
凤丹栽培群体表型多样性分析及综合评价
2022-12-05 06:54谭万庆彭丽平刘政安佟宁宁宋苗苗舒庆艳
经济林研究 2022年3期
谭万庆,彭丽平,刘政安,佟宁宁,宋苗苗,舒庆艳
(1.中国科学院 植物研究所,北京 100093;2.中国科学院大学,北京 100049)
凤丹Paeonia ostii是芍药科Paeoniaceae 芍药属Paeonia牡丹组(Sect.Moutan)的多年生落叶灌木,其成熟种子的出油率和不饱和脂肪酸的含量均较高[1-3],其中α-亚麻酸的含量最高达42%[4],而α-亚麻酸是人体必需的不饱和脂肪酸之一[5],能够在人体健康和发育中发挥重要作用。2011年凤丹和紫斑牡丹籽油被批准为新资源食品,2014年国务院68 号文件把油用牡丹列入国家重点推广的木本油料作物之一,极大地推动了油用牡丹产业的发展。作为油用牡丹主要栽培系列之一的凤丹,在河南、山东和陕西等地均有种植。目前,关于凤丹的研究主要是围绕种子发育[6]、脂肪酸成分[7]及其合成途径中关键基因的功能[8-9]和栽培模式[10]等方面展开的。然而,由于缺乏统一的苗木选择标准,凤丹籽不同地区的产量差异明显,甚至同一地区或同一栽植区域不同单株间的产量也存在明显差异,因而立足于同一栽培环境的群体表型多样性分析及综合评价,明确凤丹优良种质的选择指标和重点改良指标具有重要意义。其中,单株种子质量是凤丹单株产量的直接体现,而角数在不同株间存在广泛变异,两者可作为分析过程中的重点关注性状。
对表型性状指标与产量指标进行相关分析、回归分析和通径分析等,有利于发现影响产量的关键表型性状,其可作为高产种质选育的重要指标,而主成分分析是对不同指标进行综合评价的常用方法之一。杨玉霞等[11]在对金果榄主要农艺性状与产量的相关性分析、回归及通径分析中发现,产量高的株型应是块根个数多、块根单个质量高、块根长及茎叶质量适中的植株;杨涛等[12]利用逐步回归分析法分析发现,株高、始节高、果枝数、单铃皮棉质量和单株籽棉质量这5 个指标均为海岛棉种质评价的关键指标;王彦花等[13]采用主成分分析法建立了油茶综合品质评价模型;而刘俊涛等[14]等结合相关性分析、主成分分析及聚类分析筛选出了3 株千年桐最优单株。目前,有关油用牡丹表型多样性的研究对象多为紫斑牡丹[15-17]和多个牡丹品种[18-19],而以凤丹为供试材料的研究报道相对匮乏,且相关研究很少涉及单株产量[20-21]。崔虎亮等[18]对凤丹和其他24 个牡丹种质的主要表型性状和单株产量的相关性进行了研究,综合分析了影响其单株产量的关键因素。但是,到目前为止,关于同一生境和栽培方式下凤丹实生群体的表型(尤其是产量性状)多样性分析和综合评价的研究鲜见报道。为了充分挖掘对单株产量影响较为显著的表型性状,为凤丹优良种质的筛选提供科学依据,为油用牡丹高产品种选育奠定基础,本研究以河北地区的凤丹栽培群体为研究对象,调查其单株种子质量、复果种子数、复果种子质量和复果质量等果实性状,以及株高、冠幅和叶面积等生长性状,共计26 个表型性状;并基于其丰富的表型多样性,通过相关性分析、聚类分析、通径分析及主成分分析对其进行综合评价,现将研究结果分析报道如下。
1 材料和方法
1.1 试验材料
2019年选择河北省唐山市迁西县油用牡丹基地(118°24′E,39°58′N)内长势良好、结实量较高且无病虫害的8年生凤丹实生苗植株进行挂牌标识,2020年对其中239 株的表型性状进行测定。
1.2 调查的性状指标及其测定标准
