向的变化规律。
本研究以茂兰喀斯特区68种不同功能(类)群植物为研究对象,通过不同生长阶段植物叶面积、叶干重及比叶面积的测量及计算,探讨不同生活型植物、不同系统发育类型植物及不同科植物比叶面积的变化特征,以期深入理解植物在长期进化过程中的适应机理,为喀斯特森林生态恢复和科学管理提供一定的参考价值。
1 材料与方法
1.1 研究区概况
研究地位于贵州茂兰国家级自然保护区(107°52′12″~108°5′42″E,25°9′27″~25°20′40″N),属于世界上同纬度地区留存下来仅有的、原生性强、相对稳定的喀斯特森林生态系统,是喀斯特区原生性森林常绿落叶阔叶混交林分布面积最大的地区[10],其自然状况参见相关文献[10-12]。该区属于典型的喀斯特地貌(喀斯特峰林及峰丛洼地),土壤以石灰土为主,土层浅薄(约20 cm)且不连片,多存在于岩石裂隙或出露岩石形成的沟地中,富钙和富盐基化、有机质含量高,土壤的理化性质、剖面形态等区别于地带性土壤[10,13]。
1.2 研究方法
1.2.1 样地设置和样地调查
2015年5月,通过对贵州茂兰国家级自然保护区踏查,共设置了12个固定样地,分别为顶极常绿落叶阔叶混交林群落、乔林群落(面积均为30 m ×30 m)、灌木灌丛群落(面积为20 m × 20 m)和草本群落(面积为10 m × 10 m),每种群落各设3个样地。各样地坡度、坡向、经纬度、海拔等用手持GPS和罗盘仪测量,郁闭度、灌木草本盖度、岩石裸露度和坡位等用目测法估测。采用每木检尺法对乔木层调查(胸径≥5 cm),采用样方法对灌草层调查,记录物种名称、郁闭度(或盖度)、株数、高度、胸径(或地径)和冠幅等因子,固定样地基本情况详见表1。
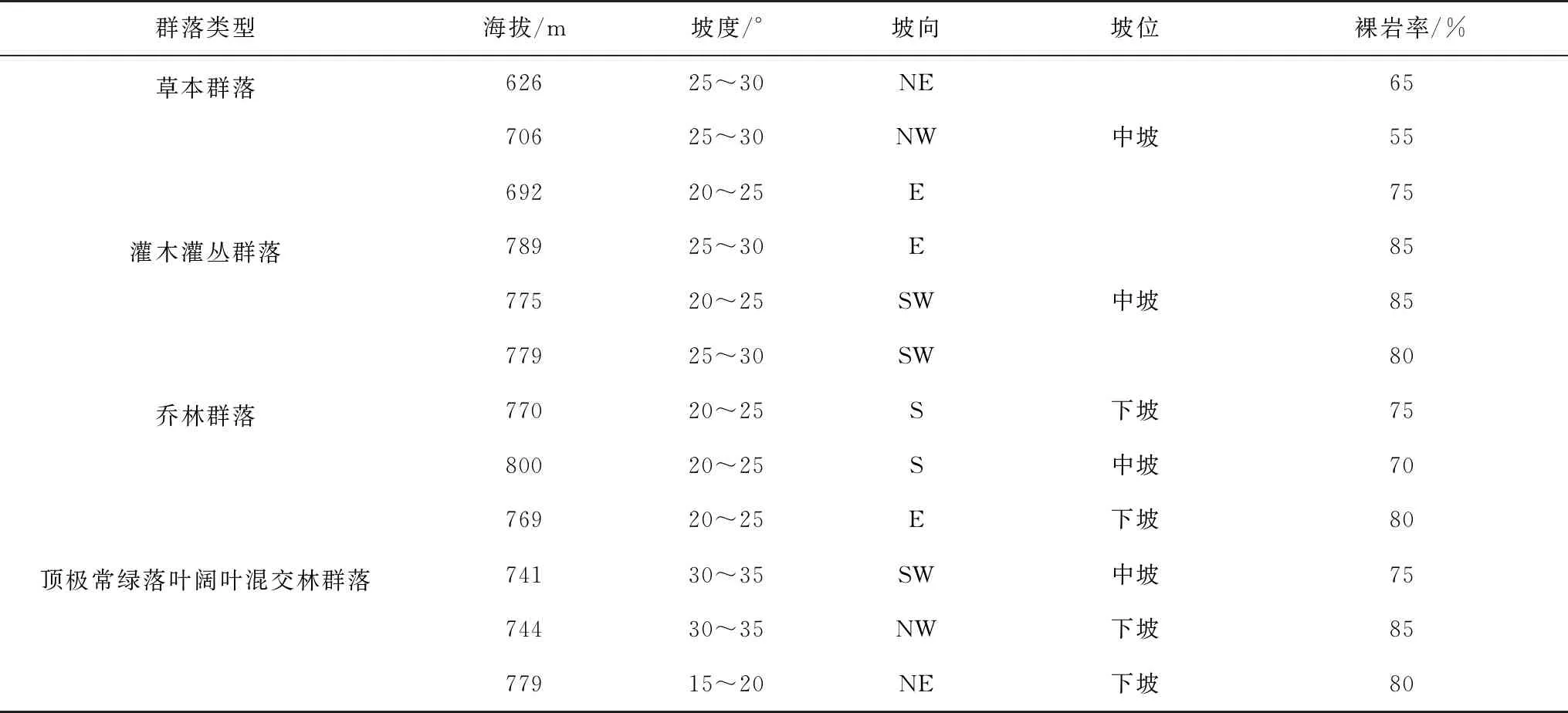
表1 固定样地基本情况
1.2.2 样品采集、处理及测定
