
不同赖氨酸水平对熊蜂幼年工蜂发育及代谢影响
2022-12-03 03:10郝冬梅樊莹徐进郭岳琴李继莲姚军
中国农业科技导报 2022年10期
郝冬梅,樊莹,徐进,郭岳琴,李继莲,姚军*
(1.中国农业科学院蜜蜂研究所,北京 100093;2.贵州省畜禽遗传资源管理站,贵阳 550001)
地熊蜂(Bombus terrestris)属于蜜蜂总科(Apidae)熊蜂属(Bombus),其不仅采集能力强[1],而且环境适应能力强,具有耐热、耐湿、耐寒、可规模化饲养和易运输等特点[2-3],已大规模的应用于农业授粉中。地熊蜂的生长发育有卵、幼虫、蛹、成蜂4 个阶段,其中,幼虫和成蜂阶段是营养摄入的主要阶段,此阶段提供优质、充足营养的饲料与熊蜂规模化饲养和后期授粉应用关系密切。在为设施作物授粉期间,单一作物的花粉很难提供熊蜂生长发育所需全部营养,适当的营养强化则有助于延长熊蜂的授粉周期。在熊蜂工厂化繁育中,尽管目前人工饲养技术已逐渐成熟[4],但为了饲养更强健、授粉周期更长的蜂群,都会对饲料配方进行不断优化。蛋白质是熊蜂所必需的营养,对成年蜜蜂的免疫应答、寿命有积极影响[5-6]。氨基酸是大多数生物合成蛋白质的基础,对蜜蜂生长、发育起关键作用,影响营养需求的平衡,成年蜂的飞行能力、健康和寿命也与其有关,最终将影响授粉效果。因此了解成年熊蜂的氨基酸需求对熊蜂规模化饲养和授粉应用都十分重要。
除了参与蛋白质合成外,氨基酸在调节基因表达、细胞信号、抗氧化反应和免疫反应方面发挥着重要作用[7]。食物中补充谷氨酰胺或精氨酸可增加小肠与脂肪组织中抗氧化基因的表达,降低促炎基因的表达;谷氨酸与天冬氨酸调节还原当量物通过线粒体膜的转移,从而调节糖酵解和细胞氧化还原状态;色氨酸通过吲哚胺2,3-双加氧酶(indoleamine 2,3-dioxygenase,IDO)的分解代谢对巨噬细胞和淋巴细胞的免疫功能有着重要作用。
氨基酸在营养学中分为必需氨基酸和非必需氨基酸,其中,生物体内无法自行合成必需氨基酸,必需通过食物获得,而蛋白质种类决定昆虫消化产生必需和非必需氨基酸的比例和类型[8-9],单一种类花粉通常不能满足熊蜂发育所需的氨基酸。与非必需氨基酸相比,饲喂含有必需氨基酸饲料可增加哺育蜂头部与觅食蜂胸部重量,通过头部与胸部质量可反映蜜蜂腺体与肌肉发育情况[10],并且蜜蜂对必需氨基酸的偏好高于非必需氨基酸[11]。研究发现,向日葵花粉中蛋氨酸与色氨酸含量低,导致使用向日葵花粉饲喂的熊蜂蜂群幼虫重量显著低于其他花粉组[5,12],饲喂单一向日葵花粉会降低工蜂寿命,抑制新出房工蜂咽下腺发育[13]。Groot[14]通过笼养试验确定了蜜蜂所需的10 种必需氨基酸。熊蜂对必需氨基酸的需求与蜜蜂相似,但熊蜂对氨基酸的需求量鲜有报道,因此有必要进一步深入研究不同氨基酸对熊蜂发育的影响。