调查的性状分为生长性状和果实性状两大类,共计26 个性状指标,具体的性状指标和测定标准见表1。2020年5月上旬,对试验群体的株高和冠幅进行测量,每株随机选择3 个当年生枝条,测定其枝长和枝径,并记录复叶数;同时,选择枝条上的第3 片复叶,测定复叶长(宽)、叶柄长和顶小叶长(宽);并将叶片拍照后采用ImageJ 软件[22]测定叶面积。2020年8月底,在果实完全成熟后,收集并调查群体各单株的果实,首先称量每个单株所有的果荚质量(干质量),然后在每个单株中随机选择3 个果荚进行果实性状的测定,称量复果质量,记录角数;再在每个果荚中随机选择3 个角测定其单角长、单角宽和单角厚,记录单角种子数,并称量单角种子质量;然后将单个果荚的所有种子混合起来,记录复果种子的数量与质量,并用复果种子质量除以种子粒数得到单粒种子的质量;最后剥开所有果荚,称量单株种子质量。所有性状的随机选择均在去掉测定值极大和极小的表型性状的前提下进行。

表1 凤丹栽培群体的26 个表型性状及其测定标准Table 1 26 phenotypic traits and measurement standards of P.ostii cultivated population
1.3 数据处理及统计分析
分别采用Excel 2020、SPSS 24 和Origin 2021b等软件进行数据整理、统计与分析。对26 个性状指标进行描述性统计,计算其极差值、变异系数(coefficient of variation,CV)、Shannon-Weaver 遗传多样性指数(index of genetic diversity,H');然后对其分别进行相关性分析和聚类分析;并以单株种子质量作为因变量,以其他25 个性状指标的测定值为自变量,进行多元线性回归分析和通径分析,其中,回归分析得到的标准化系数为直接通径系数,各个性状指标和单株种子质量之间的简单相关系数等于直接通径系数与间接通径系数之和[23];同时以角数为因子,进行果实性状的单因素方差分析;最后以单株种子质量和主成分分析的综合得分为依据进行优良单株的筛选。
根据各个性状指标的平均值(μ)与标准差(σ),按照公式(μ±kσ)(k=0、0.5、1.0、1.5、2.0)划分各个性状指标的10 个等级,其中k表示等级划分系数;根据公式(pi=ni/n)计算不同性状变异的分布频率,式中的ni表示某一性状指标处于第i个变异的材料个数,n表示材料的总数,pi表示某一性状第i个变异的分布频率[24-25]。变异系数(CV)和Shannon-Weaver 遗传多样性指数的计算公式分别如下:
2 结果分析
2.1 凤丹栽培群体数量表型性状的变异分析
凤丹栽培群体表型变异分析结果(表2)表明,26个表型性状变异系数的变幅为10.50%~57.39%,平均值为26.65%;其中,株高的变异系数最小,单株果实质量的变异系数最大,单株种子质量的变异系数为56.23%,仅次于单株果实质量的变异系数。生长性状变异系数的平均值为18.69%,果实性状变异系数的平均值33.47%,其中果实质量相关性状的变异系数高于果实形态相关性状的变异系数。

表2 凤丹栽培群体26 个表型性状的描述性统计结果Table 2 Descriptive statistics of 26 phenotypic traits in P.ostii cultivated population
