
2018—2020年豫南地区猪流行性腹泻病毒的遗传变异分析
2022-11-15 06:20曲哲会张喜文董建国赵云焕郭晓秋许琬雪李盼盼李卓燕
中国兽医学报 2022年10期
曲哲会,张喜文,董建国,刘 涛,赵云焕,2,3,郭晓秋*,李 颖,许琬雪,李盼盼,李卓燕
(1.信阳农林学院 牧医工程学院,河南 信阳 464000;2.河南省大别山区生态畜禽健康生产工程研究中心,河南 信阳 464000;3.信阳市畜禽养殖与环境控制重点实验室,河南 信阳 464000)
猪流行性腹泻(porcine epidemic diarrhea,PED)是由猪流行性腹泻病毒(porcine epidemic diarrhea virus,PEDV)引起的一种急性、高度接触性传染病,以呕吐、水样腹泻及脱水为主要临床表现[1]。各种年龄猪群均可以被PEDV感染,新生仔猪发病尤为严重,死亡率可达50%~100%。该病1976年被首次报道[2],经典疫苗毒株CV777的广泛使用,该病得到有效控制[3]。但是,2010年我国多个省市地区发生严重的PEDV疫情,尤其是免疫CV777疫苗毒株的猪群也同样发生,给养殖场造成了巨大的经济损失[4-6]。同期,美国、韩国、日本等多个国家也同样发生该病的流行[7-9]。
PEDV属于冠状病毒科冠状病毒属的冠状病毒Ⅰ群成员。PEDV的基因组为RNA,约28 kb,共编码4种结构蛋白,即S蛋白、M蛋白、E蛋白和N蛋白;非结构蛋白ORF3和pp1ab(可裂解为16种非结构蛋白)[2]。S蛋白是PEDV的表面糖蛋白,在PEDV入侵宿主细胞及诱导机体产生中和抗体方面发挥重要作用[10]。S蛋白可分为S1蛋白(1~789 aa)和S2蛋白(790~1 383 aa),其中S1蛋白包括了主要中和抗原表位及受体结合区域。S蛋白在外界环境与条件压力下极易发生基因突变、插入与缺失,因此,对PEDV的S基因进行遗传变异分析对于掌握PEDV流行株的分子遗传特征具有十分重要的意义[11]。ORF3蛋白是PEDV的唯一辅助性蛋白,其基因极其不稳定,容易发生变异而导致基因多态性,已发现弱毒株常发生连续17 aa的缺失,也成为区别弱毒株和强毒株的重要标记。此外,ORF3蛋白可促进PEDV在感染细胞中增殖[12-13]。N蛋白是一种由大部分碱性氨基酸组成的磷酸化核衣壳蛋白,基因核苷酸序列相对稳定,对病毒的组装与复制起着重要作用[14]。
为掌握2018—2020年豫南地区流行的PEDV的分子遗传特征,本研究从18个规模化猪场采集PEDV感染仔猪组织扩增出S1基因部分片段、ORF3基因和N基因,并进行核苷酸序列测定,通过与已发表参考序列进行氨基酸进化树构建、相似性及变异分析,可为该地区PEDV的防控提供理论指导。
1 材料与方法
1.1 病料PEDV感染的仔猪肠道、内容物和肠系膜淋巴结,采自2018—2020年间豫南地区18个规模猪场,由本实验室保存。
1.2 试剂病毒基因组DNA/RNA提取试剂盒(离心柱型)购自天根生化科技(北京)有限公司;反转录试剂盒购自Thermo公司;2×Taq Master Mix购自上海惠凌生物技术有限公司;DL2000 DNA Marker、50×TAE核酸电泳液购自生工生物工程(上海)股份有限公司;琼脂糖购自广州赛国生物科技有限公司;氯化钠、氯仿、无水乙醇均为国产分析纯。
