
木质纤维素预处理衍生抑制物对产溶剂梭菌的胁迫机制及解抑制策略研究
2022-12-02 00:49侍永江周泰然郭羽芬唐士杰罗洪镇
南京工业大学学报(自然科学版) 2022年5期
刘 正,侍永江,周泰然,高 磊,郭羽芬,唐士杰,罗洪镇
(淮阴工学院 生命科学与食品工程学院,江苏 淮安 223003)
化石能源是当前世界上最重要的能量来源,在经济发展、人类和社会进步中发挥了至关重要的作用。化石能源过量开采造成了资源枯竭、环境恶化等一系列问题,因此亟须开发绿色、可再生的高效能源。利用生物燃料部分替代化石燃料的潜力已逐渐凸显。在2017年当年全球能源供应中,可再生生物质供能占比约10.3%[1]。目前已有87个国家制定了使用可再生能源的经济目标,使得涉及人类粮食及能源安全的生物经济越来越受重视[2]。目前储量极为丰富的木质纤维素资源已由多种方式转化为丁醇、乙醇、柴油等多种燃料,得到研究者广泛关注[3]。
木质纤维素具有储量丰富、可再生、低成本等天然优势[4],可用于生产高效生物燃料,如,具有高热值、低蒸气压、低挥发性的丁醇[5]。目前,生产丁醇的微生物主要是产溶剂梭菌,如,拜氏梭菌(Clostridiumbeijerinckii)和丙酮丁醇梭菌(C.acetobutylicum),产溶剂梭菌通过复杂的代谢网络将底物转化为丙酮、乙醇和丁醇的混合物,其中丁醇占总溶剂的质量分数约60%。由于梭菌无法直接利用木质纤维素原料,需要对木质纤维素原料进行预处理,以提高后续发酵性能,而预处理产生的抑制物是限制木质纤维素高值化利用的主要瓶颈[6]。基于此,理解木质纤维素生物质预处理中衍生的抑制物特征及其对产溶剂梭菌的抑制机制,并建立有效的解抑制工程策略,是实现有效利用木质纤维素资源合成燃料丁醇的关键。本综述聚焦上述问题,介绍预处理中的抑制物衍生特征,重点关注产溶剂梭菌对抑制物的胁迫响应机制及解抑制策略,并讨论木质纤维素高效生物转化的发展前景。
1 木质纤维素预处理抑制物的衍生特征
木质纤维素生物质是一种非均质聚合物,主要由纤维素、半纤维素和木质素构成,3种成分的质量分数分别为40%~50%、25%~30%和15%~25%;此外,还包含少量灰分、蛋白质等成分。木质纤维素各组分含量受种类、环境温度、湿度、地理位置等影响而有所差异。构成木质纤维素的三大组分中,纤维素是一种由葡萄糖单元组成的多糖,在酸或酶作用下,其连接键被破坏,从而产生可被微生物利用的葡萄糖;半纤维素是由五碳糖和六碳糖组成的多聚物;木质素是由几种不同的苯基丙烷单体通过芳香醚键随机连接组合而成的三维无定型聚合物。纤维素、半纤维素、木质素交联形成的致密结构具有极强的抗降解特性,很大程度上阻碍了其高效生物转化。预处理能够有效破坏木质纤维素细胞壁的致密结构,去除木质素的同时降低纤维素的结晶度,提高纤维素可及度和酶解效率[7]。由于不同预处理方式对木质纤维素的作用强度等不同,导致预处理衍生的抑制物质量浓度、种类也有一定差异。