采集方法:对每个样地内的乔木层、灌木层和草本层等不同群落层次的优势种[14]分别于生长末期(2015年10—11月)和生长盛期(2016年5—6月)进行取样。乔木层树种按优势树种分不同径阶(DBH分别以7.5 cm、22.5 cm为界)进行取样,1 株树为 1个重复,用高枝剪剪取冠层不同方位的枝条,采摘其上完整、全伸展、无病害、去叶柄的叶片,充分混合后,采用四分法(样品量≥30 片),密封后置于便携式冷藏箱中备用。灌木层采集方法同乔木层。同种植物以2~3株的混合样为1个样品;高大草本或蕨类植物以5~7株(丛)混合样为1个样品;低矮草本或蕨类植物以≥300 g新鲜样为1个样品;叶片数量不等,直接用剪刀剪取新鲜、完整并充分展开的叶片。如样地内对应径阶样株数量不足,可在样地外适量补充[14]。
样品处理及测定:在植物生长末期和生长盛期,选取采集的每种优势植物叶片5~10片,用便携式叶面积仪(LI-3100,America)测量叶面积大小,洗净烘干至恒重,用电子秤(精度为万分之一)称其质量。
植物比叶面积(Specific leaf area,SLA)的计算公式如下:
(1)
1.3 数据处理
所有数据采用Excel 2010处理,Origin2021作图,SPSS 16.0进行统计分析。用K-S(Kolmogorov-Smirnov test)检验数据正态分布性,采用one-way ANOVA分析不同类型SLA之间的差异显著性,应首先将其进行对数转换,以满足ANOVA假设和正态分布。
变异系数CV(Coefficient of variation)是描述变量空间变异程度的主要指标,计算公式如下:
(2)
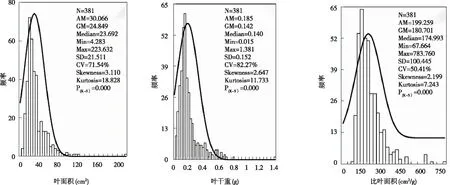
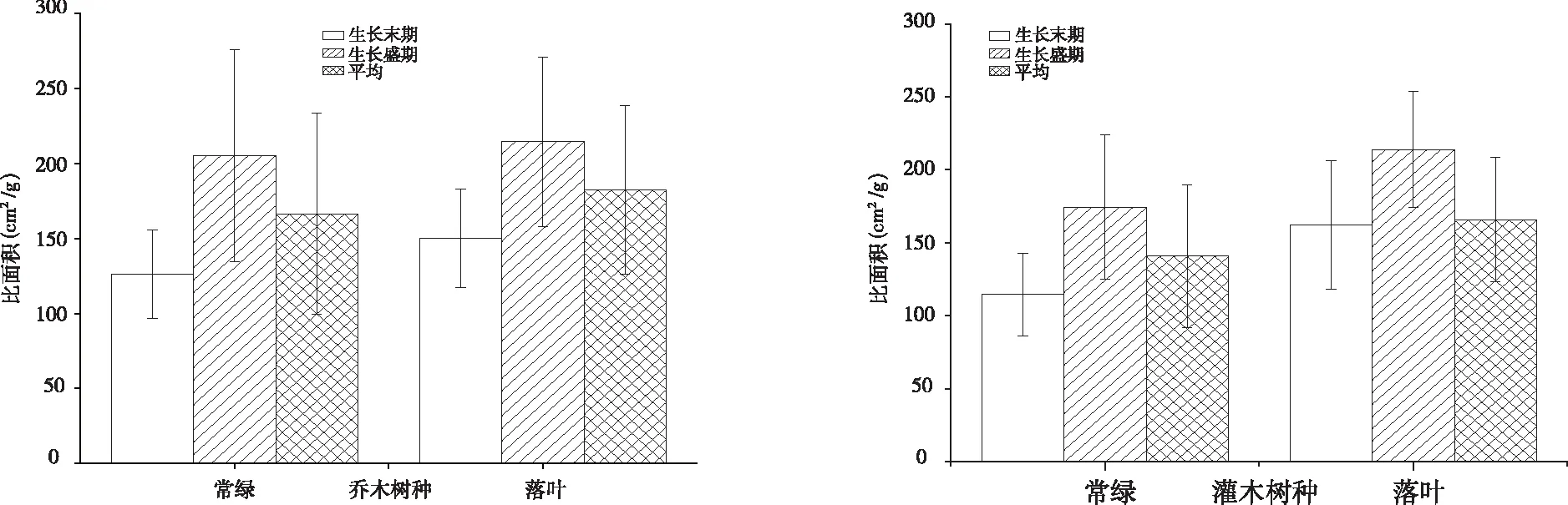
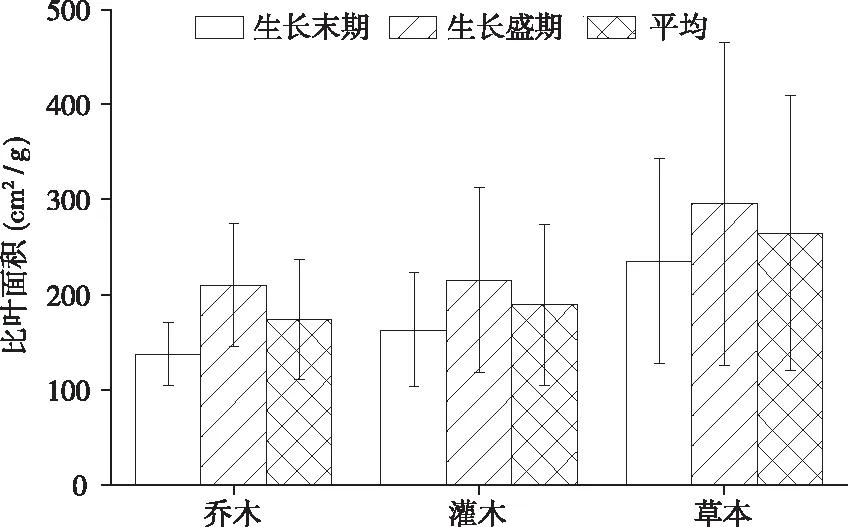
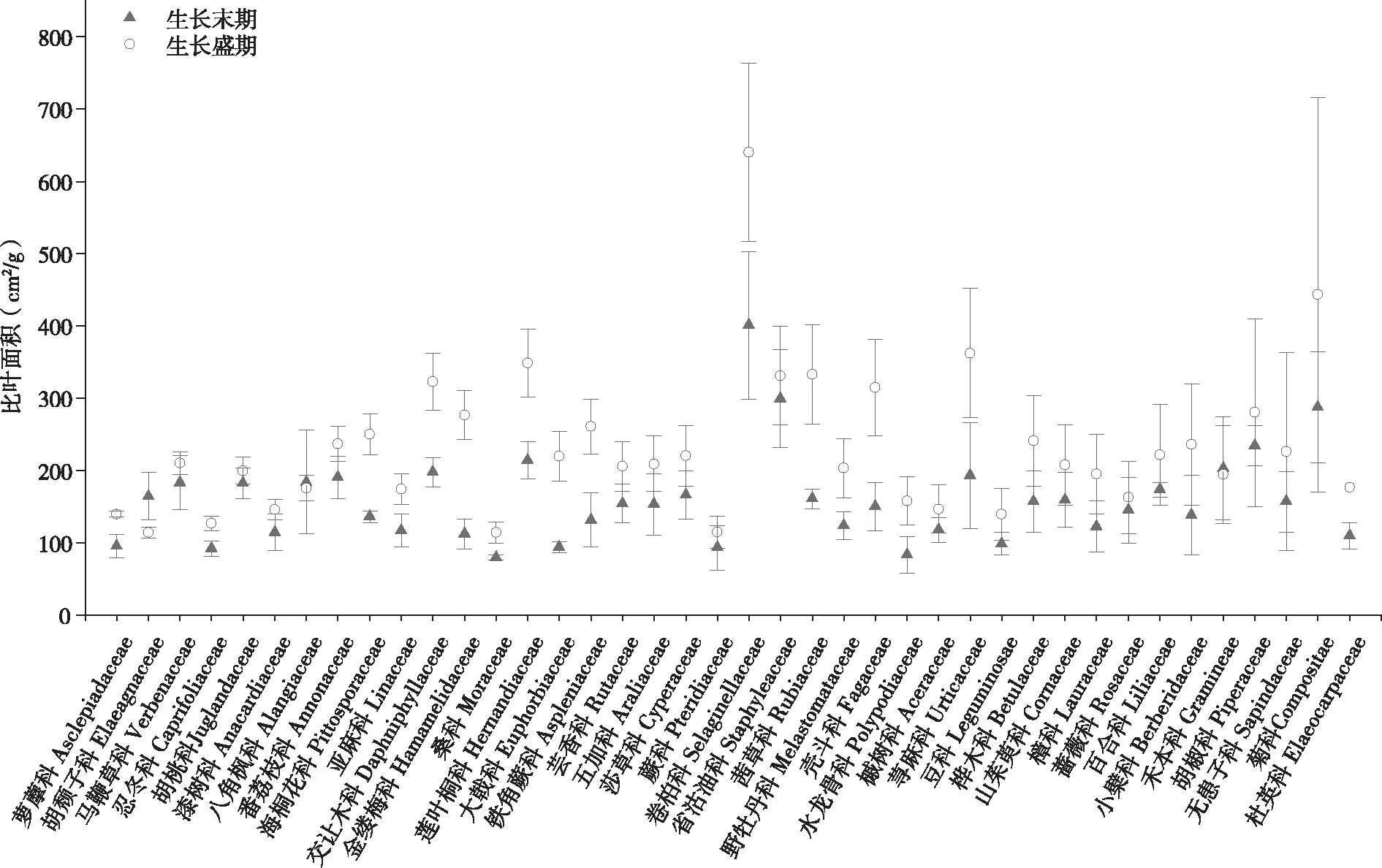