1.3 引物设计与合成参照文献[15],直接合成可用于扩增PEDV的S1基因部分片段(编码S氨基酸464~792片段)和包含ORF3基因的引物。根据PEDV YZ株(MK841495.1)的核苷酸序列作为参考序列,利用软件Primmer 5.0设计N基因扩增引物,具体引物序列见表1。上述引物均由生工生物工程(上海)股份有限公司合成。

表1 PCR扩增引物序列
1.4 病料的处理将从18个猪场采集的仔猪小肠、内容物和肠系膜淋巴结剪碎后研磨,用无菌生理盐水按照1∶5稀释,反复冻融3次后, 4℃、12 000 r/min离心10 min,取上清液。
1.5 RNA提取与反转录利用DNA/RNA提取试剂盒从处理后的样品中提取基因组RNA,具体过程按照说明书进行。以提取的RNA为模板,利用RNA反转录试剂盒合成cDNA。反转录体系(20 μL):RI 1 μL,RT 1 μL,Re Buffer 4 μL,dNTP 2 μL,OligodT181 μL,RNA 11 μL。反转录程序:42℃ 60 min,70℃ 5 min,冰浴5 min,-20℃保存备用。
1.6 PEDV的S、ORF3和N基因扩增与测序以合成cDNA为模板,分别扩增PEDV的S1基因部分片段、包含有ORF3基因片段和N基因。PCR反应体系(50 μL):2×Taq MasterMix 25.0 μL,引物S1-S(或ORF3-S、N-S) 1.0 μL,引物S1-A(或ORF3-A、N-A) 1.0 μL,cDNA 4.0 μL,ddH2O 19.0 μL。PCR反应程序:95℃预变性5 min;95℃变性 30 s,52℃退火 30 s,72℃延伸1 min 30 s,共35个循环;72℃延伸7 min。PCR产物送至生工生物工程(上海)股份有限公司进行核苷酸序列测定。
1.7 PEDV的S、ORF3和N基因序列分析利用生物学软件MegAlign进行PEDV流行株 S1、ORF3和N基因推到氨基酸序列与15株参考序列的相似性分析。利用软件Mega 7.0的Boot-strapped Neighbor-Joining算法构建进化树及氨基酸变异分析。
2 结果
2.1 PEDV的S、ORF3和N基因PCR扩增结果利用RT-PCR方法对18个猪场PEDV感染仔猪组织中扩增S、ORF3和N基因。结果表明,来自18个猪场样品中均获得与预期相符的PCR产物,其中引物S1/S2的PCR产物约990 bp,引物ORF3-S/ORF3-A的PCR产物约为930 bp,引物N-S/N-A的PCR产物约为1 400 bp。经核苷酸序列测定结果证实,18个猪场流行的PEDV主要有5株,分别命名为HNHB、HNNY、HNZY、HNHC和HNXC。
2.2 PEDV的S1基因部分序列遗传变异分析PEDV的S1基因部分序列构建进化树如图1所示,5株PEDV流行株和15株参考序列构建进化树分成2个群,即G1和G2。5株PEDV流行株均属于G2a分支,相似性为96.4%~100%;与同分支参考序列相似性为96.4%~98.5%;与G2b分支参考序列相似性为95.1%~97.0%;与G1分支参考序列相似性为93.0%~96.7%。