26 个表型性状多样性指数的变幅为1.15 ~2.09,其中,角数的多样性指数最小,株高和顶小叶长的多样性指数最大,均值为1.97;复叶数的多样性指数为1.77,而其余生长性状的多样性指数均大于2.00,较大的多样性指数表明生长性状在每一等级中的分布较为均匀[25];角数、单株果实和种子质量等8 个果实性状的多样性指数均小于2.00,较小的多样性指数表明这些性状在各等级中的分布不均匀。
单株果实质量的最小值为25.38 g,最大值为664.87 g,极差为其平均值的3.99 倍;单株种子质量的最小值为11.05 g,最大值为279.10 g,极差为其平均值的3.80 倍。单株果实及种子质量较大的变异系数和极差,表明凤丹栽培群体不同单株间产量差异显著,因而探讨影响凤丹单株产量变异的表型性状具有重要意义。
2.2 凤丹栽培群体表型性状的相关性分析
为了更好地解析凤丹栽培群体各个表型性状间的关系,对26 个表型性状指标之间的相关性进行了分析,结果如图1所示。

图1 凤丹栽培群体26 个表型性状之间的相关性Fig.1 Correlation of 26 phenotypic traits in P. ostii cultivated population
图1表明,在由26 个表型性状指标组成的325 对性状指标组合中,有272 对性状指标之间呈显著相关,其中有253 对性状指标之间呈极显著相关。在果实性状指标中,单角性状、复果性状与单株产量性状间均呈极显著正相关;坐果数与单角性状、复果性状和单株产量性状之间均呈显著相关;单粒质量与单角和复果种子数间均呈显著相关,而与单角及复果质量性状间均呈极显著相关。在其生长性状中,株高、冠幅长、冠幅宽、当年生枝长、当年生枝径、复叶长、复叶宽、叶面积、叶柄长和顶小叶长这10 个性状指标之间均呈显著相关,而除株高外的其余9 个性状相互之间均呈极显著相关;顶小叶宽与复叶长宽、叶面积、叶柄长之间均呈极显著相关;复叶数与当年生枝长之间呈极显著相关。不同类别的性状指标之间也都存在较强的相关性,如冠幅、当年生枝长、当年生枝径、复叶长、复叶宽、叶面积、叶柄长、顶小叶长与复果质量、复果种子数、复果种子质量及所有单角性状和单株产量性状之间均呈极显著相关,复叶数、当年生枝长、当年生枝径与角数之间均呈极显著相关。值得注意的是,冠幅与坐果数的相关系数最高,其次是与单株种子质量和单株果实质量间的相关系数,而冠幅与其他果实性状的相关系数均相对较低,说明冠幅主要是通过影响坐果数来影响单株产量的。共有23 个表型性状与单株种子质量之间均呈极显著相关。
2.3 凤丹栽培群体表型性状的聚类分析
基于不同单株各个表型性状的测定值,对26个表型性状进行系统聚类分析,结果如图2所示。图2表明,当平方欧氏距离为16 时,26 个表型性状可聚为6 类;其中,果实性状又可聚为2 类,角数单独聚为1 类(Ⅵ类),其余果实性状共同聚为1 类(Ⅰ类);当平方欧氏距离为15 时,可进一步将I 类性状聚为2 类,其中,坐果数、冠幅、单株果实质量和种子质量共同聚为1 个分支;除冠幅外的生长性状可聚为4 类:当年生枝长、当年生枝径和株高聚为1 类(Ⅱ类);复叶长、复叶宽、叶柄长和叶面积聚为1 类(Ⅲ类);顶小叶长宽聚为1 类(Ⅳ类);复叶数单独聚为1 类(Ⅴ类)。可见,果实性状间的相关性较生长性状间的相关性更强。
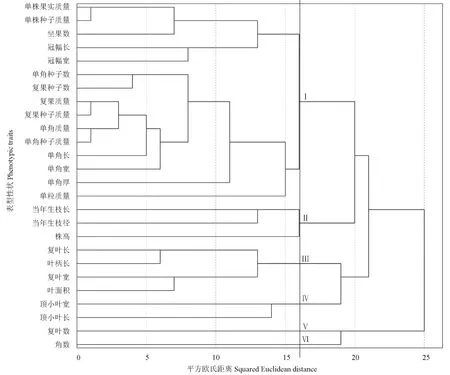
图2 凤丹栽培群体不同单株26 个表型性状的聚类图Fig.2 Clustering graph of 26 phenotypic traits based on different individual plants in P.ostii cultivated population