自然界中,熊蜂一般采集蛋白质丰富、氨基酸含量高的混合花粉[15],氨基酸含量高的花粉可提高熊蜂产卵率与幼虫质量[16]。如果饲料中缺乏某种必需氨基酸或氨基酸含量低都会抑制熊蜂蜂群的发育。熊蜂饲粮中,必需氨基酸水平过高会导致体内增加过多氮排泄物,从而缩短寿命[17]。
赖氨酸是蜜蜂生长发育必需的,在动物体内参与酶、血清蛋白、激素等蛋白质的合成,对脂肪代谢、免疫功能方面有着重要作用[18-19]。研究发现,饲粮中不同赖氨酸水平对肉鸡日增重和仔鸡血清中白蛋白、总胆固醇尿素氮有着显著影响[20]。Chen 等[21]研究发现,赖氨酸缺乏导致肉鸡体内对新城病毒抗体水平下降。此外,饲粮中添加赖氨酸可显著提高单胃型肉用动物的肌肉蛋白质含量与大菱鲆的粗蛋白含量[22-23]。当猪饲料中赖氨酸水平降低时,肌内脂肪含量增加,腰眼面积减少[24]。对于蜜蜂,赖氨酸、色氨酸等氨基酸在蜜蜂生长中起关键作用,并影响营养需求的平衡[25];Groot[26]研究表明西方蜜蜂对赖氨酸的适宜需求量为3%;王帅等[27]推荐意大利蜜蜂幼虫对赖氨酸需求量为11.08~16.08 mg·g-1。对于畜禽养殖,赖氨酸是谷类蛋白质的第一限制氨基酸,而对于熊蜂全价饲料或氨基酸营养补充剂的研发,谷物是最易于获取的原料来源。因此,本研究首选了赖氨酸作为研究重点。
代谢组学是研究生物体系代谢途径的学科,是系统生物学的重要组成部分。基于代谢组学分析熊蜂血淋巴的生物标志性成分有助于改善熊蜂健康监测和鉴别营养压力来源[28]。本研究拟通过饲喂熊蜂含有一定水平赖氨酸的饲粮,通过液相色谱∕质谱联用技术(liquid chromatograph mass spectrometer,LC-MS)检测其对熊蜂工蜂血淋巴代谢的影响,同时采用主成分分析(principal component analysis,PCA)和正交偏最小二乘法-判别分 析(orthogonal partial leastsquaresdiscriminant analysis,OPLS-DA),筛选对熊蜂工蜂血淋巴代谢影响的差异代谢物,并结合代谢通路数据库(kyoto encyclopedia of genes and genomes,KEGG)探讨涉及赖氨酸代谢的通路,有助于了解赖氨酸对熊蜂的生理功能,发掘对熊蜂生长、发育和寿命有利的因素。
1 材料与方法
1.1 供试蜂群
供试熊蜂由中国农业科学院蜜蜂研究所传粉蜂生物学与授粉饲养室提供。选择群势相当的健康蜂群,共选取体质量相近的1 日龄工蜂140 只,置于同一蜂箱中,随后从中随机选取5 只工蜂组成无王工蜂群,共28 个。将28 个无王蜂群分为4 组,分别饲喂4 种不同赖氨酸含量的糖水,每组7个重复,温度为(26±2)℃,相对湿度为60%±5%,黑暗条件下进行饲养。