预处理过程中形成的抑制物主要包括:弱酸类物质、呋喃衍生物、酚类化合物(图1)[8]。弱酸类物质主要来源于纤维素降解形成的己糖经进一步脱水形成的5-羟甲基糠醛(5-HMF),弱酸类物质包括甲酸、乙酸、乙酰丙酸。其中,乙酸由半纤维素乙酰化和少部分木质素乙酰化形成;呋喃衍生物包括5-HMF、呋喃甲酸、糠醛等,而糠醛是由半纤维素过度降解而直接形成;酚类化合物主要由木质素组分水解形成,如,4-羟基苯甲醛、阿魏酸、丁香醛、香草醛等。
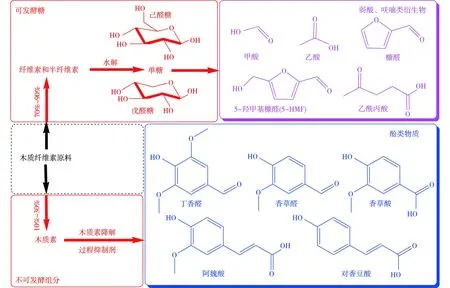
图1 木质纤维素预处理过程中产生的各类发酵抑制物[8]Fig.1 Fermentative inhibitors formation during pretreatment of lignocellulosic biomass[8]
化学预处理是木质纤维素预处理最常用的预处理技术,主要包括酸预处理、碱预处理以及有机溶剂预处理。其中,酸预处理是一种高效且应用广泛的预处理工艺。Gu等[9]用稀H2SO4预处理甘蔗秸秆时,木聚糖脱除率和糖得率分别达到88%和90%。酸预处理会使得半纤维素降解产生乙酸,同时部分纤维素降解产生的己糖脱水形成5-HMF。糠醛和5-HMF在各种水解液中的质量浓度差异较大,但普遍质量浓度均不超过3.5 g/L,甲酸、乙酰丙酸来源单一且不易形成,因此,质量浓度普遍较低(0.1~0.6 g/L),而乙酸的质量浓度最高可达8.0 g/L[10]。碱预处理通常在较为温和的条件下进行,与酸预处理的区别在于,碱预处理主要溶解木质素和半纤维素,断开分子间的酯键连接,以降低纤维素的结晶度和聚合度。Kim等[11]利用Na2CO3作为廉价碱催化剂,预处理玉米秸秆时,葡萄糖产率为267.5 g/kg,并有效破坏玉米秸秆的顽固结构。由于碱预处理主要去除木质素及半纤维素,因此所形成的抑制物主要包括甲酸、乙酸以及酚类化合物。相较于甲酸、乙酸等抑制物,酚类化合物被认为是对酶解及发酵的抑制作用最为强烈的一类抑制物[12]。大多数抑制剂具有良好的水溶性,而酚类化合物在水中的溶解度低、水溶性差,导致其检测浓度与水解液中的真实浓度有一定差异[13]。有机溶剂预处理的作用方式与碱预处理相似,利用有机溶剂和其水溶液可以破坏木质素与半纤维素之间的化学键。Jafari等[14]利用丙酮进行丁醇发酵的原料预处理,成功脱除69%木质素以及83%半纤维素,将葡聚糖质量分数提高至61%。然而,有机溶剂预处理同样无法避免发酵抑制物形成。表1总结归纳了不同预处理的优缺点及抑制物衍生特征[15-18]。
在木质纤维素水解液中,弱酸一般不会单独存在,往往与呋喃类衍生物、酚类化合物共同存在,对微生物产生协同抑制作用。弱酸的抑制效果相比于呋喃、酚类化合物的抑制效果较弱。因此,目前研究多数致力于呋喃类衍生物以及酚类化合物。其中,酚类化合物的低水溶性、成分复杂导致在其定性定量分析上存在一定难度,其抑制机制的解析应是值得重视的方向。
