
不同温度对皂荚幼苗生长和生理特性的影响
2022-12-07 08:41刘富华王秀荣何可权
山地农业生物学报 2022年6期
刘富华,王秀荣,赵 杨*,何可权
(1.贵州大学 林学院/贵州省森林资源与环境研究中心,贵州 贵阳 550025 ;2.独山县国有林场,贵州 独山 558200)
皂荚(Gleditsiasinensis)是豆科皂荚属落叶乔木,适应性强,分布广,是优良的经济生态树种,是贵州省重要的特色经济林木,于2018年被列入特色林业来重点发展,以毕节为主的喀斯特石漠化地区为其主要种植区,带动全省皂荚产业实现标准化生产、产业化经营、规模化发展。贵州省地貌类型复杂,海拔高度变化大,立体气候明显,省内多地苗圃所育皂荚苗木生长差异明显。温度是影响植物光合作用的关键因子之一,是决定其地理分布范围、生长发育、果实产量及质量的主要因素。当温度高于或低于临界温度时,植物体内代谢失衡、活性氧、自由基和丙二醛(MDA)积累,造成膜透过性上升,电解质外渗,细胞膜系统严重受损,同时光系统Ⅱ(PSⅡ)受到一定伤害,原初光能转换效率下降,电子传递活性减弱,光合作用受到抑制,阻碍植物生长[1]。木薯(ManihotesculentaCrantz)[2]幼苗受到高温胁迫时气孔导度、蒸腾速率和净光合速率显著下降。低温下绿萝(Scindapsusaureus)[3]和毛白杨(PopulustomentosaCarr)[4]幼苗叶绿素含量降低,龙葵(SolanumnigrumL.)[5]幼苗Fv / Fm、qP均降低,qN增大。当温度超出20~30 ℃范围时,红心衫(Cunninghamialanceolata)[6]、糖胶(Alstoniascholaris)[7]、辣木(M.oleiferaLam.)[8]幼苗MDA、可溶性糖、可溶性蛋白、脯氨酸含量和超氧化物歧化酶(SOD)、过氧化物酶(POD)酶活性均显著升高,生长指标、叶绿素含量、光合作用和叶绿素荧光参数都会受到抑制,严重影响幼苗生长。
随着全球气候变暖,温室效应加剧,贵州地区春夏季气候异常,春寒和高温时有发生,皂荚幼苗大面积死亡时有发生,制约着皂荚培育的发展。皂荚已成为重要的经济生态树种,大规模批量种植已成趋势,国内对皂荚的价值、栽培或改造技术方面报道较多。近几年,贵州有大量引进北方皂荚种源,在引种栽培过程中温度是重点考虑的因素,开展皂荚对不同温度环境生态适应性方面的研究却鲜有报道,尤其在幼苗对不同温度中的生理响应方面未有研究。幼苗是植物重要的生活史阶段,幼苗的生长温度是其能否存活的关键因素。为此,本文探索不同温度下生长的皂荚幼苗生长指标、光合特性和抗性生理指标的差异,寻找适合的育苗温度,为皂荚繁育提供参考依据,为皂荚幼苗培育温度调控提供理论依据与指导,促进皂荚幼苗标准化生产。