其中:弱变异性CV≤20.0%,中等变异性20.0% 图1 植物叶面积、叶干重和比叶面积的频率分布 68种植物不同生长阶段(生长盛期和生长末期)叶性状之间的相关关系均表现为叶面积与叶干重间呈极显著正相关(P<0.01),比叶面积与叶干重间呈极显著的负相关(P<0.01),叶面积和比叶面积间无相关关系(P>0.05)。 从生活型看,各生长阶段比叶面积均表现为草本显著高于灌木及乔木(P<0.01),比叶面积平均值分别为草本(264.84 cm2/g)、灌木(189.29 cm2/g)和乔木(173.76 cm2/g),乔木和灌木之间差异不显著(P>0.05)。从生长阶段看,生长盛期 > 生长末期,不同生活型乔木、灌木、草本在生长盛期和生长末期间均差异显著(P<0.05)。 图2 不同生活型植物的比叶面积 对落叶和常绿类型植物比叶面积比较见图3, 落叶乔木(182.45 cm2/g)>常绿乔木(166.51 cm2/g),落叶灌木(165.79 cm2/g)>常绿灌木(140.88 cm2/g)。落叶植物和常绿植物间均差异显著(P<0.05)。从生长阶段看,均为生长盛期 > 生长末期,落叶和常绿类型植物生长盛期和生长末期间均差异显著(P<0.05)。 图3 常绿和落叶类型植物的比叶面积 以生长末期为例,分析各生活型植物的比叶面积在不同群落样地中的差异性(见图4)。草本植物的比叶面积排序为乔林群落>顶级常绿落叶阔叶混交林群落>灌木灌丛群落>草本群落,草本植物的比叶面积在乔林群落和顶级常绿落叶阔叶混交林群落间差异不显著(P<0.05),其余群落间均差异显著(P>0.05)。灌木植物的比叶面积排序为灌木灌丛群落>顶级常绿落叶阔叶混交林群落>乔林群落,灌木植物的比叶面积在灌木灌丛群落与乔林群落间差异显著(P<0.05),其余群落间差异不显著(P>0.05)。乔木的比叶面积排序为顶级常绿落叶阔叶混交林群落>乔林群落>灌木灌丛群落,顶级常绿落叶阔叶混交林群落与乔林群落、灌木灌丛群落间差异显著(P<0.05)。 