2.4 凤丹栽培群体单株产量的通径分析
以单株种子质量为因变量,以其他表型性状指标为自变量,进行多元线性回归分析,结果显示,当选择的自变量分别为单株果实质量(X1)、单粒质量(X2)、单角质量(X3)、单角种子质量(X4)、复果种子数(X5)和角数(X6)时,回归方程的决定系数(R2)最大,为0.981。以此6 个变量与单株种子质量(Y)构成的最优回归方程为:
Y=-4.92+0.432X1+50.84X2-14.158X3+19.773X4+0.777X5-2.329X6。
凤丹主要表型性状与单株种子质量的通径分析结果见表3。由表3可知,上述6 个自变量对单株种子质量的正向影响情况如下:单株果实质量的直接正向作用最大(其直接通径系数为1.006),而单粒质量的直接正向影响最小(其直接通径系数为0.078);单角质量的直接负向作用最大(其直接通径系数为-0.450)。6 个性状主要通过影响单株果实质量而影响单株种子质量,其中,单角质量、单角种子质量和复果种子数通过单株果实质量对单株种子质量产生的间接影响均较大,其间接通径系数分别为0.699、0.675 和0.642。6 个性状也都通过单角质量对单株种子质量产生一定程度的负向影响,其中,单角种子质量因此产生的间接负向影响程度最大(其间接通径系数为-0.425),其次分别为单株果实质量、复果种子数和单粒质量,其间接通径系数分别为-0.313、-0.311 和-0.292;单株果实质量还主要通过影响单角种子质量而对单株种子质量产生一定程度的间接影响(其间接通径系数为0.186)。
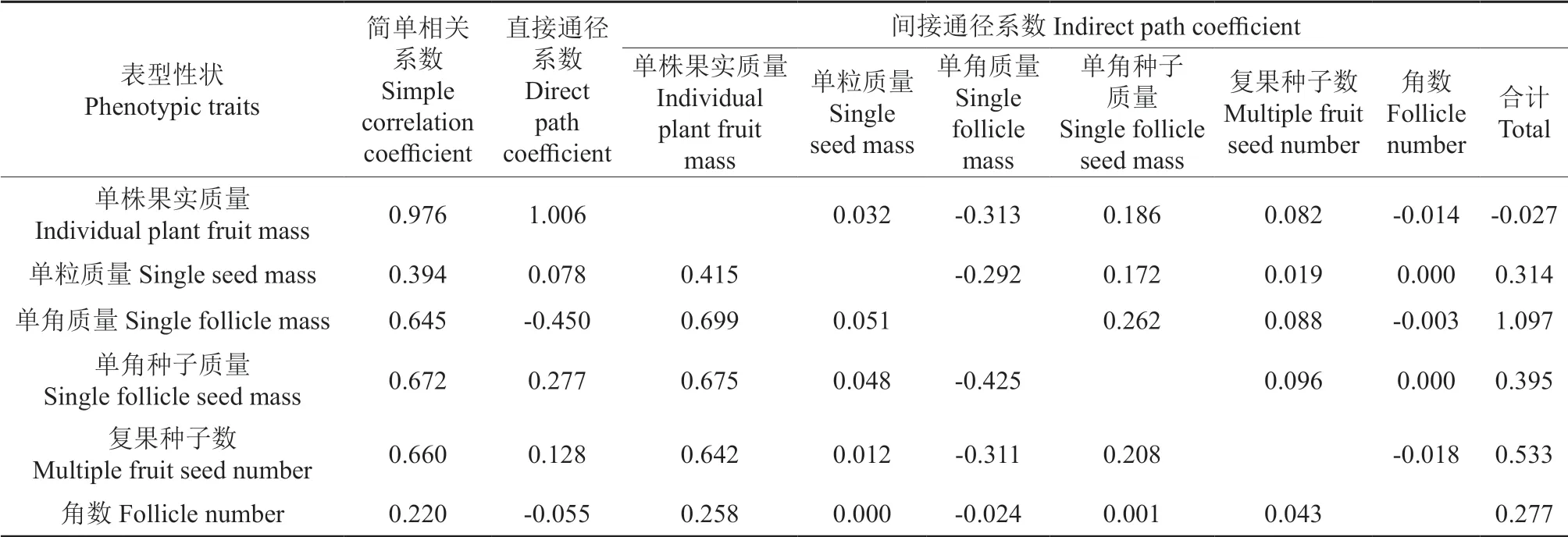
表3 凤丹栽培群体主要表型性状与单株种子质量的通径分析结果Table 3 Result of path analysis between main phenotypic traits and individual plant seed mass of P. ostii cultivated population
2.5 凤丹果实性状在不同角数下的变异情况
为了明确不同角数与果实的相对产量之间的关系,对凤丹不同角数下的果实性状测定值进行了方差分析,结果见表4。由表4可知,凤丹果实的复果质量、复果种子数和种子质量随着角数的增加整体均呈上升的变化趋势,7 个角的果实拥有最大的复果质量、复果种子数和种子质量,含8 个及以上角的果实复果质量、复果种子数和种子质量仍高于5 个角和6 个角果实。此外,单角种子数和种子质量随着角数的增加呈现先小幅度上升而后逐步下降的变化趋势;其中,6 个角果实的单角种子数和种子质量最大,5 个角果实的单角种子数和种子质量仍大于含7 个角以上的果实。

表4 角数不同的凤丹果实性状的方差分析结果†Table 4 Variance analysis result of fruit traits with different follicle number in P. ostii cultivated population
2.6 凤丹栽培群体表型性状的主成分分析
