
柑橘黄酮类化合物的提取新技术及生物活性研究进展
2022-12-01 12:20甘人友
食品与机械 2022年11期
刘 丹 郭 欢 吴 笛 甘人友,3
(1. 成都大学食品与生物工程学院,四川 成都 610106;2. 中国农业科学院都市农业研究所,四川 成都 610213;3. 国家成都农业科技中心,四川 成都 610213)
柑橘(Citrus)属芸香科柑橘亚科植物,含有丰富的生物活性物质,其中黄酮类化合物是柑橘中重要的功能活性物质。黄酮类化合物的基本骨架为C6—C3—C6,即中央三碳链连结两个芳香环构成[1]。柑橘品种较多,不同品种中黄酮类化合物含量不同。目前,柑橘黄酮类化合物的提取方法主要有碱提法、醇提法、酶法、微波辅助提取法和超声波辅助提取法等。近年来,许多研究学者建立新的安全、环保高效的提取方法,大大增加了柑橘黄酮类化合物的提取率,且大量研究学者对柑橘中黄酮化合物的生物活性进行了研究,如抗癌、抗肥胖和肠道保护等。研究拟综述近5年有关柑橘中黄酮类化合物提取新技术及其生物活性,并重点阐述其相关作用机制,以期为柑橘黄酮类化合物的开发利用提供科学依据。
1 柑橘黄酮提取新技术
1.1 低共熔溶剂提取技术
低共熔溶剂通常是由季铵盐、季盐等氢键受体和酰胺、羧酸、醇等氢键供体在一定比例下通过氢键结合形成的低熔点混合物[2]。Xu等[3]以柑橘皮为原料,探讨了基于氯化胆碱的低共熔溶剂的黏度、pH值、极性对黄酮提取效率的影响。结果表明,柑橘黄酮的提取率与氢键供体的极性呈线性关系,且氯化胆碱-乙酰丙酸-N-甲基尿素的总黄酮提取率最高。在最佳提取条件下,柑橘果皮中多甲氧基类黄酮为18.75 mg/g,类黄酮苷为47.07 mg/g,回收率分别为95.87%,86.32%。Jokic等[4]采用15种不同的以氯化胆碱为氢键受体的低共熔溶剂对4个不同品种柑橘的柑橘皮进行提取,以橙皮苷得率为指标进行筛选。结果表明,氯化胆碱—乙酰胺(摩尔比为1∶2)的提取效率最高,橙皮苷得率为112.14 mg/g。与传统溶剂提取相比,低共熔溶剂具有降解性好、易制备、低成本和低毒性等优点。
1.2 亚临界流体萃取技术
亚临界萃取是根据有机物相似相溶的原理,利用亚临界流体作为萃取溶剂,在密闭、无氧、低压的压力容器内萃取物料脂溶性成分,再通过减压蒸发将萃取剂与目的产物分离的萃取技术[5]。Kim等[6]用亚临界水半连续模式提取蜜柑皮中的生物活性黄酮类化合物。结果表明,橙皮苷和芸香柚皮苷的得率随温度(145~165 ℃)升高而增加。此外,Lachos-Perez等[7]以脱脂橘皮为原料,研究了温度和溶剂流速对亚临界水提取黄酮得率的影响。结果表明,在150 ℃、10 mL/min条件下获得最大产率,即橙皮苷(188.74±0.51) mg/g,柚皮芸香苷(21.98±1.39) mg/g。与索氏提取法、常规溶剂提取法和超声波提取法3种传统提取方法相比,亚临界水提取是一种高效提取活性化合物的方法。
1.3 磁性分子印迹聚合物选择性提取法