2 产溶剂梭菌对预处理抑制物的胁迫响应机制
2.1 弱酸类物质
Cho等[19]以C.acetobutylicum为发酵菌株,添加0.2 g/L甲酸能够略微提高丁醇发酵的总溶剂产量,而当甲酸添加量增至1.0 g/L时,总溶剂产量下降75%。此外,C.beijerinckii则因为其细胞膜含有甲酸脱氢酶,因而对弱酸有较高的耐受性,其溶剂合成能力并未受到弱酸的抑制。同时,丁醇发酵是典型的两阶段发酵过程,分为产酸期和产溶剂期,弱酸类物质会导致胞内pH的降低,形成过酸环境,梭菌自身厌氧呼吸产能较少,菌体需要消耗大量三磷酸腺苷(ATP)将胞内多余的质子排出,因而细胞的生长代谢受到限制。细胞内低质量浓度甲酸的存在,即可造成乙酸、丁酸过量积累,溶剂合成受到抑制,在梭菌发酵过程中出现“酸崩溃”现象,发酵相转型无法顺利进行[20]。

表1 不同预处理工艺的优缺点及产生的主要抑制物Table 1 Advantages and disadvantages of different pretreatment methods and major inhibitors
2.2 呋喃类衍生物
研究表明,低质量浓度的糠醛或5-HMF单独存在时,能够提高C.beijerinckii溶剂合成能力,但混合呋喃类物质则抑制菌株对木糖的利用能力,并降低溶剂产量[21]。在梭菌厌氧发酵过程中,糠醛和5-HMF可以被转化为对细胞毒性较低的糠醇和2,5-二羟甲基糠醇。然而,实际上预处理所形成的糠醛和5-HMF质量浓度较高,严重影响菌体代谢,破坏菌体细胞膜,降低糖酵解途径和三羧酸循环(TCA)中参与细胞代谢的关键酶活性,从而降低葡萄糖利用率。并且,糠醛和5-HMF的代谢转化过程依赖于细胞内还原力的参与,而胞内还原力水平是合成溶剂的关键因素。此外,呋喃化合物在酸性环境中可能发生水合反应而转化为甲酸以及乙酰丙酸。在这种情况下,多种抑制物共同存在会对细胞生长、产氢能力具有强烈抑制[22]。Liu等[23]系统研究了甲酸、乙酸、糠醛、5-HMF等主要抑制剂对C.acetobutylicum的抑制情况,结果表明,多种抑制剂胁迫下,糖酵解途径、三羧酸循环的关键酶活性降低,与丙酮-丁醇合成和氧化还原代谢相关的代谢产物以及关键酶均受到抑制。此外,参与糖异生、氧化三羧酸循环、氧化应激的过氧化物酶蛋白活性显著上升,表明梭菌对抑制剂胁迫下产生应激反应,并自发调节代谢,但这种适应机制是以降低丁醇合成效率为代价。
2.3 酚类化合物
酚类化合物主要来源于木质素组分的降解[24]。在预处理液中的含量较低,但是与相同质量浓度的糠醛、5-HMF相比,则具有更强的毒性。酚类化合物具有亲脂性,容易穿透细胞膜进入细胞内部,破坏细胞膜的完整性,引起细胞内部活性氧(ROS)积累,诱发细胞死亡。Liu等[25]对0.5 g/L阿魏酸胁迫下的C.beijerinckii进行转录组学分析(RNA-seq),结果表明,氧化还原相关辅助因子、核黄素代谢、双组分系统、DNA复制相关基因产生差异表达。