1 材料与方法
1.1 供试材料与试验设计
供试皂荚种子采摘于贵州省贵阳市花溪区一株野生大树(106°39′27″E,26°27′13″N),经净种催芽处理[9],于2021年3月15日进行容器育苗。3个月后选取长势一致的幼苗,在人工气候室内选取5个条件相同的恒温培养箱中培养,均分为三层。温度分别设置为15 ℃、20 ℃、25 ℃、30 ℃、35 ℃,每个处理设置18盆苗,3次重复。每盆种植一株幼苗,并定时更换每个重复苗木的摆放位置,培养90 d。光照强度300 μmol/m2·s、相对湿度55%、光周期设置为光期∶暗期=12∶12(h),定时浇水保持土壤湿润。
1.2 测定指标与方法
1.2.1苗高、地径的测量
测量苗高和地径均随机选取6株取平均值。苗高用直尺测量,精确到0.1 cm;地径用游标卡尺测量,精确到0.01 mm。
1.2.2光合色素含量测定
光合色素采用分光光度法[10]测定。将采集的新鲜叶片去除叶脉剪碎混匀,精确称量0.2 g(精确度0.001 g)放入干燥研钵中,加入液氮快速研磨至粉末状。先后加入95%乙醇3 mL和5 mL,继续研磨成匀浆状后静置3~5 min,将匀浆过滤后的液体全部移入25 mL棕色容量瓶中,并用95%乙醇定容,摇匀后静置12 h。以95%乙醇为空白对照,使用UV-1800紫外分光光度计在665、649和470 nm波长下测定叶绿素a(Chla)、叶绿素b(Chlb)和类胡萝卜素(Car)的吸光度。按照公式计算每克叶片的叶绿素a、叶绿素b和类胡萝卜素含量,每个温度处理测定设置3次重复取平均值。
Chla=(13.95A665-6.88A649)×V/l000 m
Chlb=(24.96A649-7.32A665)×V/l000 m
Car=[(1000A470-2.05Chla-114.8Chlb)/245]×V/l000 m
总叶绿素=叶绿素a+叶绿素b
1.2.3叶绿素荧光动力学参数测定
每个处理每个重复中选取一盆从上往下第2~4片健康良好的2片叶进行标记,于21∶00~23∶00,叶片暗适应40 min 后,用便携式荧光仪(Junior-PAM-Ⅱ)对皂荚幼苗叶片先进行慢速诱导动力学曲线测定,测定最小荧光(Fo)、最大荧光(Fm),随后设置0~1500 μmol/m2·s范围的9个梯度的光强进行快速光响应曲线参数测定[11],测定PS Ⅱ最大光化学效率(Fv/Fm)、实际光化学量子产量(Fv′/ Fm′)、PS Ⅱ实际光化学量子效率(YⅡ)、光化学淬灭系数(qP)和非光化学淬灭系数(qN NPQ)、调节性能量耗散Y(NPQ)和非调节性能量耗散Y(NO)、相对电子传递速率(ETR)、最大电子传递速率ETRmax、初始斜率(α)、光抑制参数(β)和半饱和光强(Ik)。