分子印迹技术是一种由模板、功能单体和交联剂制备的高分子功能材料以展示其选择性分子识别行为的技术,具有专一识别性、预定性和实用性等特点[8]。磁性分子印迹聚合物是利用分子印迹技术在磁性微粒表面合成具有超顺磁性、高选择性、高吸附性和特异性识别的聚合物,是一种能在外加磁场的作用下实现快速分离的新型高分子材料[9]。贺美艳[10]利用分子印迹技术的原理,以单一类黄酮橙皮素、桔皮素为模板分子,分别使用2-乙烯基吡啶、丙烯酰胺为功能单体,乙二醇二甲基丙烯酸酯为交联剂,乙腈为致孔剂,采用沉淀聚合法制备得到具有类特异性吸附、高选择性的分子印迹聚合物微球,用聚合物微球做填充材料制备分子印迹固相萃取柱,利用其类特异性吸附的特点实现了柑橘复杂基质中类黄酮的高效分离与纯化。而Wang等[11]采用以功能化Fe3O4颗粒为磁芯,橙皮素为模板,N-异丙基丙烯酰胺为功能单体,乙二醇二甲基丙烯酸酯为交联剂,2,2-偶氮二异丁腈为引发剂,乙腈—甲醇(V乙腈∶V甲醇=3∶1)为致孔剂的表面分子印迹法成功制备了新型橙皮素磁性分子印迹聚合物作为吸附剂,用于提取和富集干柑橘皮中的橙皮素。结果表明,干橘皮中橙皮素的回收率为90.5%~96.9%。线性范围为0.15~110.72 g/mL,相关系数大于0.999 1,该磁性分子印迹聚合物是一种具有选择性吸附的材料。磁性分子印迹聚合物选择性提取法与传统提取方法相比具有更好的选择性。
1.4 其他提取技术
除了上述提取新技术之外,还有闪式提取、3D打印技术等应用于柑橘黄酮提取。Liu等[12]将橙皮粉和乙醇置于120 V闪式提取器中进行提取,黄烷酮提取率为437 mg/g。与热提取相比,闪式提取法耗时更少,且能减少提取过程中热敏性化合物的损失。Subhi等[13]建立了涂布C18树脂的3D打印光盘在线流动系统耦合HPLC-DAD用于提取、分离和测定柑橘外皮中的黄酮类化合物的方法,能有效测定柑橘皮中的6种黄酮类化合物。该方法为黄酮类化合物的提取测定提供了一种全自动的、能有效减少提取时间和溶剂的消耗,且灵敏性高的方法。
2 柑橘黄酮类化合物的生物活性
柑橘黄酮具有多种生物活性,主要包括抗氧化、抗炎、抗菌、抗肥胖、抗糖尿病、抗癌、肠道保护、心血管保护、肝脏保护和神经保护等(图1)。
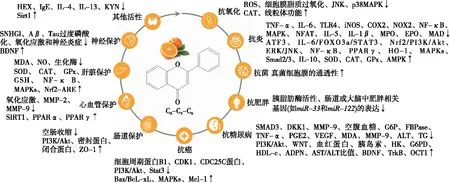
图1 柑橘黄酮生物活性及其作用机制Figure 1 Bioactivities of flavonoids from citrus and related mechanism of action
2.1 抗氧化
一些体外研究[14-15]采用DPPH、ABTS、FARAP等抗氧化评价方法证实了柑橘黄酮的抗氧化活性。对抗氧化活性起作用的主要黄酮类化合物为甜橙黄酮、4′,5,6,7-四甲氧基黄烷酮、川陈皮素、3,3′,4′,5,6,7-六甲氧基黄酮和柚皮芸香苷。且柑橘果皮中黄酮类化合物的种类和总含量均高于柑橘果肉。因此,柑橘果皮提取物的抗氧化能力更高[15]。
在Fe3+诱导氧化应激的A549细胞模型中,富含类黄酮的柑橘汁提取物能够降低活性氧(ROS)的产生、减少膜脂质过氧化、改善线粒体功能和防止DNA氧化损伤,其作用机制可能与铁的螯合性质及黄酮提取物增加抗氧化过氧化氢酶(CAT)活性有关[16]。香叶木苷是一种天然的黄酮类化合物,存在于各种柑橘类果实的果皮中。柑橘黄酮——香叶木苷通过减弱c-Jun氨基末端激酶(JNK)和p38丝裂原活化蛋白激酶(MAPK)信号通路,并降低细胞色素c、Bax和半胱氨酸天冬氨酸蛋白酶的表达水平,增加Bcl-2的表达,阻止高糖诱导的糖尿病视网膜病变模型ARPE-19人视网膜色素上皮细胞的氧化损伤[17]。简而言之,柑橘黄酮通过抑制ROS的产生、减少细胞膜脂质过氧化、增加CAT活性、改善线粒体功能以及抑制JNK和p38MAPK信号通路从而发挥其抗氧化活性,但其在体内的作用机制还需进一步的研究探讨。