利用基于RNA-seq的加权基因共表达网络分析手段,对多种典型酚类化合物存在下丁醇发酵过程的研究表明:对香豆酸主要影响细胞核糖体蛋白的基因表达及鞭毛的组装、DNA重组;丁香醛与香草醛则显著影响嘌呤、淀粉、蔗糖的代谢及产孢和信号转导功能;阿魏酸则引起糖基转移酶相关基因表达代谢紊乱[26]。此外,笔者研究团队借助发酵动力学结合转录组学探讨了酚酸、酚醛分别胁迫下的丁醇发酵性能,结果表明,酚醛比酚酸对C.acetobutylicum细胞生长和溶剂合成具有更强的抑制,酚类化合物改变了细胞的膜转运蛋白、磷酸转移酶系统、糖酵解相关的基因表达[27]。此外,木质纤维素生物质真实水解液中的酚类化合物应同时包含酚酸、酚醛,本团队进一步探究了产溶剂梭菌在复合酚类化合物存在下的胁迫机制,结果表明复合酚类化合物胁迫12 h后,葡萄糖利用效率降低21%。RNA-seq分析结果表明,复合胁迫条件使得与孢子生成、细胞分裂、抗氧化活性、膜转运蛋白、热休克蛋白等关键生理过程相关的基因均出现差异表达(图2),并绘制了产溶剂梭菌耐受酚类化合物的全局抑制机制图[28]。以上研究结果将为后续菌株抗逆性能提升给予理论指导。
由2.1~2.3节可知,弱酸类物质、呋喃类衍生物、酚类化合物这三种物质对产溶剂梭菌的抑制作用存在差异。刘欢欢等[29]选取糠醛、5-HMF、甲酸、乙酸和苯酚等作为主要抑制剂进行耐受性研究,代谢组学分析结果表明,低质量浓度的甲酸和苯酚(0.5 g/L)对丁醇发酵抑制作用强于高质量浓度的糠醛(2 g/L),而低质量浓度的呋喃类化合物(1 g/L)能够提高丁醇发酵性能,并且苯酚胁迫主要影响脂肪酸代谢和核酸代谢,而糠醛能够导致胞内还原性化合物积累出现紊乱,甲酸胁迫则主要使胞内代谢活动迟缓,菌体生长缓慢。综上所述,木质纤维素真实水解液中的抑制剂种类繁多,其浓度特征也随预处理方法的不同而出现显著差异(表1),并且抑制剂之间可能存在协同抑制作用。
3 解除抑制效应的工程策略
3.1 脱毒策略
解除预处理衍生物抑制效应最直接的方式是脱毒工艺,其使得预处理液中的抑制物浓度降低。不同脱毒工艺的优缺点如表2所示。一般可采用化学试剂与抑制物反应使得抑制物的毒性降低,或者采用吸附、分离等物理方式除去抑制物,它们都属于发酵之前脱除抑制物的方法。常用的脱毒方法包括:水洗、过碱中和、活性炭吸附、离子交换吸附、电化学脱毒、电凝处理等。
水洗通常使用大量的水对木质纤维素生物质预处理后进行重复、多次洗涤,速度较快,但存在诸多缺点如:产生大量废水,损失可发酵糖、抑制物去除不彻底等[30]。过碱中和通常使用Ca(OH)2与抑制物进行反应,该方法能够有效去除抑制物。Zhang等[31]利用碱和活性炭脱除水解物中的抑制剂,呋喃衍生物和芳香单体的去除率达到75.6%和68.1%,并除去较多酚酸物质。但是其缺点也较为明显,会损失部分可发酵糖;用于脱毒的试剂无法回收利用;过程中产生的沉淀及盐会影响后续的糖化、发酵,降低了工艺经济性。