1.2.4生理指标测定
剪取每株幼苗3~4片叶片剪碎混匀,去除主脉贮藏于-80 ℃的冰箱中用于叶片生理生化指标的测定。参照李合生[12]的方法,采取硫代巴比妥酸比色法测定MDA的含量;采用考马斯亮蓝法测定可溶性蛋白质含量;采用蒽酮比色法测定可溶性糖含量;采用酸性茚三酮法测定游离脯氨酸含量;采用紫外分光光度法测定 CAT 酶活性;采用氮蓝四唑(NBT)法测定 SOD 酶活性;用愈创木酚法测定POD 酶活性。参照Nakano[13]的方法测定抗坏血酸过氧化物酶(APX)活性。各项指标均测定 3次,取平均值。
1.3 数据分析
采用Excel 2019软件进行数据处理,Origin2021软件进行图形绘制。采用 IMB SPSS Statistics 26 软件进行单因素方差分析,采用 Duncan进行多重比较和差异显著性检验(P<0.05)。
2 结果与分析
2.1 不同温度处理对皂荚幼苗生长指标的影响
如图1所示,皂荚幼苗苗高和地径两个指标均随温度升高呈现先上升再下降的趋势。25 ℃和30 ℃处理下的苗高和地径均显著高于其他3个处理,其中,25 ℃处理最大,15 ℃处理最小。25 ℃处理苗高和地径分别为14.8 cm和3.50 mm,15 ℃处理苗高和地径分别为12.2 cm和3.00 mm。苗高和地径均表现为25 ℃>20 ℃>30 ℃>35 ℃。
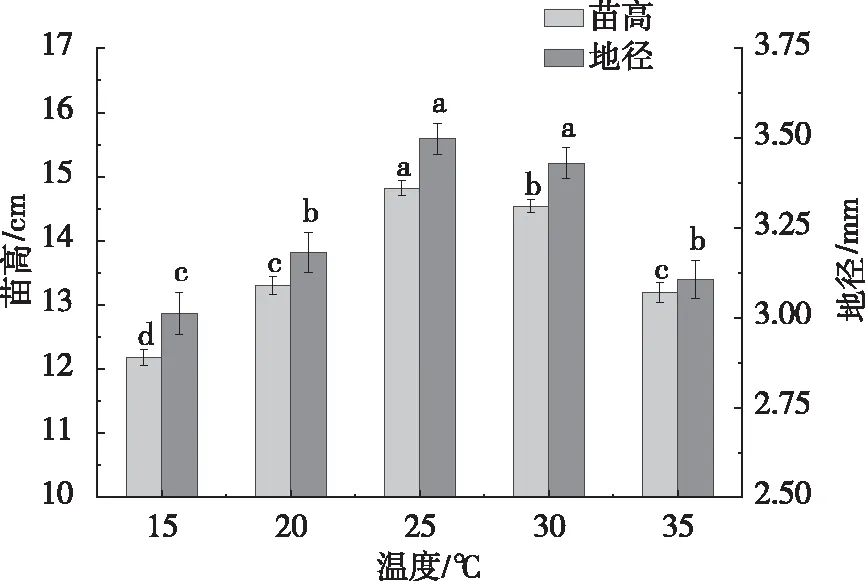
注:不同小写字母表示各处理间差异显著(P< 0.05)。图1 不同温度对皂荚幼苗生长的影响Fig.1 Effects of different temperatures on the growth of G.sinensis seedlings
2.2 不同温度对皂荚幼苗叶绿素含量的影响
由表1可知,不同温度对皂荚幼苗叶片叶绿素含量有显著的影响。在15~30 ℃范围内随温度的升高皂荚幼苗叶片中叶绿素 a、叶绿素 b、类胡萝卜素和总叶绿素含量均升高,叶绿素 a、叶绿素 b、类胡萝卜素、总叶绿素含量在25 ℃和30 ℃处理急剧上升,在35 ℃处理时轻微下降。25 ℃处理叶绿素 a、叶绿素 b、类胡萝卜素、总叶绿素含量均为最高,分别为0.787 mg/g、0.289 mg/g、 0.351 mg/g、 1.076 mg/g。叶绿素a/b 质量分数随温度升高而下降。
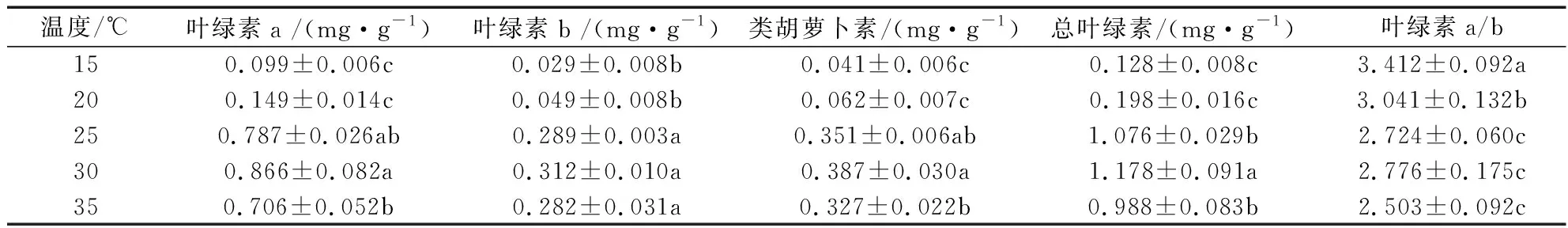