2.2 抗炎
柑橘类黄酮化合物的抗炎作用在脂多糖(LPS)诱导的炎症小鼠巨噬细胞(RAW264.7)模型、小鼠模型和大鼠模型中得到证实。柚皮素可以抑制LPS诱导的炎症小鼠巨噬细胞中肿瘤坏死因子α(TNF-α)、白介素-6(IL-6)、toll样受体4(TLR4)、诱导型一氧化氮合酶(iNOS)、环氧合酶2(COX2)和烟酰胺腺嘌呤二核苷酸磷酸氧化酶2(NOX2)的表达,也可以抑制活化B细胞核因子κ-轻链增强子(NF-κB)和丝裂原活化蛋白激酶(MAPK)的激活并直接增加活化转录因子3(ATF3)的表达,从而改善炎症反应[18]。柑橘皮类黄酮——川陈皮素通过激活巨噬细胞中IL-6/信号转导及转录活化因子3(STAT3)/叉头框蛋白O3a(FOXO3a)信号通路增强自噬来抑制LPS诱导的炎症反应[19]。He等[20]研究发现柚皮素、川陈皮素和橙皮素对LPS诱导的RAW264.7细胞具有抗炎作用,其机制可能与TNF-α诱导的NF-κB通路抑制COX-2和iNOS表达有关。此外,Yang等[21]研究了柑橘类黄酮在佛波酯(PMA)/离子霉素诱导的EL-4小鼠T淋巴瘤细胞中降低活性氧(ROS)和白介素-5(IL-5)的作用机制,发现柑橘类黄酮通过不同途径抑制PMA/离子霉素诱导的EL-4细胞中的IL-5和ROS的产生。即橙皮苷通过转录因子红系衍生的核因子2相关因子(Nrf2)与磷脂酰肌醇3-激酶/蛋白激酶B(PI3K/AKT)或细胞外信号调节激酶(ERK)/Jun氨基末端激酶(JNK)信号通路结合诱导血红素氧合酶-1(HO-1)的表达,从而下调活化T细胞核因子(NFAT)活性和IL-5的分泌。同样,栀子宁通过降低EL-4细胞中NFAT活性和上调过氧化物酶体增殖物激活受体γ(PPARγ)诱导HO-1表达,进而抑制IL-5的产生,表明诱导HO-1表达可能抑制哮喘炎症[21]。
在卵清蛋白诱导的过敏性哮喘小鼠模型中,柑橘总黄酮通过调节MAPK和Smad2/3信号通路能有效地改善过敏性哮喘炎症并抑制气道重塑[22]。补充柑橘黄酮(圣草次苷或圣草酚)能减少LPS诱导的牙周炎症小鼠模型的牙龈上皮平坦度、慢性和急性炎症细胞浸润及牙龈乳头结缔组织的损失,显著地抑制牙周炎症。且圣草次苷和圣草酚均可抑制牙龈的白介素-1β(IL-1β)和TNF-α的表达,并增加白介素-10(IL-10)的表达,降低髓过氧化物酶(MPO)和嗜酸粒细胞过氧化物酶(EPO)活性,增加超氧化物歧化酶(SOD)、CAT和谷胱甘肽过氧化物酶(GPx)活性,降低丙二醛(MDA)含量[23]。在内毒素血症小鼠模型中,柚皮素通过激活5′-单磷酸腺苷活化蛋白激酶(AMPK)来增加ATF3的表达,从而改善炎症反应,提高生存率[18]。即柚皮素通过AMPK-ATF3依赖的LPS/TLR4信号通路负调控来保护小鼠免受内毒素血症。
2.3 抗菌
