
一碳气体蛋白在水产饲料中的应用研究进展
2022-12-03 03:24■薛敏
饲料工业 2022年22期
■薛 敏
(中国农业科学院饲料研究所,北京 100081)
全球新冠肺炎疫情的影响及快速增长的人口对全球粮食供应带来前所未有的压力[1]。与此同时,大量温室气体排放导致的全球气候变暖,迫使127个国家或地区对“碳中和”和“碳达峰”做出承诺[2]。尽管畜牧业的快速发展可满足全球肉类的需求,但生产过程所排放的温室气体给“双碳”任务带来巨大挑战。与畜牧行业相比,水产养殖业更环保、更可持续,并可以提供更有营养的肉制品[3]。在过去的几十年里,水产养殖一直是全世界增长最快的食品生产板块之一。目前鱼粉仍是水产饲料中最理想的蛋白质来源,其适口性好,可消化率高,且含有未知的促生长因子。据统计,到2030年全球水产养殖行业将需要大约5 000万吨水产饲料,而鱼粉产量将长期稳定在500万吨左右,这无法满足水产养殖业的快速发展[4]。各类植物蛋白(如豆粕、棉粕和菜粕等)现已广泛作为鱼粉替代品应用于水产饲料中,这些植物蛋白具有相对较高含量的蛋白质,且市场价格相对较低。然而,植物蛋白的应用存在几个明显的局限性,如含有抗营养因子、适口性差和消化率低[5]。因此,开发低价、环保和可持续的新型饲料蛋白源对于满足水产养殖业的快速发展至关重要。
水产养殖副产品(鱼头、鱼皮、骨、鳞、鳍、鳔、血液和内脏等)、大型藻类(海带粉、马尾藻粉和石莼粉等)、昆虫粉(黄粉虫粉、黑水虻粉和蚯蚓粉等)以及单细胞蛋白(菌体蛋白、微藻、酵母等)被认为是最具潜力的鱼粉替代品[4,6-9]。其中单细胞蛋白,是指细菌、真菌或微藻在生长过程中利用各种底物(例如糖蜜、纤维和废气)产生的蛋白质。由于其蛋白质含量高、消化率高、氨基酸平衡性好,并且含有丰富的生物活性物质,市场潜力巨大[10-12]。在这些单细胞蛋白中,一碳(C1)气体菌体蛋白(C1GP)近年来受到了越来越多的关注。与传统农业不同,C1GP 是以C1 气体,如天然气(主要是甲烷)、甲醇、合成气[主要是一氧化碳(CO)和二氧化碳(CO2)]为底物经过某些特殊的细菌发酵生产的[13]。整个生产过程中不使用任何耕地和农药,生产更加高效和可持续,可有效地缓解人类粮食资源短缺的压力[14]。因此,在粮食短缺和“双碳”任务的双重需求下,C1GP开发具有巨大的前景。
目前生产C1GP 所用的细菌主要有甲烷氧化菌、甲醇氧化菌、乙醇梭菌和氢氧化菌(见表1)。其中,甲烷氧化菌,自1998年便开始小规模商业化生产,其产品Bioprotein®、FeedKind®和String Pro®已在水产养殖动物上进行了广泛试验[15]。一些利用甲醇气体为底物生产的菌体蛋白也已在水生动物上进行了相关评估试验(如PRUTEEN®和KnipBi 蛋白)[16-23]。利用含CO工业尾气生产的菌体蛋白,即乙醇梭菌蛋白(Rich-More®),近年来首次大规模商业化生产并被我国农业农村部批准的新型饲料蛋白源[24-25]。氢氧化细菌,是能够利用氢气作为电子供体,将CO2还原为有机物的一类细菌。能够利用分子态氢和氧之间的反应所产生的能量,并以碳酸作为唯一碳源而生长。遗憾的是,目前还没有关于其相关商业化产品的报道。C1GP产品的生产流程主要包括以下几个方面(见图1):工业或农业排放的C1气体经过净化和压缩后被收集并通过管道运输用于气体发酵;气体发酵系统可以根据细菌生长特性选择严格厌氧或好氧条件,发酵后所得的产品经离心、浓缩、喷雾干燥等一系列处理生产菌体蛋白;菌体蛋白在饲料企业中应用,并加工成商品饲料,用于鱼、虾养殖。