表1 不同温度对幼苗叶片叶绿素含量的影响
2.3 不同温度对皂荚幼苗叶片叶绿素荧光参数的影响
2.3.1不同温度处理下 Fo、Fm、Fv/Fm与Fv/Fo的比较
由表2可知,随温度升高,皂荚幼苗叶片Fo、Fm、Fv/ Fm、 Fv/ Fo均表现出先上升后下降的趋势。25 ℃处理的Fo、Fm、Fv/Fm、潜在光化学活性(Fv/ Fo)最高,显著高于其他处理(P<0.05),其次是20 ℃和30 ℃处理。15 ℃和35 ℃处理Fo、Fm、Fv/Fm、Fv/ Fo较低。
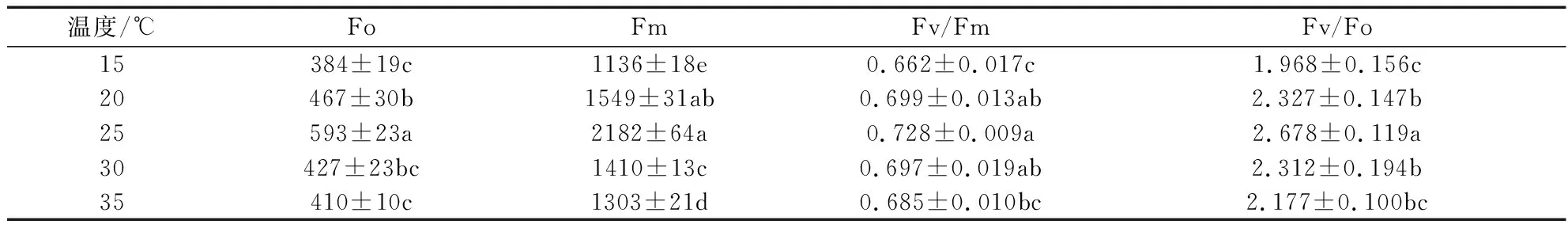
表2 不同温度叶绿素荧光参数
2.3.2不同温度下PSⅡ反应中心参数变化比较
由图2可知,在光响应变化过程中,不同温度处理的Fv′/ Fm′和Y(Ⅱ)均随着光合有效辐射(PAR)升高而降低,Fv′/ Fm′下降幅度较小。25 ℃处理Fv′/ Fm′和Y(Ⅱ)最高,显著高于其他处理(P<0.05)。Fv′/ Fm′和Y(Ⅱ)均随温度升高而先升高后降低。

图2 不同温度有效光化学量子产量 Fv′/ Fm′和实际光合量子产量 Y(Ⅱ)和的光响应Fig.2 Light response of effective photochemical quantum yield Fv′/Fm′ and actual photosynthetic quantum yield Y(Ⅱ) and under different temperature
由图3可知,不同温度下qP随着 PAR的增加而减小,在低光强下,qP、NPQ和qN变化比较大,在625~1500 μmol/m2·s内逐渐稳定。不同温度处理qP值随温度升高而先升高后下降25 ℃处理的qP值最大,15 ℃处理qP值最小。不同温度处理的NPQ和qN随着PAR的增加而增大。15 ℃处理NPQ和qN最大,显著高于其他处理,35 ℃处理NPQ和qN最小。

图3 不同温度光化学淬灭系数qP与非光化学淬灭系数(NPQ 、qN)的光响应Fig.3 Light response of photochemical quenching coefficient qP and non-photochemical quenching coefficient (NPQ,qN) at different temperatures
由图4可见,不同温度处理的Y(NPQ)值随PAR增加而逐渐升高,PAR<420 μmol/m2·s时,各处理的Y(NPQ)值急剧上升,当 PAR>420 μmol/m2·s之后Y(NPQ)值趋于饱和。Y(NPQ)在整个变化过程中,15 ℃处理的Y(NPQ)显著高于其他处理(P<0.05),其他处理之间Y(NPQ)差异不显著,30 ℃处理Y(NPQ)最小。Y(NO)随PAR的增加整体波动较小,30 ℃处理Y(NO)值最大,15 ℃处理的Y(NO)值最小。