图4 在不同群落样地中各生活型植物比叶面积比较 不同系统发育类型植物的特点各个生长阶段比叶面积均表现出蕨类植物>种子植物,其比叶面积平均值分别为285.08 cm2/g和191.39 cm2/g,但两者之间差异不显著(P>0.05)。就生长阶段来看,蕨类植物和种子植物比叶面积均为生长盛期>生长末期,且不同生长阶段间均差异显著(P<0.05)。 单子叶植物和双子叶植物比叶面积在不同生长阶段表现不同,生长末期比叶面积表现为单子叶植物(204.46 cm2/g)>双子叶植物(156.24 cm2/g),且两者之间差异显著(P<0.05),生长盛期则相反,双子叶植物(224.08 cm2/g)>单子叶植物(198.91 cm2/g),且差异不显著(P>0.05)。双子叶植物比叶面积生长盛期>生长末期,且不同生长阶段间差异显著(P<0.05),单子叶植物比叶面积生长末期 > 生长盛期,且两阶段间差异不显著(P<0.05)。 图5 不同系统发育植物的比叶面积 所采集植物分属40科,比叶面积在不同科植物间多数差异显著(P<0.05)。从不同生长阶段来看,37科植物的比叶面积均表现为生长盛期 > 生长末期,仅八角枫科(Alangiaceae)、禾本科(Gramineae)、胡颓子科(Elaeagnaceae)表现为生长末期 > 生长盛期。生长盛期、生长末期比叶面积均以卷柏科(Selaginellaceae)为最高640.09cm2/g和401.07 cm2/g,分别以胡颓子科为114.17 cm2/g和金缕梅科(Hamamelidaceae)为最低79.49 cm2/g,变异系数范围分别为3.0%~60.97%和4.71%~39.05%。 喀斯特区68种植物比叶面积变化范围介于67.66~783.76 cm2/g,平均为180.70 cm2/g。比叶面积数值大小直接受叶片厚度、形状和重量等的影响,在一定程度上反映了叶片截获光的能力和在强光下的自我保护能力[15]。叶片的比叶面积越大越有利于增强植物捕获资源的能力,越小则有利于提高养分利用率和水分保存率[16]。