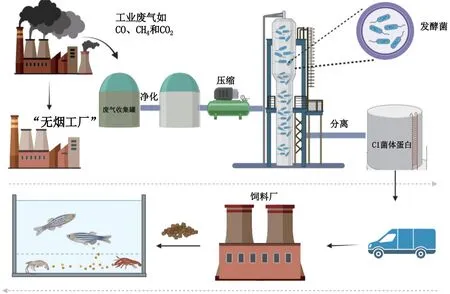
图1 C1GP产品的生产以及水产养殖应用

表1 C1GP产品的相关信息如产品名、产地公司(国家)、产品粗蛋白和粗脂肪含量
文章综述了以工业废气为底物发酵生产的C1GP产品,涉及目前的主要C1 气体来源、发酵所用的菌种、产品的营养组成,以及其对水产动物消化率、生长性能和生理功能的影响,旨在为水产养殖的可持续发展提供支持。
1 一碳气体及微生物同化途径
1.1 C1气体的介绍
全球C1气体排放量巨大,是温室气体排放的罪魁祸首。自工业革命以来,人类活动致使大气中的CO2浓度从1750 年 的280 mg/kg 增 加 到2021 的419 mg/kg[26]。甲烷(CH4)为另外一种温室气体,其全球年排放量为6~7亿吨。如今大气中的甲烷浓度已达到约1 875 μg/kg,是工业化前的两倍多。其中约40%的甲烷通过自然来源排放到大气中,剩余的60%来自人类活动(如反刍动物、农业活动、化石燃料开发、垃圾填埋场等)。合成气[以CO为主,包括CO、CO2和氢气(H2)的混合气体]是一种重要的化工中间体,全球年产量约为6 EJ,也是废弃物高温热解的主要产物之一[27]。甲醇(CH3OH)在室温下是液态的,但作为化学工业合成气的一部分,它会与高温合成气一起以气体的形式排放。这些C1气体的直接排放或燃烧会造成严重的环境污染和资源浪费。因此,通过工业生物技术将这些气体转化为饲料蛋白质等产品是首选的一种技术手段。
1.2 微生物发酵C1气体过程中的同化作用
固碳作用是微生物发酵C1气体最重要的生物反应,不同细菌其固碳的途径有所不同。氢氧化细菌,又称Knallgas细菌,是一种利用氢气作为电子供体,将CO2还原为有机物的一类细菌。例如,在自养条件下,Ralstonia eutropha细菌可以通过卡尔文-本森-巴沙姆循环(Calvin Benson Bassham,CBB)固定CO2。尽管CBB 循环的固碳效率不如其他方法[28-29],但Ralstonia eutropha是第一个被认为可以生产C1GP 的细菌。甲烷氧化菌和厌氧型甲烷菌[30],可用于甲烷发酵的微生物。它们通过核酮糖单磷酸途径(RuMP)、丝氨酸循环或CBB 循环实现对甲烷的利用。其中依赖氧气的甲烷氧化菌在甲烷合成甲醇的过程中需要氧气的活化[31-33]。在厌氧条件下生长的甲烷菌,如报道的生活在深水中的古生菌Archaea anme,主要是借助辅酶,依靠与硫酸盐还原物种的共生关系完成甲烷的同化[31-32,34]。乙醇梭菌和李氏梭菌(Clostridium autoethanogenum和Clostridium ljundahl)等严格厌氧菌是通过还原乙酰辅酶A 途径(又称Wood-Ljungdahl 途径)将合成气转化为乙酰辅酶A,然后再合成蛋白质等化学物质,此过程生产效率最高[35-36]。在同化过程中,CO作为碳单位,H2参与还原当量和能量的产生[37]。如果细胞内代谢需要额外的能量,CO 可以在甲基支链