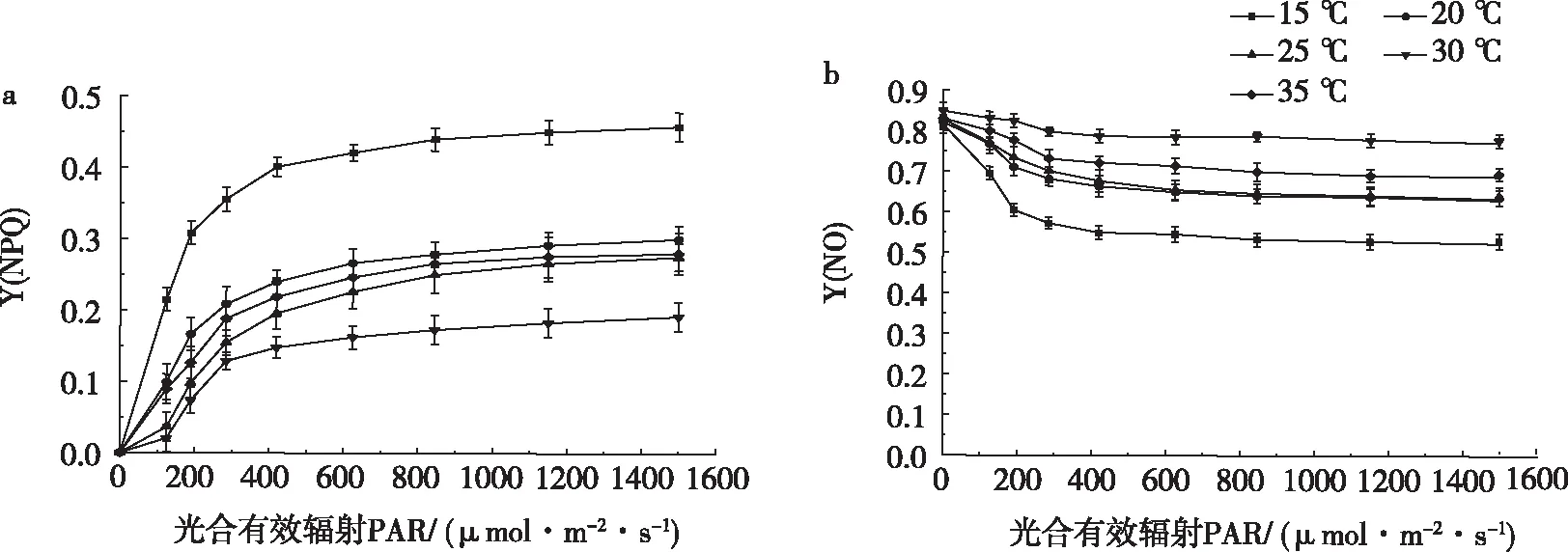
图4 不同温度调节性Y(NPQ)和非调节性Y(NO)能量耗散参数的光响应Fig.4 Light response of different energy dissipation parameters of temperature-regulated Y(NPQ) and non-regulated Y(NO)
由图5可知,低光强下各处理电子传递速率(ETR)值迅速升高,当 PAR>420 μmol/m2·s时,各处理ETR值趋于饱和,各处理之间差值稳定。不同温度处理的ETR随PAR的增加而呈现先上升后下降的趋势,25 ℃处理ETR值最高,15 ℃处理ETR值最小。25 ℃处理和20 ℃处理ETR值较高。
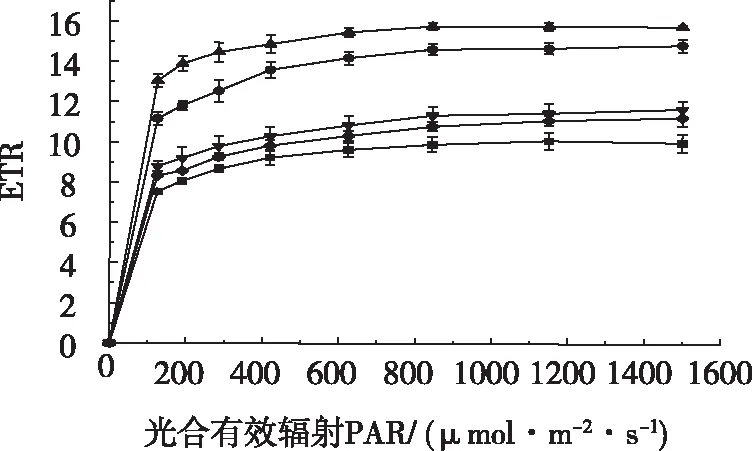
图5 不同温度电子传递速率(ETR)的光响应Fig.5 Light response of electron transfer rate (ETR) at different temperatures
由表3可知,随着温度升高,最大电子传递速率(ETRmax)、初始斜率α、光抑制参数β、半饱和光强Ik值呈先上升后下降的趋势,各处理之间差异显著。25 ℃处理的 ETRmax、α、β、Ik最高,均显著高于其他处理(P<0.05)。15 ℃处理ETRmax、α、β、Ik最低。
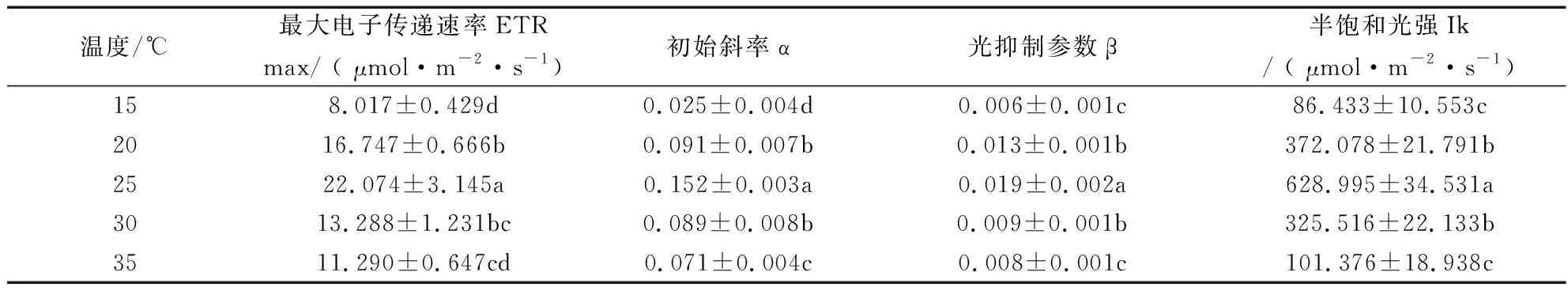
表3 不同温度光响应曲线拟合参数
2.4 不同温度对皂荚幼苗叶片生理指标的影响
2.4.1不同温度对皂荚幼苗叶片MDA和渗透调节物质含量的影响
随温度升高,皂荚幼苗叶片MDA含量呈先降后升趋势(表4)。在 20 ℃和25 ℃处理MDA 含量保持较低水平, 15 ℃和35 ℃处理MDA 含量显著增加,35 ℃处理MDA 含量最高。不同温度处理对幼苗可溶性蛋白、可溶性糖和脯氨酸含量有显著影响。随温度升高,可溶性蛋白、可溶性糖和脯氨酸含量呈现先降低再升高的趋势。在20~30 ℃范围内可溶性蛋白、可溶性糖和脯氨酸含量差异较小,均保持较低水平。15 ℃和35 ℃处理可溶性蛋白、可溶性糖和脯氨酸含量显著高于其他3个处理(P<0.05),均保持较高水平,15 ℃和35 ℃处理脯氨酸含量分别是25 ℃的1.74倍和1.59倍。15 ℃处理可溶性蛋白、可溶性糖和脯氨酸含量均为最高,25 ℃处理可溶性蛋白、可溶性糖含量最低。