由于水解液中的抑制物多数为羰基化合物,如,羧酸(乙酸、阿魏酸)、木质素衍生的酚醛(丁香醛)、糠醛、5-HMF。含有羰基的这类化合物,其碳氧双键容易极化,在碳原子上产生正电荷,使得碳原子可以与核酸、蛋白质等生物亲核试剂形成共价键。因此,大多数带有伯氨基和侧链官能团的氨基酸能够与上述羰基化合物发生反应,从而降低其抑制发酵的能力。如带有巯基和胺基的半胱氨酸,容易与亲电的羰基反应,解除这类化合物的抑制。Xie等[32]利用多种亲核氨基酸选择性解毒生物质水解物,结果表明,半胱氨酸、组氨酸、赖氨酸、色氨酸和天冬酰胺表现出良好的解毒效果。以半胱氨酸作为亲核氨基酸处理时,96%的5-HMF被脱除,产品质量浓度大幅提高。
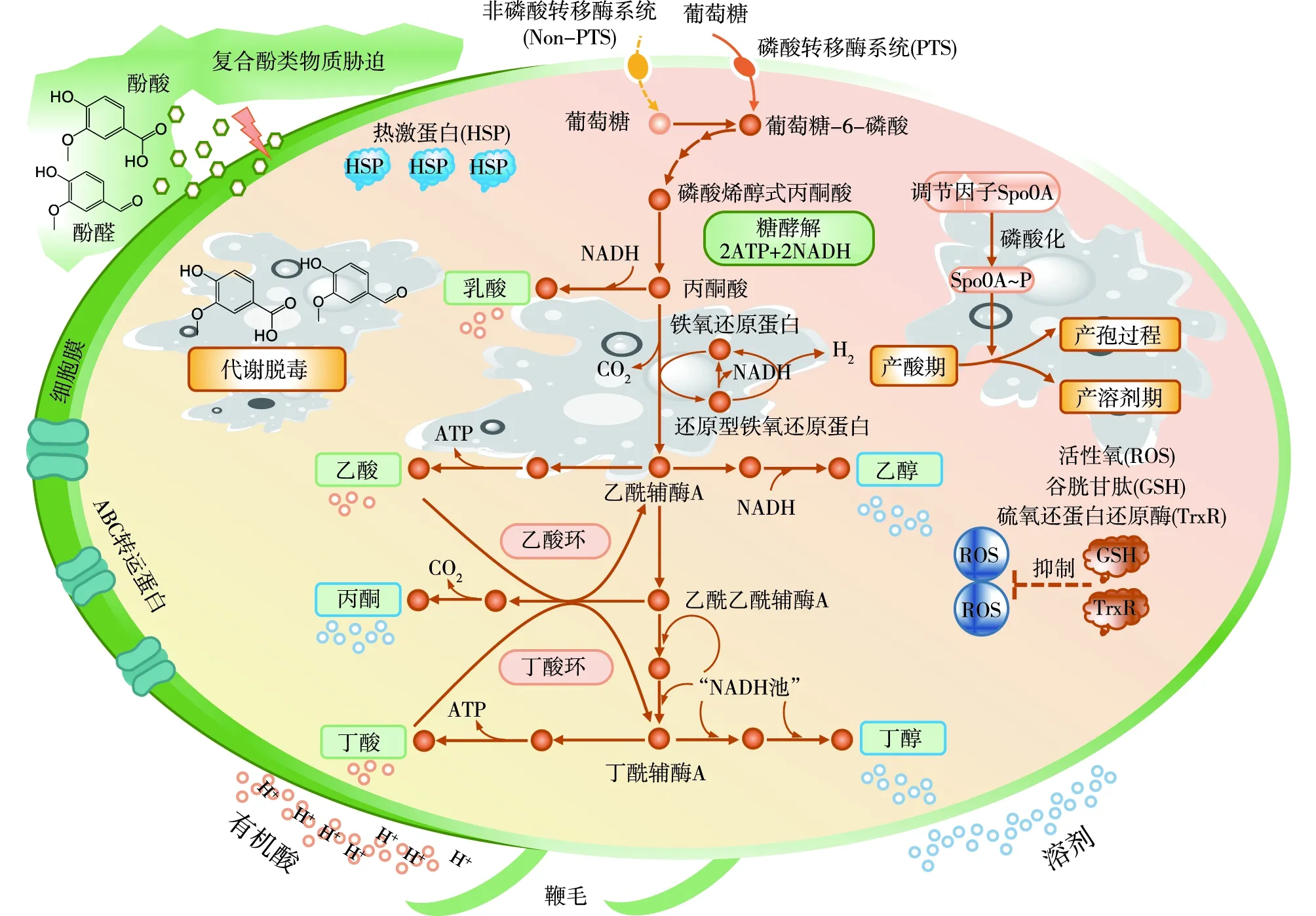
图2 C. acetobutylicum ATCC 824对复合酚类化合物胁迫的整体响应机制[28]Fig.2 The overall response mechanism of C. acetobutylicum ATCC 824 to combined phenolic stress[28]

表2 木质纤维素原料预处理液脱毒方式及其优缺点Table 2 Detoxification methods of lignocellulosic hydrolysate and their advantages and disadvantages
此外,许多物理化学方法在脱除抑制物方面效果良好,并且能够保证极低的糖损失率。Jiang等[33]采用电化学解毒的方法处理玉米芯水解液24 h后,酚类化合物下降约40%,呋喃及糠醛质量浓度也有不同程度的降低,最终丁醇产量是未解毒批次的1.6倍,并且在整个过程中避免了可发酵糖的损失以及副产物的形成。Jeong等[34]使用Al电极在8 g/L的NaCl中电凝处理3 h,成功去除了47.8%酚类化合物以及大部分5-HMF和糠醛,仅有约2%的糖损失率。
生物脱毒法指通过微生物产生的酶脱除抑制剂,能够实现抑制剂原位脱毒、降低操作成本、提高整体发酵性能。从真菌中分离出来的漆酶近年来广泛应用于木质素的解聚过程,其结构域有4个不同的Cu离子,能够介导单酚、多酚、芳香胺、无机离子等多种底物的氧化。Allard-Massicotte等[35]在过滤、絮凝等物理化学脱毒方式的基础上,以酶加量为100 mg/g的脱毒条件,成功脱除水解液中包含香草醛、丁香醛、邻苯二酚、没食子酸在内的复合酚类化合物,降解率达77%,水解液脱毒后酚类化合物总质量浓度仅为0.28 g/L,低于其抑制微生物代谢的最低值,最终提高一倍以上的丁醇产量。近期研究从腐烂木材中分离得到真菌,并纯化出具有良好热稳定性、pH稳定性的漆酶,在甘蔗渣中脱毒5.5 h即可去除79.28%的酚类抑制剂,具有一定的工业应用潜力[36]。
3.2 代谢工程改造策略