上被氧化成CO2,这比从氢中产生能量成本更低,效率更高[38]。甲醇营养微生物是一类可以利用甲醇作为底物生成C1GP 的微生物,分为甲醇营养细菌和酵母。大多数时候,它们通过RuMP、木酮糖单磷酸途径(XuMP)和丝氨酸循环途径来同化甲醇。
氮代谢被认为是生命有机体中仅次于光合作用的第二重要的生物反应[39],并且固氮菌广泛存在于各种环境中[40],尤其是对于那些生活在严重贫瘠环境中的微生物来说,氮代谢能力是不可或缺的[41]。已有大量证据证实氮代谢广泛存在于固氮菌中[42-43],例如梭菌(Clostridiumsp.)和甲烷氧化菌(Methylomonassp.)[44],它们都含有各类固氮酶,可将无机氮转化为有机氮[45]。在氮代谢过程中,氮气(N2)、亚硝酸根(NO2-)、硝酸根(NO3-)、一氧化二氮(N2O)和氨气(NH3)可通过固氮酶作用转化为NH4+,随后分别通过丙氨酸或谷氨酸生成C1GP。此外,文章提到的梭菌和甲烷氧化菌已经被证实具有从大气中固氮的性质,并且可利用大气中的氮气作为其生长的唯一氮源[44,46]。
2 C1GP在水产饲料中的应用
2.1 C1GP的营养成分
C1GP的营养成分受到菌种、底物和发酵条件,以及发酵后续加工等因素的影响。总的来说,C1GP 产品具有较高含量的粗蛋白和均衡的氨基酸组成,并富含维生素、矿物质、磷脂和其他功能性化合物[12]。如表1 所示,一些C1GP 产品的粗蛋白含量高于或与鱼粉相当[16,47]。此外,还发现同一产品不同批次的C1GP,其营养成分也会发生变化。这主要归因于发酵所用气体含量和发酵条件的变化。必需氨基酸方面,与优质鱼粉相比,甲烷菌蛋白的赖氨酸和蛋氨酸含量相对较低,而乙醇梭菌蛋白的精氨酸和色氨酸含量稍低。同时也发现,乙醇梭菌蛋白和甲醇菌蛋白都富含赖氨酸、蛋氨酸和支链氨基酸[48]。C1GP 产品的粗脂肪含量通常较低且变化较大,例如乙醇梭菌蛋白粗脂肪的含量为0.16%~2.00%,甲醇菌蛋白为0.84%~1.29%,而甲烷菌蛋白的粗脂肪含量则高于7.00%(见表1)。并且它们的脂肪酸组成也不同,如以甲醇为底物发酵生产的菌体蛋白其主要脂肪酸是十六烷酸和顺-9-十六烷酸,而以甲烷为底物则是磷脂、16∶0和16∶1 脂肪酸[48]。C1GP 产品的总能量与鱼粉相当。粗灰分方面,发酵甲醇生产的菌体蛋白中的灰分含量略高,如甲醇菌蛋白中的灰分含量超过8.5%,而某些甲烷菌蛋白的粗灰分含量较低,为6.3%~8.0%,乙醇梭菌蛋白粗灰分含量最低,为3.3%~6.4%[48]。
2.2 C1GP的消化率
消化率是饲料蛋白原料一个重要营养特征,不同C1GP 产品在不同养殖动物间的消化率差异巨大。Øverland等[11]已综述了甲烷和甲醇菌蛋白的营养物质的消化率。甲烷菌蛋白的氨基酸的消化率因氨基酸类型不同而变化。例如,大西洋鲑(Salmo salar)对各类氨基酸的消化率为51.9%~91.8%,其中赖氨酸(91.7%)和精氨酸(91.8%)的消化率最高,但半胱氨酸(51.9%)的消化率较低[15]。此外,氨基酸的消化率也很大程度上取决于生产过程中选用的菌株和底物。Skrede等[49]比较了以甲烷和甲醇为底物生产的菌体蛋白在水貂体内的消化率,发现甲醇菌蛋白的氨基酸表观消化率要远高于甲烷菌蛋白。