表4 不同温度对皂荚幼苗叶片MDA和渗透调节物质含量的影响
2.4.2不同温度对皂荚幼苗抗氧化酶活性的影响
在 20~30 ℃范围,幼苗保护性酶CAT、SOD、POD和APX活性均保持较低水平(表5)。 随着温度的升高或降低,保护性酶活性逐渐增强,其中CAT和SOD活性的增强更为显著 。15 ℃和35 ℃处理CAT、SOD、POD和APX活性均显著高于其他3个处理。25 ℃处理CAT、SOD、POD和APX活性分别为624.31 μmol/g·min、161.07 U/g·min、41.67 U/g·min、50.01 μmol/g·min。相比较25 ℃处理,15 ℃和35 ℃处理CAT升高至1.9倍和1.6倍;SOD升高至5.7倍和6.8倍;POD升高至1.7倍和2.2倍;APX升高至1.2倍和1.32倍。
3 结论与讨论
不同温度下皂荚幼苗的生长指标、光合特性及生理指标不同。通过对比幼苗的生长指标、光合特性及生理指标可知,25 ℃是皂荚育苗最适宜的温度。超出20~30 ℃范围幼苗的生长指标会明显降低,光合作用下降,MDA含量、渗透调节物质含量和抗氧化酶活性会显著上升,不利于幼苗生长发育。

表5 不同温度对皂荚幼苗抗氧化酶活性的影响Tab.5 Effects of different temperatures on antioxidant enzyme activities of G.sinensis seedlings
在高温和低温环境条件下,植物光合作用随着温度胁迫而下降,光合作用相关的酶活性降低,光合作用受到限制,导致植物营养物质的合成与积累受阻,从而影响植物苗髙、地径的生长。辛福梅等[14]研究发现高温和低温胁迫均抑制巨柏(Cupressusgigantea)幼苗的生长,导致株高、地径、生物量和生根数量均有所降低。本试验中,15 ℃和35 ℃对皂荚幼苗苗高和地径有明显的抑制作用。比较苗高、地径可知,皂荚幼苗最适宜生长在25 ℃,在15 ℃和35 ℃下生长较差。
温度对叶绿素含量有一定的影响,陈青君等[15]发现,温度胁迫会使黄瓜(CucumissativusL.)幼苗叶绿素含量显著下降,与本试验研究趋势一致。在低温时,酶促反应降低,引起光合反应降低,导致叶绿素含量下降,这可能是造成15 ℃时皂荚叶片叶绿素含量比较低的原因。25 ℃和30 ℃处理叶绿素含量保持较高水平,说明皂荚幼苗在此温度下光合作用较强,适宜在此温度范围内生长。