1.2 饲粮配置
供试饲粮为60%(质量体积分数)蔗糖水溶液,含18种氨基酸(赖氨酸、精氨酸、组氨酸、苯丙氨酸、甲硫氨酸、色氨酸、亮氨酸、异亮氨酸、苏氨酸、缬氨酸、甘氨酸、丝氨酸、丙氨酸、胱氨酸、酪氨酸、羟脯氨酸、谷氨酸、天冬氨酸),总氨基酸添加量为蔗糖(干质量)的3%。除赖氨酸外,各种氨基酸添加量参照酪蛋白中氨基酸组成添加。所用氨基酸均为L-型氨基酸,购自生工生物工程(上海)股份有限公司。赖氨酸处理设置4 个水平,添加量分别占总氨基酸的0%(对照)、2%、4%、6%。
1.3 饲喂管理
4 组工蜂群分别饲喂4 种不同含量赖氨酸的饲粮,9:00—10:00 饲喂,饲喂结束后将瓶中剩余的蔗糖与氨基酸混合液清除掉,并清洗饲喂瓶,每隔3 d 饲喂1 次,注入新熬制的蔗糖与氨基酸混合液。每天观察微蜂群情况,及时将死亡的工蜂移除,至20 日龄时终止饲喂,完成试验蜂群的整个饲养阶段。
1.4 测定指标及方法
1.4.1 工蜂存活率 在试验期间,每天观察每组工蜂死亡情况,并记录死亡数量。
1.4.2 工蜂肌肉蛋白含量 试验结束后每个处理随机选取工蜂,称重后取出胸部肌肉,加入0.9%的生理盐水,使用微型涡旋震荡器(QL-901,江苏南海市其林贝尔仪器制造有限公司)匀浆,2 500 r·min-1离心10 min后取上清液。采用总蛋白定量测试盒(A045-2-2,南京建成生物工程研究所)测定胸部蛋白含量,每组处理5个重复。
1.5 血淋巴代谢物提取和检测
1.5.1 样品采集 无王蜂群饲养至15 日龄时,随机选取工蜂10 只,选取头部一端触角,使用70%乙醇对触角周围的区域消毒,随后用镊子去除该触角,轻轻按压工蜂腹部至工蜂触角部位出现血淋巴,立即用移液枪将其吸出[29],收集于1.5 mL离心管,保存于-80 ℃冰箱备用。
1.5.2 代谢物提取和检测 利用100%甲醇提取血淋巴中的代谢物,每个处理组6 个平行。将冻存的样品在冰上充分解冻,取10µL 淋巴样品,加入50µL甲醇,超声10 min,使样品充分溶解,4 ℃、12 000 r·min-1离心20 min,取上清液于内衬管中,置于2 mL棕色进样瓶中供LC-MS分析。
超高效液相色谱系统(Ultimate 3000,美国Thermo Fisher Scientific 公司)检测条件:ACQUITY BEH Amide 色谱柱(Waters,150 mm×2.1 mm,1.7 µm),进样量2 µL,柱温50 ℃;流动相A 为30%乙腈溶液(含0.1%甲酸和10 mmol 甲酸铵),流动相B 为95% 乙腈溶液(含0.1% 甲酸和10 mmol 甲酸铵),流速0.3 mL·min-1。梯度洗脱:0.0~1.0 min,100% B;1.1~11.0 min,100%~30%B;11.1~11.5 min,30%~100%B;11.6~15.0 min,100%B。