不同水产养殖动物对甲烷菌蛋白中的营养物质表现出不同的表观消化率。例如,大西洋鲑对Bioprotein®中粗脂肪和粗蛋白的消化率分别为87.2%[11]和81.9%[15],而南美白对虾(Litopenaeus vannamei)对FeedKind®中粗脂肪和粗蛋白的消化率为98.9%和90.3%。甲醇菌蛋白中营养物质的表观消化率也因养殖物种不同而存在较大的差异。关于乙醇梭菌蛋白,南美白对虾对其粗脂肪、粗蛋白、磷和大多数必需氨基酸的消化率分别为93.9%、98.9%、63.2%和90.0%以上,而杂交石斑鱼(Epinephelus fuscoguttatus♀×E.lanceolatus♂)对其粗脂肪和粗蛋白的消化率则分别为85.5%和82.2%[48]。
一般来讲,C1GP 产品应用于鱼类饲料其营养物质的消化率与鱼粉相当。例如,FeedKind®可以替代日本鰤鱼(Seriola quinqueradiata)中高达30%的鱼粉,而不影响饲料营养成分的消化率[50]。乙醇梭菌蛋白可以有效替代70%以上的鱼粉,而不会影响大口黑鲈(Micropterus salmoides)日粮中粗蛋白的消化率[51]。然而,也有研究发现随着大西洋鲑和虹鳟鱼(Oncorhynchus mykiss)摄食Bioprotein®比例的增加,包括蛋白质、氨基酸、脂肪、能量和铜在内的营养物质消化率显著下降[48]。不同研究结果之间的差异部分归因于不同养殖动物利用C1GP 产品的特定能力的不同。此外,当这些C1GP产品添加到水产动物饲料中时,由于发酵条件和后续加工条件的差异,产品之间的营养成分的消化率也会有所不同,甚至同一产品的不同批次也会发生变化[50]。因此,C1GP产品在发酵后进行后续加工时,应尽可能保证较高的营养成分和消化率[52]。消化酶在提高营养物质消化率方面也起着重要作用。研究发现,乙醇梭菌蛋白替代黑鲷(Acanthopagrus schlegelii)幼鱼饲料中58.2%鱼粉对其肠道中的蛋白酶、脂肪酶和淀粉酶活性没有显著影响,尽管胃淀粉酶活性随着乙醇梭菌蛋白添加量的增加而逐渐降低[47,53]。与之相反,当饲料中鱼粉的替代量达到39%时,幼鱼的肠道脂肪酶和淀粉酶活性会显著增加,同时干物质和蛋白质的表观消化率也会增加,并且替代比例在75%时,胃和肠道中的蛋白酶活性最高[53]。此外,有研究表明饲料中添加乙醇梭菌蛋白还能提高尼罗罗非鱼(Oreochromis niloticus)营养吸收相关酶的活性[54]。这些结果的出现可能归因于乙醇梭菌蛋白发酵过程中会产生一定含量的乙酸等功能成分,这些成分可以调节肠道微生物,改善肠道健康,从而增强鱼类的消化和吸收功能[55]。
2.3 C1GP对水产动物生长性能的影响