微生物对发酵抑制剂中呋喃类、酚类化合物的代谢方式主要包括氧化还原、转运调控等。其中,通过氧化还原反应将高毒性的醛类转化为具有较低毒性的酸和醇,二者再进入微生物的中心碳代谢从而实现呋喃、酚类化合物的最终降解[37-38]。微生物对弱酸类物质的代谢机制与其他两类抑制物不同,主要通过乙酸代谢、重吸收实现代谢。乙酸在乙酰辅酶A合成酶作用下,合成乙酰辅酶A,进入三羧酸(TCA)循环,乙酰丙酸先转化为乙酰丙酸辅酶A后进一步被裂解,形成甲酰辅酶A和乙酰辅酶A,进入TCA循环,完成代谢。Ezeji等[39]研究表明,C.beijerinckiiBA101对低质量浓度的乙酸、糠醛、5-HMF有较强的耐受性,糠醛和5-HMF在3 g/L下能被C.beijerinckiiBA101代谢利用,从而提高发酵性能。进一步研究发现,酚类化合物如香豆酸、阿魏酸在2 g/L质量浓度下,即可以完全抑制产溶剂梭菌的发酵,0.5 g/L香豆酸使得丁醇产量降低了90%[40]。因此,提高菌株对酚类化合物耐受性是丁醇发酵需解决的关键问题。Guo等[41]通过连续培养分离出具有高抑制物耐受性的C.beijerinckii,以未经脱毒的预处理液作为底物(含2.1 g/L酚类化合物和40.3 g/L还原糖)时,得到2.6 g/L丙酮、7.5 g/L丁醇和0.5 g/L乙醇。Lu等[42]通过基因工程技术构建了过表达乙醛脱氢酶/乙醇脱氢酶基因(adhE2)和辅酶A转移酶基因(ctfAB)的工程菌C.beijerinckiiCC101-SV6,与仅表达adhE2的C.beijerinckii工程菌株相比,其溶剂合成及有机酸吸收的能力显著提高。同时C.beijerinckiiCC101-SV6能合成约12 g/L丁醇,并高效吸收有机酸,有效防止“酸崩溃”的发生;而且该菌利用葡萄糖和木糖的能力也有所提高,表现出对抑制剂的高耐受性。只是单一过表达乙醛脱氢酶/乙醇脱氢酶可能增加了发酵初期代谢负担,使得胞内ATP供应不足,还原力失衡,因而,部分乙酰辅酶A无法正常代谢有机酸。Jiménez-Bonilla等[43]过表达Pseudomonasputida的外排泵基因提高了C.saccharoperbutylacetonicum对阿魏酸的耐受性,结果表明,整体基因的过表达降低了菌株对抑制剂的耐受性,而单一过表达亚基基因(srpB)就能够显著提高菌株的鲁棒性,最终丁醇产量能够达到不添加抑制剂批次的水平。Wu等[44]以过表达或敲除编码葡萄糖特异性的磷酸转移酶系统相关基因,构建了C.acetobutylicum工程菌株,由于该基因具有调控葡萄糖利用效率、溶剂合成等多种功能,因而在指数生长期能量/辅因子再生、溶剂合成和抑制剂耐受水平均得到提高,最终表现出较好的丁醇发酵性能。
3.3 统合生物加工及微生物共培养策略