质谱仪(Q-Exactive,美国Thermo FisherScientific公司)采用HESI离子源,在正、负离子切换模式下采集。参数设置:离子源温度320 ℃,喷雾电压正离子3.5 kV 和负离子3.0 kV,鞘气体流速40 arb,辅助气流速5 arb,S-lens 射频电压60%,母离子扫描分辨率70 000,扫描范围70~1 000 m·z-1,自动增益控制目标离子数1×106,最大离子注入时间50 ms,扫描速率1 scan·s-1。
1.6 数据分析
1.6.1 蛋白质含量统计分析 工蜂胸部蛋白含量利用SPSS 20.0统计软件进行单因素方差分析,组间比较采用最小显著性差异法(LSD 法);生存曲线采用Logrank 检验分析,统计结果以P<0.05 具有统计学差异,使用Graphpad Prism 8软件进行图表的绘制。
1.6.2 代谢组学数据分析 将原始代谢组学数据Raw 文件导 入Compound Discoverer 2.1软件(Thermo Fisher Scientfic)进行离子峰识别、峰对齐、峰面积归一化处理及数据库搜索,使用KEGG(http:∕∕www.genme.jp∕kegg∕)和HMDB(http:∕∕www.hmdb.ca∕)数据库进行化合物鉴定。随后采用MetaboAnalyst 5.0 进行统计学和富集分析:通过主成分分析反映处理组间的样本总体代谢差异;正交偏最小二乘法-判别分析(orthogonal partial leastsquares-discriminant analysis,OPLS-DA)反映组内样本间的变异度大小;对OPLS-DA 图验证模型是否稳定和出现过拟合现象,其中R2X和R2Y分别表示所建模型对X和Y矩阵的解释率,Q2表示模型的预测能力,Q2>0.5 表示模型具有较好的判别分析能力,Q2与R2越接近说明模型越稳定且无过拟合现象。根据变量重要性投影(variable importance in the projection,VIP)值>1、P值<0.05、FC 值>1.5或FC 值<1∕1.5筛选差异代谢物,最后进行富集分析。
2 结果与分析
2.1 饲粮赖氨酸水平对工蜂胸部肌肉蛋白含量的影响
由图1 可知,随着饲粮赖氨酸水平的增加,20 日龄工蜂胸部肌肉蛋白含量呈上升趋势。与饲粮无赖氨酸对照(0%)相比,饲粮赖氨酸含量4%时工蜂胸肌肉部蛋白含量显著升高。当赖氨酸水平提高至6%时,胸部肌肉蛋白含量显著高于2%和0%处理组(P<0.05)。
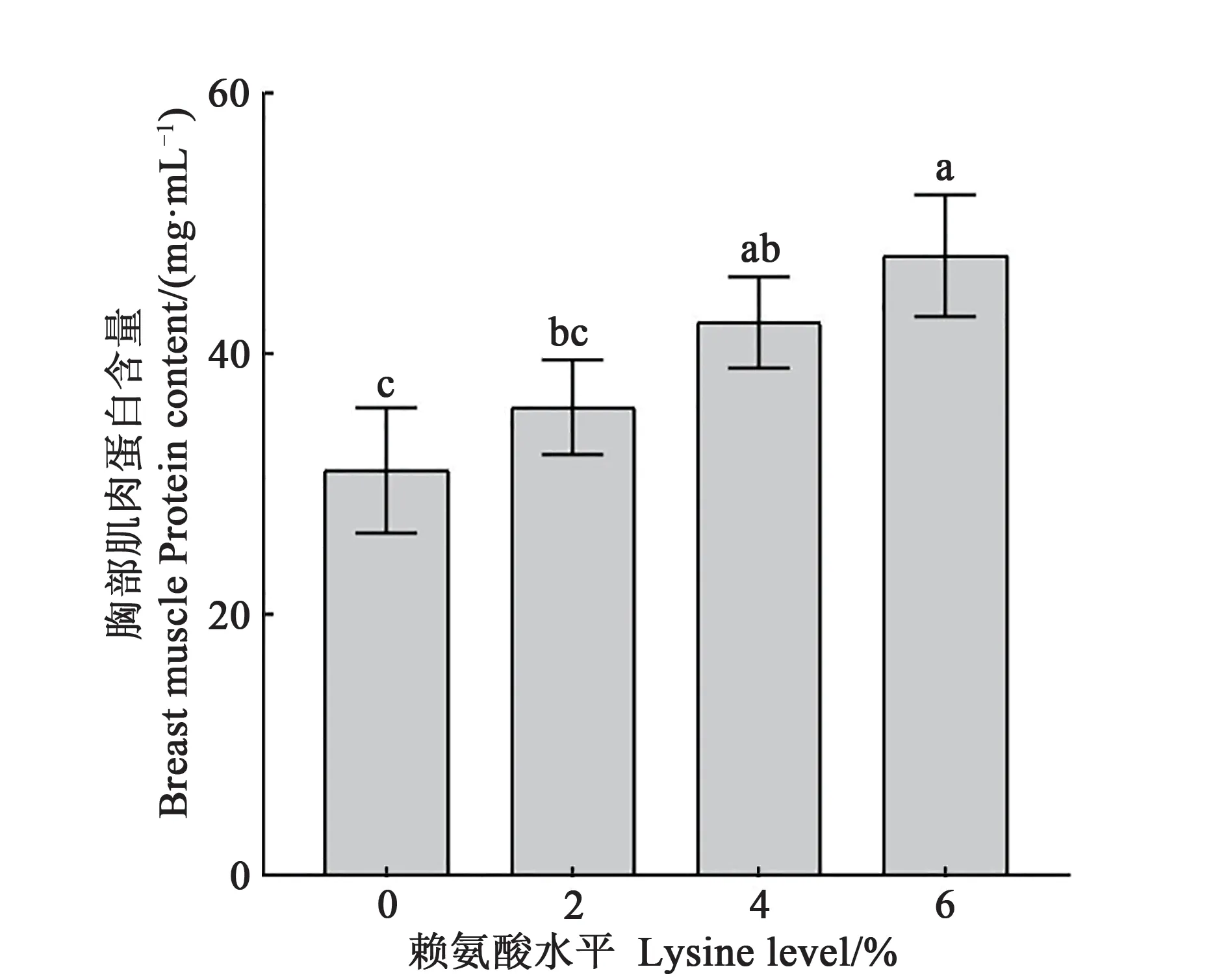