柑橘黄酮对铜绿假单胞菌和金黄色葡萄球菌等具有抑制作用。柑橘中芦丁对铜绿假单胞菌的最低抑菌质量浓度为800 μg/mL,同时用200 μg/mL芦丁和2.5 μg/mL抗生素庆大霉素处理时,对铜绿假单胞菌的生物膜的形成具有协同抑制作用[24]。低浓度的柚皮素通过改变苯丙氨酸、酪氨酸和色氨酸残基的微环境引起膜脂肪酸组成和膜蛋白构象的扰动,而高浓度柚皮素通过增加细胞膜的通透性引起金黄色葡萄球菌细胞形态发生改变[25]。DNA分子对接研究[25]表明,柚皮素通过沟槽结合再与基因组DNA的A-T碱基对区域结合,从而诱导了该生物大分子二级结构和形态的变化,即柚皮素的抗菌作用可能与金黄色葡萄球菌细胞质膜的破环和DNA靶向作用有关。此外,通过喂食柑橘类黄酮能显著降低虹鳟鱼感染鲁氏耶尔森菌O1生物型2的死亡风险[26]。
2.4 抗肥胖
胰脂肪酶是在胃肠道水解甘油三酯的一种关键酶。因此,抑制其活性可以减少脂质吸收从而有效改善肥胖症状[27]。柑橘皮中橙皮苷是主要的胰脂肪酶活性抑制剂[28]40。橙皮苷可以通过氢键和范德华力与胰脂肪酶相互作用,并且不改变胰脂肪酶的二级结构而降低胰脂肪酶的活性,改善肥胖[28]36-46。而圣草次苷、圣草酚和甜橙黄酮3种柑橘黄酮主要通过疏水作用力分别与胰脂肪酶周围的氨基酸残基相互作用,使胰脂肪酶周围的芳香族氨基酸残基所处微环境发生改变同时导致酶分子的二级结构发生变化,从而抑制酶活性改善肥胖[29]。作用机制的差异可能与黄酮化合物的结构有关,但还需进一步的研究确定黄酮化合物的结构与活性之间的关系。
此外,在细胞模型、肥胖斑马鱼和肥胖小鼠模型中,柑橘类黄酮化合物表现出显著的抗肥胖作用。在油酸诱导的脂质积累的人肝癌细胞HepG2中,柑橘皮黄酮提取物和柑橘纯黄酮混合物可能是通过抑制miR-33和miR-122的表达来增加肉碱棕榈酰转移酶1α(CPT1α)基因和减少脂肪酸合酶(FAS)基因的mRNA的表达,从而改善脂质代谢[30]。在饮食诱导的肥胖斑马鱼模型中,富含黄酮的橘汁通过调节肠道和大脑中一些肥胖相关的基因表达来改善肥胖,如瘦蛋白A、胃饥饿素、促食素、阿黑皮素原(POMC)和神经肽Y(NPY)[31]。在高脂肪饮食喂养的肥胖小鼠模型中,柑橘黄酮川陈皮素能有效地预防肥胖、肝脂肪变性、血脂异常和胰岛素抵抗[32]。但其在体内的作用机制不依赖于肝脏或脂肪细胞的腺苷酸活化蛋白激酶(AMPK)[32]。在高脂肪胆固醇饮食诱导肥胖的低密度脂蛋白受体基因敲除(Ldlr-/-)小鼠模型中,补充柑橘类黄酮可以通过增强能量消耗和增加肝脏脂肪酸氧化,逆转现有的肥胖和脂肪细胞的大小和数量[33]。在高脂饮食喂养的去卵巢肥胖小鼠模型中,补充柑橘类黄酮柚皮素可以抑制体重增加,降低高血糖,减少腹内脂肪增加,增加小鼠的自主活动,减少小鼠骨骼肌中甘油二酯的积累[34]。
2.5 抗糖尿病
在体外和动物研究中,柑橘黄酮表现出显著的抗糖尿病作用。在人肝癌细胞HepG2细胞模型中,柑橘黄酮香风草甙通过激活胰岛素抵抗性HepG2细胞中的PI3K/Akt信号通路促进葡萄糖摄取[35]。柑橘果皮中的多甲氧基黄酮则通过增加WNT信号通路的关键因子TCF7L2的表达,抑制SMAD3和DKK1的表达来调节血糖[36]20-26。
在链脲佐菌素(STZ)诱导的糖尿病大鼠模型中,柑橘类黄酮香叶木苷可能作用于肾上腺增强内啡肽的分泌,从而刺激阿片受体,减弱肝糖异生,增加比目鱼肌葡萄糖摄取,从而降低高血糖[37]。橙皮素通过改善STZ诱导的糖尿病大鼠的抗氧化能力来降低高血糖和高血脂[38]。柑橘类黄酮橙皮苷能够降低空腹血糖水平,增加血红蛋白和胰岛素水平,降低糖基化血红蛋白,增加大鼠肝组织中碳水化合物关键代谢酶己糖激酶(HK)和葡萄糖-6-磷酸脱氢酶(G6PD)的活性,降低葡萄糖-6-磷酸酶(G6P)和果糖-1,6-二磷酸酯酶(fFBPase)的活性,来防止糖尿病大鼠的体重减轻,并提高肝组织中糖原含量[39]。