统合生物加工(CBP)是将原料水解、酶水解及后续溶剂发酵过程整合为一步的工艺,因而,产溶剂梭菌能够直接利用原料生产丁醇等溶剂。将两种或多种微生物同时进行培养,利用底物进行发酵,实现解除抑制物抑制效应的同时进行溶剂的合成。从经济角度,该工艺能够降低甚至减少昂贵的预处理成本,提高经济可行性;从技术角度,共培养能够实现不同微生物之间的功能互补,扩展底物谱,提高其对产物和抑制剂的耐受能力,高效合成溶剂。Wen等[45]开发出一种基于双梭菌共培养的CBP系统,以纤维素梭菌(C.cellulovorans)和C.beijerinckii作为发酵菌株,通过敲除乙酸激酶基因和乳酸脱氢酶基因,并过表达丁酸激酶基因,转变碳代谢更多流向丁酸生成方向;过表达与有机酸重吸收相关的基因(ctfAB),以提高乙酸转化速率。利用双梭菌体系实现了纤维素酶生产、木质纤维素水解和己糖/戊糖共发酵,一步生成丁醇,发挥纤维素梭菌能够直接利用纤维素生成可发酵糖的优势,避免发酵抑制物的形成。Cui等[46]将C.beijerinckiiG117与重组枯草芽孢杆菌(Bacillussubtilis1A1)共培养,B.subtilis消耗培养基中的O2后,为C.beijerinckii创造厌氧环境,并且B.subtilis产生的乳酸被C.beijerinckii重吸收,B.subtilis1A1通过表达二级醇脱氢酶将C.beijerinckiiG117产生的丙酮还原为异丙醇,维持胞内氧化还原平衡。利用不同菌株之间生理生化性质、代谢途径和能力的差异,不仅成功实现发酵抑制物的原位降解,而且能够同步合成目标产物。不足的是,许多微生物共培养体系中,菌株之间在分子水平上的微生物相互作用机制仍未可知。在工业化应用中存在以下问题:如何获取适用于CBP技术的微生物菌株、工艺中的多种水解酶如何高效共表达、整体发酵协同性如何提高、水解酶之间的组分比例如何调控等。此外,为提高微生物共培养系统的性能,使微生物更好地发挥相互作用,以后可重点从氧传递速率控制、菌株种群比例调控、共培养系统分子机制解析等方面开展研究。
代谢工程改造策略提高细胞代谢鲁棒性是近年研究的热点方向。然而木质纤维素衍生抑制剂对产溶剂梭菌的影响是全局性的,往往涉及细胞生长、溶剂合成、抑制剂代谢等多个过程,以代谢工程改造手段提高其耐受性难度较大。抑制物对产溶剂梭菌全局影响的机制极其复杂,若单独通过改造某一代谢通路或基因,无法达到预期效果。CBP和微生物共培养策略是提高产溶剂梭菌对产物丁醇及抑制剂耐受性的有效手段。但在其应用中仍然存在不同微生物之间的底物利用竞争问题。此外,由于不同微生物或酶的最适生长条件和最高活性条件的不同,共培养体系的性能无法发挥到极致,并且菌株之间在分子水平上的相互作用机制尚缺乏深入研究。通过解决上述问题将为产溶剂梭菌高产丁醇策略的提出提供指导。
4 结论与展望
木质纤维素预处理衍生的抑制物是限制生物炼制产业化发展的主要瓶颈。本文介绍了木质纤维素不同预处理工艺及抑制物衍生特征,并归纳总结了解除抑制的前沿工程策略,为木质纤维素资源转化提供了借鉴。当前,CBP系统、微生物共培养和代谢工程等技术已应用于解除抑制剂的抑制效应研究,但仍存在诸多难以解决的技术难题。除文中所述的解抑制策略之外,许多应用前景广阔的技术路线值得研究者们关注。