对凤丹栽培群体26 个表型性状进行主成分分析,提取出6 个主成分,其累计贡献率为76.619%,特征值总和为19.922,其中主成分1 的贡献率达到41.180%。26 个表型性状在6 个主成分中的载荷矩阵见表5。从表5中可以看出,主成分1(F1)的特征值为10.707,其不仅包括单角、复果和单株产量相关的全部性状,还涵盖了单粒质量、冠幅和当年生枝长(径)等指标的信息,因此,主成分1(F1)可称为果实因子或产量因子;主成分2(F2)的特征值为3.084,所代表的指标分别为复叶长、复叶宽、叶面积、顶小叶长和叶柄长,能够反映凤丹叶片形态,因此,主成分2(F2)可称为叶片因子;主成分4(F4)的代表指标包括复叶数和角数;主成分3、5、6 所代表的指标各有1 个,分别为坐果数、株高和顶小叶宽。

表5 凤丹栽培群体26 个表型性状的主成分载荷与贡献率Table 5 Factor loading matrix and contribution rate of 26 phenotypic traits in P. ostii cultivated population
2.7 凤丹栽培群体优良单株的筛选
利用主成分向量矩阵值除以相应主成分的特征值得到各个主成分的得分系数,再将其与各个表型性状测定值相乘即可得到每个样本在各主成分中的得分,最终根据各个主成分的贡献率能够获得各样本的综合得分,该得分可作为调查样本的综合评价指标值[13,26]。因而以不同主成分的贡献率构建的凤丹栽培群体不同单株的综合评价函数式为:
Y=0.411 8F1+0.118 6F2+0.083 1F3+0.064 0F4+0.048 1F5+0.040 6F6。
上式中:Y表示综合得分;F1、F2、F3、F4、F5、F6分别表示主成分1、主成分2、主成分3、主成分4、主成分5、主成分6 的得分。
239 株凤丹在前3 个主成分中的分布情况如图3所示。从图3中可以看出,239 株凤丹以原点为中心,均匀地分布在由前3 个主成分构成的坐标轴中。进一步利用综合评价函数,可以获得各单株的综合得分,其综合得分是各个单株26 个表型性状的综合体现。因此,将单株种子质量的平均值、标准差与主成分分析得到的综合评分相结合,并以此为依据,可以筛选出试验群体中产量高且表型性状优异的优良单株,凤丹栽培群体中不同等级植株的筛选结果见表6。以单株种子质量平均值加1 倍标准差作为筛选值时,有34 株入选;而以单株种子质量平均值加2 倍标准差作为筛选值时,则有9 株入选;若以单株种子质量平均值加3 倍标准差作为筛选值时,只有3 株入选。3 级植株的单株种子质量均值为143.88 g,与试验群体单株种子质量的平均值相比,其增量为104.18%;2 级植株的单株种子质量均值为191.78 g,与试验群体单株种子质量的平均值相比,其增量为172.15%;1 级植株的单株种子质量均值为240.55 g,与试验群体单株种子质量的平均值相比,其增量为241.35%:每个等级植株的单株种子质量均值较其上一等级植株的均值增加50 g 左右。进一步将入选的34 株凤丹与主成分分析得到的前34 名植株进行比较,结果约有50%的植株在以两个评价标准的评价结果中重合;而主成分1 主要体现试验群体的果实性状,因而将入选的34 株凤丹与在主成分1 中排名前34 名的植株进行比较,约有60%的重合率。这一比较结果说明,以单株种子质量的平均值加标准差为标准筛选得到的植株,一部分具有综合的优良性状,另一部分却相对缺乏代表性。有趣的是,以平均值加标准差为标准筛选得到的排名前12 的植株,全部包含在主成分1 得分排名前18 的植株内,因此,这12 株凤丹可以被评选为优良单株,而以2 级植株的简易评价标准来选择优良单株是合适的。

表6 凤丹栽培群体中优良单株的筛选结果†Table 6 Screening result of elite individual plant in P. ostii cultivated population