甲烷菌蛋白作为最早的C1GP产品之一,作为鱼粉替代品已广泛应用于各种水产养殖动物,包括大西洋鲑、庸鲽(Hippoglossus hippoglossus)、虹鳟鱼、日本鰤鱼、尼罗罗非鱼、黑鲷、大口黑鲈和南美白对虾[48]。对大西洋鲑的研究发现[56-58],饲喂添加300 g/kg Bioprotein®的饲料不会对其生长性能产生负面影响,但添加至370 g/kg 时,大西洋鲑的生长性能和存活率均受到显著抑制。然而,另一项研究发现,随着饲料中Bioprotein®含量(0、100 g/kg 和200 g/kg)的增加,大西洋鲑的生长性能略有下降[59]。这些研究结果的差异部分归因于不同的实验条件、饲料配方或鱼体大小。除替代水平外,不同食性鱼类利用甲烷菌蛋白能力也有明显差异。FeedKind®可以成功替代日本鰤鱼[50]和大口黑鲈[60]饲料中高达30%的鱼粉,或尼罗罗非鱼[61]饲料中全部鱼粉,而不会对生长性能和健康产生负面影响。然而,另一篇研究发现,饲料中添加18% Bioprotein®会显著降低庸鲽的生长性能[62]。FeedKind®在南美白对虾上也进行了大量实验,其中一项研究发现FeedKind®可以替代饲料配方中100%鱼粉而对虾的生长性能、存活率和饲料利用率没有负面影响[63]。甲醇菌蛋白,如PRUTEEN®,最早在悉尼岩牡蛎(Saccostrea commercialis)上进行了相关试验[18-20]。这些研究表明,171 g/kg是其最佳添加水平,与其他单细胞蛋白(如酵母)相比,饲料中添加235 g/kg PRUTEEN®可显著提高牡蛎糖原含量和肥满度。KnipBio是另外一种甲醇菌蛋白,其被认为是鲑科鱼类饲料中一种高度可消化的新型蛋白原料[23]。KnipBio 蛋白可有效替代石鲈(Haemulon chrysargyreum)饲料中30%、大西洋鲑中55%,以及南美白对虾中50%鱼粉而对机体生长性能和饲料表观消化率无负面影响[64]。虹鳟上最新的研究发现,10% KnipBio 饲料添加会略微降低其生长性能,研究者认为生长性能的下降归因于采食量的降低,这可以通过在饲料中添加诱食剂来解决[65]。两项利用实验室规模生产的甲醇菌蛋白作为豆粕替代在南美白对虾上的研究表明,这种菌体蛋白的消化率低于豆粕[21]。此外,饲料中添加40 g/kg的甲醇菌蛋白对对虾的生长性能没有显著影响,而较高的添加水平(>60 g/kg)则抑制了对虾生长性能、饲料转化率和氨基酸保留效率[22]。关于CO 菌体蛋白,在2018 年发表了第一篇利用乙醇梭菌蛋白替代草鱼(Ctenopharyngodon idllus)饲料中豆粕的研究[25]。随后,大量研究表明,乙醇梭菌蛋白是一种安全、优质的鱼粉或豆粕的替代品,且部分替代对养殖动物的生长性能和健康没有任何不利影响[48]。值得注意的是,南美白对虾饲料中添加高比例的乙醇梭菌蛋白,即替代45%以上的鱼粉,会对生长和免疫功能产生不利影响[66-67]。生长性能和健康状态下降的原因可归结为两个方面。一方面,与鱼粉相比,乙醇梭菌蛋白中相对地缺乏一系列功能性物质,如必需氨基酸、必需脂肪酸、生长因子、牛磺酸等;另一方面,根据肝胰腺转录组分析的结果,饲料中高水平的乙醇梭菌蛋白会对蛋白质合成和营养利用产生负面影响,并扰乱免疫反应和代谢过程,从而降低生长性能[48]。
2.4 C1GP对水产动物生理功能的影响