图1 不同处理下工蜂胸部肌肉蛋白含量Fig.1 Chest muscle protein content of worker bees under different treatment
2.2 饲粮赖氨酸对工蜂死亡数量的影响
在不同的赖氨酸添加量饲喂中,熊蜂的存活率不同(χ2=21.70,P<0.001)。由图2和表1可以看出,当饲料中缺乏赖氨酸(0%)时,工蜂存活率最低,赖氨酸水平为6%时,工蜂存活率最高,且两处理间差异极显著(P<0.001);赖氨酸水平为2%与4%时,工蜂存活率相近,组间差异不显著。

表1 不同赖氨酸水平下工蜂存活率的差异Table 1 Differences in survival rate of workerbees under different lysine levels
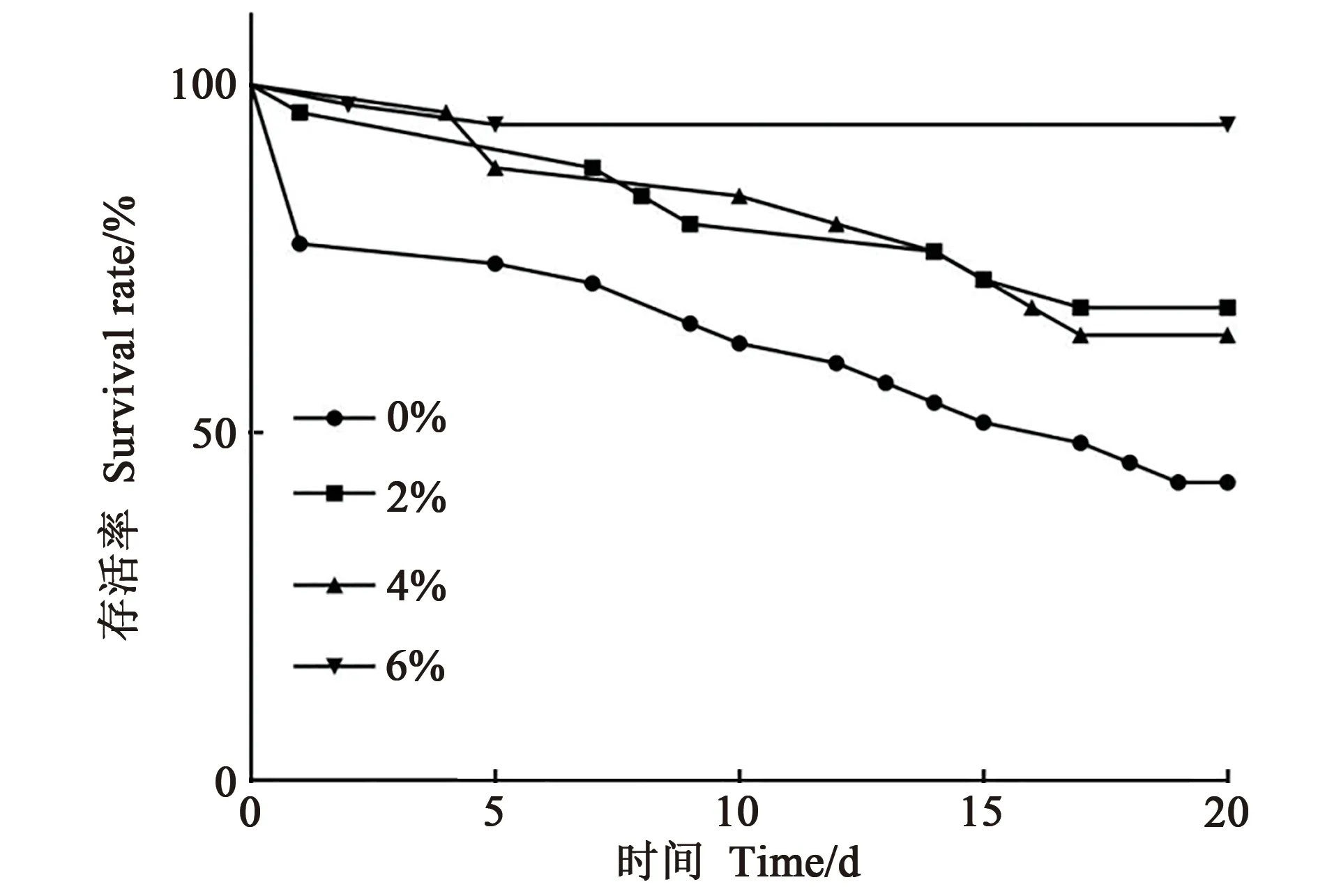
图2 不同赖氨酸水平下工蜂存活率Fig.2 Survival rate of worker bees under different lysine levels
2.3 工蜂血淋巴代谢组学分析
2.3.1 主成分分析 对饲喂赖氨酸水平为0%与6%的工蜂血淋巴代谢物进行主成分分析,结果如图3 所示。可以看出,0%的赖氨酸饲喂组与6%饲喂组差异明显。为了更准确地显示2 种赖氨酸含量对工蜂血淋巴的代谢物影响,使用正交偏最小二乘法-判别分析(OPLS-DA)模型进行了深入分析,血淋巴代谢物OPLS-DA 得分如图4 所示。对模型进行排序验证,参数为:R2X(cum)=0.725。R2Y(cum)=0.998,Q2(cum)=0.996,Q2>0.5,且Q2与R2Y相接近,表明模型稳定可靠,没有出现过拟合现象。

图3 PCA得分Fig.3 PCA score scatter plot

图4 OPLS-DA 得分图Fig.4 OPLS-DA score scatter plot
2.3.2 差异代谢物的鉴定与分析 如表2 所示,通过OPLS-DA 分析,根据VIP>1、P<0.05、FC 值>1.5 或FC 值<1∕1.5 的标准,经KEGG、HMDB 数据库检索,共鉴定出28 种差异代谢物,包括氨基酸及其衍生物、嘌呤类化合物等,其中,含量上升的物质有10 种,下降的物质有18 种。哌啶酸、L-酵母氨酸、黄嘌呤上升差异倍数最为显著,可能是饲粮中赖氨酸含量增加导致。而过量赖氨酸可能影响其他氨基酸的吸收,如组氨酸等。

表2 差异代谢物Table 2 Differential metabolites
2.3.3 KEGG 富集分析 KEGG 通路富集显示,28 种差异代谢物富集到了20 条代谢通路(表3),其中,5 条具有显著差异的富集通路:组氨酸代谢,丙氨酸、天冬氨酸和谷氨酸代谢,精氨酸生物合成,嘌呤代谢,泛酸和辅酶A生物合成。