且橙皮苷能降低TNF-α、前列腺素E2(PGE2)、血管内皮生长因子(VEGF)和MDA的水平,减弱空腹血糖、炎症、晶状体液体内流、晶状体渗透不平衡及纤维过度水合作用来减轻糖尿病及糖尿病性白内障[40]。橙皮苷还可通过增加脑源性神经营养因子(BDNF)及酪氨酸蛋白激酶受体B(TrkB)的表达来抑制视网膜神经节细胞的凋亡,即橙皮苷的抗凋亡和降糖作用与BDNF/TrkB信号通路有关[41]。柚皮素通过调节糖尿病大鼠的氧化—硝化应激、炎症因子(TNF-α、TGF-1β)的释放和抑制基质金属蛋白酶-9(MMP-9)活性来改善糖尿病神经性疼痛[42]。此外,在自发型糖尿病KK-Ay小鼠模型中,柑橘多甲氧基黄酮组分能显著降低糖尿病小鼠的血清谷丙转氨酶(ALT)的水平和甘油三酯(TG)的含量,提高高密度脂蛋白胆固醇(HDL-c)和脂联素(ADPN)的浓度以及谷草转氨酶/谷丙转氨酶(AST/ALT)的比值,有效改善糖尿病小鼠糖脂代谢[36]27-38。在高糖诱导的糖尿病大鼠中,柚皮素能够缓解高血糖诱导的肝脏中有机阳离子转运蛋白1(OCT1)表达的减少,并导致二甲双胍积累和乳酸生成增加[43]。因此,糖尿病患者在使用二甲双胍治疗时应避免摄入含橙皮素的果汁。
柑橘中黄酮类化合物具有显著抗糖尿病活性,其作用机制与激活PI3K/Akt、WNT等信号通路,抑制MMP-9活性,降低空腹血糖、ALT、TG,提高HDL-c、ADPN、AST/ALT的比值,改善BDNF、TrkB的表达等有关。不同柑橘黄酮类化合物的作用机制不同,但仍需进一步研究活性与化合物结构的关系。
2.6 抗癌
柑橘黄酮类化合物具有抗癌抗肿瘤活性。在细胞试验中,柑橘黄酮类化合物通过下调细胞周期蛋白B1、CDK1和CDC25C蛋白的表达水平,在G2/M期诱导人肺癌细胞A549细胞阻滞,从而抑制细胞增殖;并通过上调Bax/BcL-xL比例诱导半胱天冬酶依赖性细胞凋亡[44]。柑橘黄酮通过激活MAPKs和抑制PI3K/Akt通路诱导细胞周期阻滞和凋亡,并抑制Hep3B肝癌细胞的迁移[45]。源于葡萄柚的纯化黄酮类化合物能通过调控白血病细胞凋亡相关因子Mcl-1的表达,抑制白血病细胞增殖,诱导白血病细胞凋亡[46]。橙皮素通过提高线粒体途径凋亡相关蛋白的水平来诱导肺腺癌细胞PC9细胞凋亡并呈时间—剂量依赖性,且可能与内质网应激有关[47]。桔皮素可以抑制信号转导及转录活化因子3(Stat3)信号通路诱导肿瘤干细胞死亡[48]。在动物试验中,柑橘类黄酮柚皮素可抑制去卵巢肥胖小鼠的肿瘤生长[49]。在氧化偶氮甲烷/葡聚糖硫酸钠诱导的结肠炎致结肠癌小鼠模型中,摄入5-去甲基化川陈皮素能显著抑制小鼠结肠炎驱动的结肠癌变,该作用与其在结肠中的代谢物有关[50]。即柑橘黄酮类化合物可能通过激活MAPKs和抑制PI3K/Akt、Stat3信号通路,增加Mcl-1的表达等来诱导癌细胞的凋亡。
2.7 肠道保护