图3 凤丹栽培群体239 个单株在前3 个主成分中的分布情况Fig.3 Distribution of the first three principal components of 239 individual plants in P.ostii cultivated population
3 讨 论
本研究对凤丹栽培群体26 个表型性状进行测定及综合分析,结果发现:凤丹栽培群体的果实性状较其生长性状具有更大的变异系数,这与张晓骁等[16]和刘娜等[17]对紫斑牡丹的调查结果均一致;26 个表型性状多样性指数的平均值为1.97,这与孙珍珠等[25]对宽皮柑橘和张学超等[27]对苹果的分析结果均相似。而果实性状多样性指数较生长性状多样性指数更小,说明在同一栽植环境下,凤丹生长性状相对稳定,但果实性状因受环境、植株生长状况及授粉等方面的影响而表现出更大的变异,而果实性状中与果实质量相关性状的变异系数较其形态性状的变异系数更大,这与陈永忠等[28]对1 361 份油茶种质果实的调查结果一致。相关性分析结果表明,冠幅、坐果数及复果种子质量与单株种子质量的相关系数均较高,这一结果在聚类分析中得到了验证;然而,回归分析结果与相关性分析结果存在一定的差异,其主要原因是,回归分析中的单株果实质量囊括了上述部分与单株种子质量之间呈极显著相关的性状指标,而进一步通过逐步回归分析选择出更多的性状以建立最优的方程,以挖掘出更多的能影响单株种子质量的性状。通径分析结果表明,单角质量对单株种子质量体现出最大的直接负向影响,但直接相关系数却为较大的正值,这与其通过其他性状产生的间接正向影响较大有关,这一分析结果与杨玉霞等[11]在对金果榄主要农艺性状与产量的通径分析中得出的茎叶质量影响单株产量的结果类似。
单株种子质量是单株产量的最终体现,可作为评价凤丹单株质量的关键指标,在凤丹产业发展中占据重要地位,其受到环境、众多表型性状及其他因素的影响,然而,目前尚缺乏有关凤丹生长性状、果实性状与其单株产量间的相关性的研究报道。本研究结合相关性分析、聚类分析、回归分析及通径分析发现,单株果实质量、坐果数和冠幅对单株种子质量的影响均较显著,其次是复果种子质量、复果种子数、单角质量、单角种子质量及角数,这较崔虎亮等[18]对多个油用牡丹品种和刘娜等[17]对紫斑牡丹的研究结果增加了冠幅和角数2 个指标,而冠幅和角数对产量的影响分别在白鑫磊等[29]对文冠果和朱红莲等[30]对子莲的研究结果中有所证实。关于葡萄[31]和番茄[32]的相关研究结果表明,心皮数对果实产量存在显著影响,而角数是凤丹心皮数量在果实中的直接体现,其在不同单株间存在广泛的变异。因此,厘清角数与产量性状的关系对于凤丹优良种质的选育具有重要意义。史明伟等[33]对40 株不同角数的凤丹进行观察,结果发现,复果种子数随着角数的增加而增多,果质量和果径随着角数的增加也呈现出上升的变化趋势,且均在7 个角时达到最大值。本研究对凤丹栽培群体果实性状在不同角数下的测定值进行了方差分析,分析结果与史明伟等[33]的研究结果相似,并进一步确定,随着角数的增加,单角种子数和种子质量性状整体上呈下降趋势,而复果质量、复果种子数和种子质量整体呈上升趋势。根据所调查的性状与单株种子质量间的关系,本研究确定,在凤丹选育过程中,可将单株果实质量、坐果数和冠幅作为主要评选指标,而将复果种子质量、复果种子数、单角质量、单角种子质量及角数作为次要评选指标,以对所选凤丹单株进行综合评价。