表3 KEGG 通路富集详情Table 3 Enrichmegnt of KEGG pathway
3 讨论
3.1 饲粮赖氨酸水平对工蜂生长的影响
赖氨酸作为必需氨基酸,其主要作用是参与机体蛋白质合成,如血清蛋白、酶、多肽、激素等。长期进食低赖氨酸饲料会降低大鼠体内血清球蛋白与β-球蛋白的含量抑制肝脏蛋白质的合成,影响大鼠的生长[30]。当饲料中赖氨酸水平增加时,大鼠大脑、肝脏和肾的蛋白含量也随之增加[31]。当赖氨酸水平为6%时,工蜂胸部肌肉蛋白含量达到为47.53 mg·mL-1。
工蜂的寿命是衡量熊蜂授粉价值的重要指标,饲粮中不同赖氨酸水平对其寿命影响很大,当饲粮中赖氨酸水平为0%时,工蜂存活率最低,且随着时间的增加死亡数量也在增加。这可能是熊蜂饲粮中缺乏赖氨酸,导致一些蛋白无法合成,从而无法满足工蜂生长发育的需求。当赖氨酸水平增加至2%和4%时,工蜂死亡数量减少;当赖氨酸水平增加至6%时,工蜂仅在前5 日内有工蜂死亡,且死亡数量最少,与其他3 组存活率存在极显著差异。综上可知,饲粮中赖氨酸水平为6%时,工蜂存活率最高。随着赖氨酸水平增加,工蜂生存率随之上升,这可能与体内蛋白质含量增加有关。
3.2 赖氨酸对工蜂代谢的影响
在5 条显著富集通路中,组氨酸代谢为极显著(P<0.01)富集通路。有3 种显著差异代谢物富集到了组氨酸代谢通路中,其中2 种为下调,包括组胺、尿刊酸,L-天冬氨酸为上调。在血淋巴代谢物中,组氨酸含量出现下调(FC=0.80),这可能与赖氨酸水平过高有关。赖氨酸水平过高可能会抑制肠道对其他氨基酸的消化吸收,从而使其他氨基酸在机体内含量降低。研究发现,不同赖氨酸水平饲粮喂食生长猪,其肝门静脉血清中组氨酸含量随着赖氨酸水平的增加而降低[32]。组氨酸作为必需氨基酸之一,参与蛋白质的合成,是动物体血红蛋白的重要组成部分,其代谢产物具有调节神经系统、维持机体健康的作用。在组氨酸代谢途径中,组氨酸在组氨酸转氨酶的作用下产生尿刊酸,参与紫外线引起的免疫抑制作用[33]。组胺由组氨酸脱羧而产生,组胺作为神经元与脑部神经递质,参与神经系统的调节[34]。除此之外,组氨酸还可作为中间产物参与谷氨酸、天冬氨酸等氨基酸的合成[35]。
天冬氨酸为非必需氨基酸,但外源性天冬氨酸可缓解三羧酸循环中间产物如草酰乙酸和苹果酸的缺失,从而提高ATP的合成[36-37]。α-酮戊二酸是三羧酸循环的中心物质,参与多条代谢途径,对细胞新陈代谢有着重要作用[38]。本研究结果显示,天冬氨酸与α-酮戊二酸显著富集到精氨酸生物合成与丙氨酸、天冬氨酸和谷氨酸代谢两条途径中,且这两种代谢物均显著(P<0.05)上调。精氨酸与赖氨酸都属于必需氨基酸,精氨酸为一氧化氮与多种生物活性物质合成的前体,对机体的生长发育有着重要作用。研究表明,猪饲料中过量赖氨酸与精氨酸不存在拮抗作用[39]。在本研究中,赖氨酸水平为6%时,血淋巴代谢物精氨酸含量上升,赖氨酸与精氨酸未出现拮抗作用。高水平赖氨酸是否与精氨酸存在拮抗作用尚需进一步研究。