一些研究报道了柑橘黄酮对肠道的保护作用。在非甾体抗炎药(双氯芬酸)诱导的体内外肠道损伤模型中,纯柑橘总黄酮通过PI3K/Akt信号通路促进细胞自噬来保护肠道屏障的完整性[51]。在三硝基苯磺酸(TNBS)诱导的炎症性肠病大鼠中,口服125~500 mg/kg干柑橘能显著地缓解大鼠体重减轻和腹泻,减少结肠炎炎症细胞浸润,并抑制促炎因子的产生[20]。且柚皮素和川陈皮素对离体空肠收缩有抑制作用。柚皮素的作用机制部分与COX、NOS、肌醇三磷酸(IP3)有关,并最终降低空肠运动[20]。此外,柑橘类黄酮柚皮素、川陈皮素和橙皮素能缓解TNBS诱导的溃疡性结肠炎小鼠模型体重减轻和结肠缩短,降低疾病活动指数评分,显著上调密封蛋白-2、闭合蛋白和紧密连接蛋白-1(ZO-1)的表达[52]。同时,黄酮类化合物可增强经上皮电阻,降低LPS损伤的单层上皮系统的通透性,上调闭合蛋白和ZO-1的表达。此外,在体外肠道—血管屏障模型中,柚皮苷通过调节内皮细胞的连接、减少细胞凋亡水平、促进细胞迁移和修复内皮细胞屏障,来保护肠道微血管内皮细胞单层屏障的完整性,防止TNF-α诱导的破坏,有效地改善肠—血管屏障[53]。柑橘黄酮处理可以显著地改变肠道中多种与人体健康密切相关菌群的丰度,使其向有益人体健康方向调节。并且高剂量的橙皮苷和柚皮苷可以显著地增加能量代谢和短链脂肪酸合成相关基因的相对丰度,橙皮苷的作用效果强于柚皮苷[54]。因此,柚皮素、川陈皮素和橙皮素等柑橘类黄酮可能通过调节PI3K/Akt信号通路、保护肠道黏膜屏障完整性、调节肠道菌群及短链脂肪酸的合成等途径来改善炎症性肠病。
2.8 心血管保护
黄酮类化合物可以用于保护心脑血管。在RAW264.7细胞模型中,柑橘花中的高圣草酚和橙皮苷-7-O-β-d-吡喃葡萄糖苷通过调节PPARγ、磷脂ATP结合盒转运体A1(ABCA1)、磷脂ATP结合盒转运体G1(ABCG1)、清道夫受体B类1型(SRB1)、清道夫受体A类1型(SRA1)和分化簇36(CD36)的基因和蛋白表达水平来抑制LPS诱导的炎症反应和氧化低密度脂蛋白诱导的泡沫细胞形成,高圣草酚预防动脉粥样硬化的效果比橙皮苷-7-O-β-d-吡喃葡萄糖苷更强[55]。柑橘类黄酮柚皮素可以显著地激活去乙酰化酶(SIRT1)并具有抗衰老作用,能保护心肌免受衰老依赖性功能障碍[56]。
在动物模型中,川陈皮素可通过抑制氧化应激和基质金属蛋白酶(MMP-2和MMP-9)的水平来改善糖尿病大鼠心血管功能紊乱。在代谢综合征大鼠模型中,橙皮苷通过改变代谢综合征大鼠心脏和肾脏组织中不同的心血管风险生物标记物蛋白组学特征发挥其保护作用[57]。橘皮中的总黄酮可通过抗氧化和抗炎作用改善高脂喂养诱导的高血脂仓鼠的肝功能,发挥降血脂作用,其作用机制可能是通过增加PPARα和PPARγ的表达[58]。柑橘黄酮类化合物可能是通过激活SIRT1、PPARα和PPARγ,抑制氧化应激和MMP-2、MMP-9活性,从而发挥心血管保护作用。
2.9 肝脏保护
柑橘黄酮类化合物具有肝脏保护作用。在胆管结扎诱导小鼠肝纤维化模型中,富含多甲氧基黄酮类化合物的柑橘皮提取物具有抗氧化、抗炎、抗凋亡活性,能有效调节胆管结扎诱导的肝脏损伤。在镉诱导的肝脏损伤大鼠模型中,柑橘类黄酮香叶木苷能有效地降低MDA和一氧化氮(NO)含量,改善SOD、CAT、GPx活性,增加谷胱甘肽(GSH)含量[59]。因此,在可能接触镉的情况下,可将香叶木苷作为辅助剂与食物一起服用,以预防肝脏疾病的发生。此外,在酒精诱导的肝脏损伤小鼠模型中,来源于柑橘提取物的多甲氧基黄酮类化合物对酒精诱导的肝损伤具有保护作用,其机制与调节细胞色素P450 2E1(CYP2E1)介导的肝脏氧化应激有关[60]。