叶绿素的合成需要绝大部分酶的参与,在15 ℃和35 ℃处理时光合色素较低,可能是由于温度胁迫致使酶活性下降甚至失活导致光合色素合成减少,或是叶绿体自身功能的紊乱造成,叶绿素的分解速率加快,分解大于合成,也可能是叶绿素合成的中间产物5-氨基酮戊酸和卟啉IX的生物合成,降低叶绿素生成量[1]。叶绿素a/b值随温度升高明显下降,低温下生长的罗勒(Ocimumbasilicum)幼苗叶绿素a/b比值明显高于高温下生长[16],这与本试验研究趋势一致。叶绿素a/b比值的降低,是由于温度的升高,叶绿素a被氧化成叶绿素b,使叶绿素b的含量上升所致。
叶绿素荧光参数可以反映光合作用热耗散情况和环境因子对植物光合作用及胁迫响应的影响[17],表示其抗逆性[18]。Fm反映PSⅡ的电子传递情况;Fv/Fo可衡量PSⅡ潜在光合活性;Fv′/Fm′反映实际捕获能量的传递效率;Y(Ⅱ)表示实际光能转化效率[19]; Y(NPQ)代表光保护能力。本试验研究表明,25 ℃处理的皂荚幼苗的Fm、Fv/ Fo、Fv/ Fm、Fv′/Fm′、Y(II)和qP高于其他处理组,说明25 ℃处理皂荚幼苗PSⅡ反应中心光合原初反应能量转化效率和光合电子传递速率较高,实际光合效率最高,光合作用较强,PSⅡ天线色素吸收的光能用于光化学电子传递份额较高。而15 ℃和35 ℃处理较低,说明高温和低温环境下幼苗受到了光抑制作用,导致 PSII光合效率和实际量子产量均受到抑制,幼苗光合结构以及 PSⅡ活性中心损害,天线色素捕获的光能无法推动光合电子的传递,电子传递受阻,光能利用率下降,这与王璐等[20]的研究结果一致。
皂荚幼苗在光响应过程中,提高qN、NPQ减少光能对光系统的破坏。在15 ℃处理下的qN最高,说明此温度下通过qN调节过量能量耗散的能力下降,低温下幼苗将大部分吸收而来的光能用于热耗散,这与杨盛昌等[21]研究低温对红树(Rhizophoraapiculata)的影响趋势一致。qN和NPQ随着温度的升高而降低,说明随温度升高幼苗将过剩光能热耗散能力下降。对比qP、qN和NPQ可发现,皂荚幼苗叶片荧光参数发生淬灭方式主要为非光化学淬灭。随着 PAR 的增加,Y(NPQ) 明显增加,表明幼苗通过调节 Y(NPQ) 来减少过剩光能对光系统结构的损害,与钱永强等[22]研究银芽柳(Salixleucopithecia)趋势一致。Y(NO)表示光损伤程度,皂荚幼苗在30 ℃以上时光损伤程度较高。25 ℃处理实际光量子产量最大,但Y(NPQ)、Y(NO)并不是最小的,15 ℃时光保护能力最大,光损伤最小,说明幼苗可以通过不同的作用机制进行适应性调节以抵御温度胁迫,这与李兵阳[7]研究趋势一致。幼苗可通过提高电子传递速率来响应能量过多耗散带来的影响。25 ℃处理ETR 、ETRmax 、光能利用效率(α) 、光抑制参数(β) 和光耐受能力(Ik)最高,说明其电子传递链传递速率最高,光化学反应启动较快,耐光抑制的能力较强,15 ℃和35 ℃处理耐光抑制能力较弱。