饲料中添加一定比例的C1GP对水产养殖动物的健康会起到积极影响。在大西洋鲑的研究中发现饲料中添加Bioprotein®可以有效预防大豆诱导的肠炎[56,58,68],并且肠炎现象会随着饲料中菌体蛋白添加水平增加而逐渐消失。此外,不同加工工序处理的甲烷菌蛋白对大豆诱导的肠炎都有显著的缓解作用[68],并且这种保护作用具有一定的剂量依赖性[56]。然而,乙醇梭菌蛋白对鱼类健康的影响在不同养殖种类之间并不一致。例如,利用100 g/kg乙醇梭菌蛋白替代草鱼饲料中55%豆粕可以改善其肠道形态,进而改善生长性能[24],而这个添加比例对尼罗罗非鱼的肠道造成负面影响[54]。一项对建鲤的研究表明,饲料中添加200 g/kg的乙醇梭菌蛋白对其中肠的组织结构没有显著影响[69]。在水产配合饲料中添加一定水平的C1GP有利于促进养殖动物的生长性能并提高抗病能力,这部分归因于饲料中菌体蛋白的若干免疫刺激和抗氧化调节特性,因为它们含有丰富的生物活性成分,这些成分可以提高免疫相关基因的转录水平,如雷帕霉素(TOR)、补体3(C3)和干扰素-γ(IFN-γ),提高血浆免疫球蛋白M和白蛋白水平,调节抗氧化酶和相关基因,以及调节肠道微生物组成[48]。另一方面,气体菌体蛋白饲料可以改善鱼类的生长性能和健康状态,这可能归因于前面所讨论的高消化率和代谢调节功能。在代谢调节方面,饲料中添加乙醇梭菌蛋白可以调节糖脂代谢,通过AMPK信号通路维持尼罗罗非鱼机体的能量平衡[70]。对对虾代谢组学的研究发现,饲料中添加甲烷菌蛋白对对虾的三羧酸循环(TCA)循环、抗坏血酸和醛酸代谢以及谷胱甘肽代谢有显著影响[48]。Chen等[71]还发现,饲料中添加甲烷菌蛋白可以降低肝胰腺中MDA 含量和提高抗氧化酶活性,增加肠道黏膜褶皱高度,调整肠道微生物组成,增强对副溶血性弧菌(Vibrio parahaemolyticus)感染南美白对虾的抵抗力。此外,C1GP中的核苷酸水平远远高于鱼粉和豆粕,核苷酸被认为是鱼类的半必需营养素,饲料中一定水平的核苷酸可以促进一系列的免疫反应[48]。
3 展望
C1GP 已被证明是一种安全有效的蛋白原料,对水产养殖动物的生长性能和健康状态都起到积极作用。C1GP 的技术创制将有助于水产养殖业绿色、可持续发展。但是C1GP产品在水产饲料中的广泛应用仍存在一些挑战。首先是潜在的生物和化学有害成分,尽管到目前为止没有任何证据证明这些由工业废气转化而成的蛋白质存在有害物质。但大家的担忧与疑虑不能忽视,因此政策引导和正面宣传很有必要。另一个挑战是目前产量还不高,尽管在过去几年中,一些公司通过开发新技术、修饰菌株以及优化发酵过程在大规模生产方面取得了较大进展,但相比于我国对饲料蛋白质的需求而言,依然是杯水车薪。值得庆幸的是C1GP 作为优质的功能性蛋白,可与其他蛋白原料混合使用,以满足养殖动物的营养需求,以及应对常规原料价格的波动。总体而言,随着合成生物学技术的快速发展,构建转化效率高、适应性广、营养丰富的新型菌种和气体发酵技术将成为未来水产养殖新原料开发的重要领域。
猜你喜欢
当代水产(2022年8期)2022-09-20
当代水产(2022年6期)2022-06-29
当代水产(2022年1期)2022-04-26
当代水产(2022年2期)2022-04-26
中国饲料(2021年17期)2021-11-02
当代水产(2021年4期)2021-07-20
猪业科学(2021年3期)2021-05-21
中国畜牧杂志(2021年5期)2021-05-17
当代水产(2021年2期)2021-03-29
国际种业前沿动态(2020年16期)2020-10-14