筛选优良单株是目前获得优良基因型和品种选育的主要途径之一,通过对优良单株进行繁育,能够较快地获得改良的种质或品种。刘娜等[17]对419 株紫斑牡丹的12 个果实性状进行了调查与分析,确定了单株果实数、单株果实鲜质量、种子数和种子鲜质量为衡量紫斑牡丹单株产量潜力的主要性状指标,并以聚类中心 ± 标准差法制定了紫斑牡丹试验群体的产量分级标准,最终得到了8 株高产单株。本研究采用主成分分析法,提取出6 个主成分,其中主成分1 的贡献率占前6 个主成分贡献率总和的53.75%,并能够反映绝大多数果实性状的信息;结合单株种子质量和主成分1 的得分,筛选得到了12 株优良单株,同时确定以单株种子质量的平均值加2 倍标准差为优良单株的简易筛选标准,这与王晓丽等[34-35]对蓝桉超级苗的初步选择标准和优良单株早期选择标准均相似,但不同于刘娜等[17]的分级标准,本研究以单株种子质量为导向,提供一个更加简单直接的筛选标准,此标准可适用于优良单株的快速选择。然而,产量是否稳定及其他影响产量的因素在此筛选标准中也因此被忽略了,如相关性分析结果表明,冠幅能够影响坐果数进而影响单株产量,但冠幅大、单位面积产量低的植株占地面积却较大,而在相同的生长条件下冠幅较小、单位面积产量较高的植株占地面积小,然而其长势往往相对较弱。因此,后续研究可根据本研究提供的筛选标准,进一步结合多年的调查结果及冠幅等性状,以筛选出更加适宜的优良单株。
为了掌握凤丹栽培群体的表型变异规律,并筛选出凤丹选育过程中的关键评选指标,本研究以239 株凤丹为研究对象,对其26 个表型性状进行调查,整合多项分析结果,筛选得到的主要和次要评选指标可作为凤丹选育的参考依据。但是,在现有的研究背景下,本研究以同一地区的239 株单株为研究对象,旨在明确同一栽植环境下不同单株间的表型变异特征,研究结果对于该地区凤丹的选育具有一定的指导意义,但是,不同地区的环境和气候等因素差异显著,因而本研究结果也有待于在更多地区中予以验证。
4 结 论
本研究对凤丹栽培群体26 个表型性状进行调查与分析,结果发现,凤丹栽培群体不同单株间表型变异丰富,其果实性状的变异系数大于生长性状的变异系数;23 个性状与单株种子质量间均呈极显著相关,其中,冠幅、坐果数与单株果实和种子质量聚为1 个分支;回归分析结果表明,单株果实质量、单粒质量、单角质量、单角种子质量、复果种子数和角数对单株种子质量的影响均较显著,其中7 个角的果荚拥有更重的复果质量和复果种子质量;从调查的凤丹栽培群体中筛选得到12 株优良单株,并确定单株种子质量的平均值加2 倍标准差为优良单株的简易筛选标准。综合各项分析结果,在凤丹优良种质选育过程中,可以将单株种子质量、单株果实质量、坐果数和冠幅作为主要评选指标,而复果种子质量、复果种子数、单角质量、单角种子质量及角数为次要评选指标及应重点改良的性状。
猜你喜欢
特产研究(2022年6期)2023-01-17
中国农业科学(2022年13期)2022-07-26
植物资源与环境学报(2020年4期)2020-08-10
江苏农业科学(2020年21期)2020-03-08
天然产物研究与开发(2018年3期)2018-05-07
天然产物研究与开发(2018年3期)2018-05-07
现代园艺(2017年21期)2018-01-03
考试周刊(2016年11期)2016-03-17
中国康复理论与实践(2015年10期)2015-12-24
安徽农业科学(2015年25期)2015-12-22