对比其他地区研究结果,桂北岩溶区植物叶片比叶面积变化范围为62.56~530.04 cm2/g[17],喀斯特石漠化区比叶面积 的变化范围101.89~195.68 cm2/g[18],淮北相山5种优势植物比叶面积的变化范围160.97~305.01 cm2/g[19],山东桑沟湾19种植物比叶面积的变化范围172.7~315.8 cm2/g,变异系数为27.8%[20],海南岛研究101个物种比叶面积的变化范围为38~232 cm2/g[21]。可以看出,喀斯特区植物比叶面积数值处于中等偏上水平,且变化范围广、变异系数较大。说明喀斯特地区植物复杂程度高,性状值较分散,性状的可塑性较强[22],能消除不具竞争力的叶片投资策略,通过调节自身的物种组成,形成不同的功能性状组合来适应环境。 图6 不同科植物的比叶面积 从生活型看,草本植物的比叶面积显著高于乔木和灌木植物。山东桑沟湾植物比叶面积在3种生活型间从大到小依次为1 a生草本植物>多年生草本植物>木本植物[20]。对长白山研究表明,相对于乔木、灌木的比叶面积普遍较大且种间差异小[23]。大青沟保护区147物种的研究表明,草本植物的比叶面积最大,且与乔木和灌木差异显著[24]。本研究与以上研究结果一致,不同生活型植物对环境的适应能力明显不同,乔木和灌木会分配较多的生物量和氮于细胞壁,以增强叶片韧性,同时积累较多的光合产物为越冬和第二年的生长做准备,而分配到光合器中的氮则较少,光合能力较低,乔木和灌木叶片比叶面积低是牺牲了部分光合能力,而使自身得到较好保护,延长了寿命,是植物在适应环境时,叶片构建成本支出和回收的平衡。草本植物则分配较多的有机氮于光合器官,光合器官同化二氧化碳的速率也明显提高,植物生长加快,导致细胞内结构和非结构碳水化合物含量降低,从而植物采取薄叶片高比叶面积的性状[24-26]。且草本植物处于木本植物的下方,为了克服遮阴的不利影响,获得更多的光照资源,叶片变薄变大,叶密度降低,导致比叶面积增大[20,24],拥有快速获取资源的有利条件。