首先,木质纤维素预处理过程是一个高度复杂性、非线性特征的系统,抑制物的质量浓度衍生特征受原料、预处理工艺等因素的影响,运用传统的动力学模型难以进行准确预测。若能从预处理中减少抑制物的产生,进而降低后续脱毒成本,会提高整体工艺的经济性。目前,人工神经网络(ANN)技术作为一种有效的预测建模方法,具有快速寻找最优解的特点,适用于处理木质纤维素预处理等高度复杂的反应过程,在生物炼制领域有巨大的应用潜力,并且目前已被开发用于生物燃料生产、热解、生物质热力学性质和高热值预测等研究领域[47-48]。其次,基于适应性实验室进化(ALE)策略能够提高菌株耐受多种抑制物的能力,通过重复分批培养或在设定的胁迫条件下延长培养时间,使菌株发生所需的遗传变异[49];但该过程往往需要经历漫长的培养周期。因此,利用合成生物学设计并构建相关生物元件和基因文库,借助基因、转录、蛋白、代谢等组学手段,挖掘与产溶剂梭菌的抑制物耐受性相关的关键基因,实现对细胞生理活动、代谢变化、基因表达的精准把控。在DNA、RNA、蛋白质水平调控产溶剂梭菌的代谢网络,减少副产物生成,避免中间产物积累,维持菌株内部代谢平衡,最终实现产溶剂梭菌直接利用木质纤维素原料或提高细胞耐受性的生物功能。近期研究者组合利用ALE和转录组学分析显著提高了Yarrowialipolytica对芳香醛的耐受性,由转录组分析和反向代谢工程手段证实了过表达醛酮还原酶基因和醛脱氢酶基因可将芳香醛转化为相应的醇和酸,从而获得对芳香醛具有高耐受的进化株[50]。最后,还需关注的科学问题是丁醇的产物抑制效应,这是限制丁醇高效发酵的瓶颈之一,约5 g/L丁醇就可抑制产溶剂梭菌的正常生理代谢。因此,利用原位分离耦合技术将溶剂在发酵过程中分离出来,能够减少溶剂对细胞毒害,提高整体发酵性能。最新的研究已通过构建一系列针对不同菌株的基因编辑工具,以引入外源基因等方式系统改造产溶剂梭菌,获得高效生产高附加值的脂肪酸酯等产品的工程菌株,大幅提高了生产效率[51]。未来,利用从基因元件、模块到整体代谢系统的合成生物学等新兴交叉学科的优势,结合基因组学、转录组学、蛋白质组学、代谢组学等手段,从多角度研究生物系统,获得更多优良的工业发酵菌株,最终实现木质纤维素生物质高效生物转化的工业化目标。
猜你喜欢
煤炭与化工(2022年7期)2022-11-17
当代水产(2022年1期)2022-04-26
当代水产(2022年2期)2022-04-26
小资CHIC!ELEGANCE(2021年41期)2021-11-08
中国饲料(2021年17期)2021-11-02
当代水产(2021年2期)2021-03-29
颂雅风·艺术月刊(2021年3期)2021-03-17
分析仪器(2020年5期)2020-11-09
老友(2020年2期)2020-03-08
安徽农业科学(2019年11期)2019-07-10