可溶性糖、可溶性蛋白和脯氨酸是植物体内重要的渗透调节物质,能够降低水势,增加持水力,防止细胞过度失水,从而减少低温和高温环境对植物体造成的伤害,并提高抗寒性和抗高温性[1]。15 ℃和35 ℃处理可溶性糖、可溶性蛋白和脯氨酸含量显著增加,说明此温度胁迫下幼苗会受到伤害,从而主动积累会通过增加渗透调节物质含量来调节适应环境温度胁迫,这与张天翔等[8]和韩晓燕等[27]的研究趋势一致。可溶性蛋白含量增加,可能是受温度胁迫糖酵解途径转变为磷酸戊糖途径,从而产生较多的NADPH和ATP,有利于蛋白质的合成[1],或是幼苗体内诱导产生了与抗寒、抗高温有关蛋白的合成,促使幼苗增加抗逆性。可溶性含量增加,可能是温度胁迫使淀粉等大分子物质水解成可溶性糖的速率加快。
本试验对皂荚幼苗经过不同温度处理90 d后的生长指标、光合色素和生理指标进行比较,初步探索了皂荚幼苗对不同温度处理的生理响应。若要更深入研究探讨皂荚幼苗的抗寒性和抗热性的生理生化机制,需胁迫下皂荚生理指标的动态变化测定记录,做进一步的探索。本试验幼苗的生长环境是人工模拟环境,与自然环境相比,影响因素较少,若要探究适合的育苗环境,需要增加多种因素综合分析比较。本文未对不同种、不同种源的皂荚进行比较分析研究,在引进其他种源之前应进行试验并分析其可行性。
皂荚幼苗生长在不同温度下其生长指标、光合特性及生理指标表现不同。25 ℃处理苗高地茎值最大,叶绿素含量最高,光合作用较强,实际光合量子产量较高,光损伤程度较轻,耐光抑制能力较强。MDA含量、渗透调节物质含量和抗氧化保护酶活性随温度升高呈现先下降后升高的趋势。15 ℃和35 ℃处理幼苗MDA、渗透调节物质含量和抗氧化保护酶活性急剧上升,以提高抗逆性。25 ℃下皂荚幼苗不易受活性氧和自由基危害,渗透调节作用平衡,是最适宜的育苗温度。
猜你喜欢
阅读(科学探秘)(2020年8期)2020-11-06
中国果业信息(2019年1期)2019-01-05
中成药(2018年8期)2018-08-29
生物学教学(2017年9期)2017-08-20
人生与伴侣·共同关注(2016年11期)2016-04-13
幸福·婚姻版(2016年1期)2016-03-07
医学研究杂志(2015年9期)2015-07-01
中国当代医药(2015年9期)2015-03-01
现代检验医学杂志(2015年6期)2015-02-06
食品工业科技(2014年6期)2014-05-10