草本植物在乔林群落比叶面积较大,灌木植物在灌木灌丛群落中较大,乔木在顶级常绿落叶阔叶混交林群落中较大。乔木群落及顶级常绿落叶阔叶混交林群落在长期的自然演替过程中已达到与生境条件相互适应的动态平衡过程,是一种小生境条件作用下的隐域性稳定植被系统[24],而草本群落处于相对不稳定状态中。可见植物在不同的群落会采取不同的生态策略来适应环境。 本研究中落叶植物的比叶面积大于常绿植物,说明岩溶地区常绿植物叶片对光照的获取能力比落叶植物低,常绿植物的光合作用速率比落叶植物低。以下研究也有相同结论,桂北岩溶区常绿树种的比叶面积显著低于落叶树种,从大到小依次为落叶阔叶>阔叶灌木>常绿阔叶>针叶[19];东北次生林常绿针叶和落叶阔叶植物的比叶面积达到极显著差异[27]。因为常绿植物叶生命周期长,叶片大部分干物质用于构建保卫细胞,增加叶厚或叶肉细胞密度,用以防御虫食,还可以防止过度失水,较低的比叶面积可维持植物的生长和繁衍[28-29]。落叶植物叶生命周期较短,仅在生长季内合成光合产物,通过扩大单位重量叶面积提高光捕获量,通过增大比叶面积提高瞬时生产率以补偿生长和维护叶片的成本,并最大程度地给植物提供光合产物,气候条件适宜时在较短的时间内完成植物的快速增长[28]。2 结果与分析
2.1 植物叶功能性状的总体分布统计特征

2.2 不同生活型植物比叶面积比较

2.3 不同系统发育类型植物比叶面积比较

2.4 不同科植物的比叶面积
3 结论与讨论
猜你喜欢
建筑与预算(2022年6期)2022-07-08
农村实用技术(2020年1期)2020-12-16
乡村科技(2018年8期)2018-02-13
成才之路(2017年21期)2017-07-27
安徽农学通报(2017年9期)2017-05-19
现代农业科技(2016年22期)2017-03-24
诗潮(2017年2期)2017-03-16
湖南林业科技(2017年1期)2017-02-06
学周刊·下旬刊(2015年10期)2015-07-22
河南科技(2014年16期)2014-02-27