嘌呤在动物体内主要以嘌呤核苷酸的形式存在,在作为能量供应、代谢调节等方面有着重要作用。腺苷是合成腺嘌呤的重要中间体。本研究中嘌呤检测出差异的有腺嘌呤、黄嘌呤。腺嘌呤、黄嘌呤与腺苷一起富集到嘌呤代谢途径中,最终生成尿酸。黄嘌呤、尿酸在饲粮6%赖氨酸组饲喂下工蜂血淋巴中含量高于0%赖氨酸组。在昆虫体内,嘌呤代谢的最终产物是尿酸[40]。尿酸具有抗氧化功能,可减少自由基对蛋白质、DNA 及膜脂质的损害[41]。但是,饲粮赖氨酸水平在6%下工蜂血淋巴代谢物腺苷、腺嘌呤呈下调趋势,对于这一现象的原因是否与赖氨酸含量过高有关还需进一步研究。
辅酶A是一种含有泛酸的辅酶,是多种酶反应通路的辅助因子,参与物质代谢,可增强机体免疫力[42]。辅酶A的乙酰化形式是乙酰辅酶A,碳水化合物、蛋白质、脂质都会通过乙酰辅酶A进入三羧酸循环。试验结果检测出差异倍数较大代谢物为L-酵母氨酸、哌啶酸,且含量均为上调,这可能与赖氨酸分解代谢有关。赖氨酸可经过酵母氨酸途径与哌啶酸途径转化为乙酰辅酶A参与TCA循环。
在本研究中,饲粮中赖氨酸水平的增加提高了熊蜂工蜂胸部肌肉蛋白含量与工蜂存活率,赖氨酸水平为6%时胸部肌肉蛋白含量与存活率最高。采用LC-MS 方法检测饲喂赖氨酸缺乏组与6%赖氨酸组工蜂血淋巴代谢物,筛选鉴定出28种差异代谢物,主要涉及氨基酸与嘌呤的合成代谢途径。综上所述,考虑到饲粮6%赖氨酸组工蜂血淋巴代谢物中组胺、腺嘌呤等代谢物下降,熊蜂工蜂饲粮中赖氨酸水平为4%~6%时,更适合幼年工蜂的生长发育与生存。
本文研究了不同赖氨酸水平对幼年地熊蜂的生长及其血淋巴代谢物的影响,有助于探明地熊蜂工蜂赖氨酸的营养需求,更好地了解氨基酸在熊蜂饲粮中的重要性,为熊蜂规模化饲养和授粉应用中的营养补充提供部分理论依据。
猜你喜欢
第二课堂(课外活动版)(2022年5期)2022-05-25
第二课堂(课外活动版)(2022年1期)2022-04-23
湖南饲料(2021年4期)2021-10-13
今日农业(2021年6期)2021-06-09
第二课堂(课外活动版)(2021年12期)2021-01-18
第一财经(2019年8期)2019-08-26
时尚育儿(2018年8期)2018-05-14
儿童故事画报·发现号趣味百科(2017年10期)2018-03-13
儿童故事画报·发现号趣味百科(2017年9期)2018-03-13
儿童故事画报·发现号趣味百科(2016年4期)2016-06-24