而在非酒精性脂肪肝病大鼠模型中,柑橘总黄酮通过调节NF-κB和MAPKs信号通路发挥保肝抗炎作用[61]。在游离脂肪酸培养的人LX-2细胞和高脂饮食诱导的非酒精性脂肪肝病的雄性C57BL/6小鼠模型中,柑橘总黄酮上调Nrf2和下游靶基因的蛋白质表达,激活Nrf2-ARE信号通路,从而起到保肝作用[62]。在高脂饮食诱导的非酒精脂肪肝炎C57BL/6 J小鼠模型中,纯柑橘总黄酮通过调节小鼠肠道菌群和胆汁酸代谢改善非酒精性脂肪性肝炎。
2.10 神经保护作用
柑橘黄酮具有保护神经的作用。柑橘黄酮可能通过下调SNHGl表达,促进1-甲基-4-苯基-吡啶离子诱导的帕金森病细胞模型SH-SY5Y细胞增殖,抑制其凋亡[63]。此外,柑橘类黄酮3, 5, 6, 7, 8, 3′, 4′-七甲氧基黄酮能够改善皮质脂酮诱导的类似抑郁的行为,恢复短暂性全脑缺血小鼠模型的海马体BDNF的表达、神经形成和神经重塑[64]。柚皮素通过减少小鼠脑中淀粉样蛋白(Aβ)的产生、Tau过度磷酸化、氧化应激和神经炎症,改善衰老小鼠模型的认知缺陷[65]。柑橘黄酮具有神经保护作用,作用机制可能是下调SNHGl的表达、降低氧化应激和改善神经炎症以及BDNF的表达。柑橘黄酮是治疗帕金森病、阿尔茨海默症等神经性疾病的潜在成分,但仍需进一步的研究验证。
2.11 其他活性
柑橘黄酮类化合物除抗炎等作用外,还有一些其他生物活性,如抗过敏、抗抑郁和抗衰老等。在RBL-2H3细胞系中,柑橘黄酮减少了抗原诱导的β-己糖苷酶(HEX)的释放,且在卵蛋白诱导的过敏性哮喘小鼠模型中,经不同浓度的黄酮处理后,免疫球蛋白E(IgE)、白介素-4(IL-4)、白介素-13(IL-13)显著减少,即黄酮在体外和体内均具有抗过敏活性。在亚慢性轻度社会失败应激模型小鼠中,膳食摄入柑橘类黄酮橙皮苷能通过抑制犬尿氨酸(KYN)信号通路的增强来提高应激恢复能力[66]。柑橘黄酮能够上调促甲状腺因子Sirt1并不同程度地影响甲状腺Nrf2的表达,有助于恢复老年大鼠甲状腺功能受损[67]。
3 总结与展望
柑橘中含有多种黄酮类化合物,而用于从柑橘中提取黄酮类化合物的新技术,如低共熔溶剂提取、亚临界提取及磁性分子印迹选择性提取等高效、环境友好型提取技术,为柑橘黄酮类化合物的开发利用提供了便利。大量体外和体内试验研究表明柑橘黄酮具有抗氧化、抗炎、抗菌、抗肥胖、抗糖尿病、抗癌、肠道保护、心血管保护和神经保护作用等生物活性,其作用机制涉及多种信号通路,但缺乏对柑橘类黄酮化合物的结构与生物活性之间的关系的研究。因此,可利用网络药理学、分子对接和分子动力学等方法,研究柑橘黄酮类化合物相关活性的作用机制及作用靶点。同时对柑橘类黄酮在人体内的生物利用度需要进一步探讨。
猜你喜欢
家庭医药(2021年18期)2021-12-03
保健与生活(2021年13期)2021-07-14
世界科学技术-中医药现代化(2021年10期)2021-03-02
恋爱婚姻家庭(2019年30期)2019-03-23
天然产物研究与开发(2018年11期)2018-11-30
中成药(2018年9期)2018-10-09
益寿宝典(2017年2期)2017-02-26
恋爱婚姻家庭·养生版(2017年2期)2017-02-15
中国当代医药(2015年24期)2015-03-01
中国当代医药(2015年9期)2015-03-01