如表2所示,与G1群中经典毒株CV777相比较,5株PEDV流行株S氨基酸序列在10个位置发生相同氨基酸置换,分别为A517S、S523G、V527I、G549S、A605E、I635V、N707D、N724S、Y766S;此外,流行株HNHB、HNNY和HNZY在6个位置发生相同氨基酸置换,即G520D、L521H、Q574H、K621T、I667F、A704V;流行毒株HNHC存在7个位置发生氨基酸置换,即I496S、S522N、K563N、L612F、M641I、N719S、Q765H;流行毒株HNXC存在5个位置发生氨基酸置换,即L521R、F536L、L612F、N719S、Q765H发生氨基酸突变。
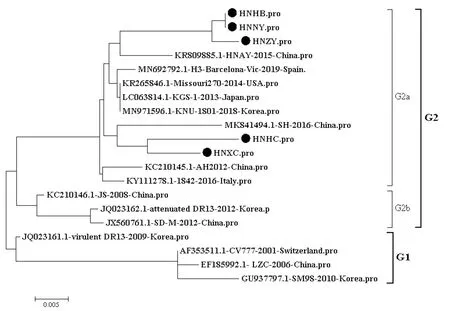
图1 PEDV S1部分基因推导氨基酸进化树分析
2.3 PEDV的ORF3基因推导氨基酸序列遗传变异分析PEDV的ORF3基因推导氨基酸序列构建进化树如图2所示,5株PEDV流行株与参考毒株序列可分成2个群,即G1和G2群。HNXC株和HNHC株属于G2a群,与G1群参考序列相似性均为92.4%;与G2b群参考序列相似性为94.7%~96.4%;与G2c群流行毒株序列相似性为95.6%~96.4%。HNHB株、HNNY株和HNZY株都属于G2c群,相似性为99.6%~100%;与G1群参考序列相似性为88.0%~89.1%;与G2a群序列相似性为92.4%~94.8%;与G2b群序列相似性为94.7~96.9%。G1群PEDV毒株均为弱毒株,ORF3均不同程度出现氨基酸序列片段的缺失,与其相比较,5株PEDV流行株均为编码224个氨基酸的完整序列,不存在氨基酸的缺失,具有强毒株的分子遗传特征。如表3所示,与经典毒株CV777相比较,5株PEDV流行株在5个位置发生了相同氨基酸的置换,分别为V21A、V54I、V79I、A101T和N166S;PEDV流行株HNNY、HNZY和HNHB在7个位置发生相同氨基酸的置换,分别为L25S、S63A、I70V、L81I、L92F、C107F和D168N;HNXC和HNHC在2个位置发生相同氨基酸的置换,分别为F80V和H182Q;此外HNZY在41位发生氨基酸置换(A41T),HNXC和HNHC均在92位氨基酸发生置换,分别为L92F和L92I。

表2 PEDV S1部分基因推导氨基酸与经典毒株CV777比较发生氨基酸置换
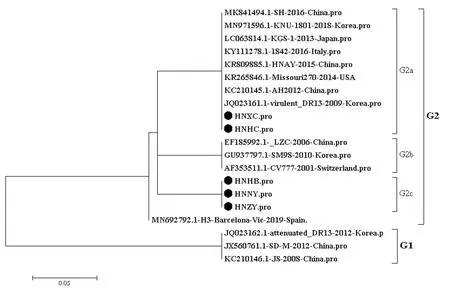
图2 PEDV ORF3氨基酸进化树分析
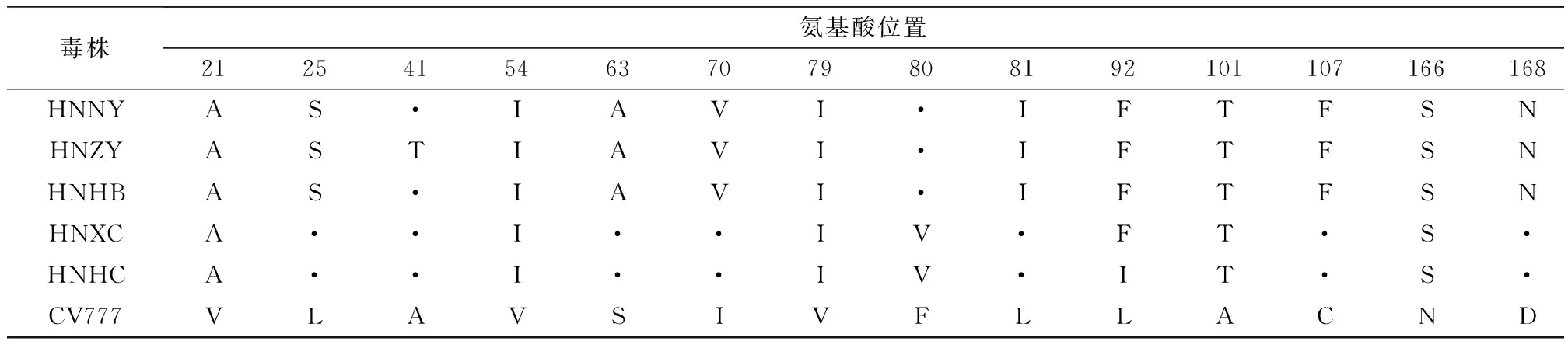
表3 PEDV ORF3基因推导氨基酸与经典毒株CV777比较发生氨基酸置换
2.4 PEDV N基因推导氨基酸序列遗传变异分析PEDV的N基因推导氨基酸序列构建进化树如图3所示,5株PEDV流行株和参考序列共形成2个群,2010年之前流行株(包括经典疫苗毒株CV777)和SD-M株、DR13株组成G1群;5株流行株与2010年以后的流行株组成G2群,其中HNZY、HNHB与参考株属于G2a亚群,HNXC株、HNHC株和HNNY株属于G2b亚群。流行株HNZY和HNHB与G1群毒株相似性为95.7%~96.8%;与G2b亚群相似性为97.1%~98.9%;与G2a亚群其他毒株相似性为98.4%~99.3%。流行株HNXC、HNHC和HNNY与G1群毒株相似性为95.0%~96.8%;与G1亚群内毒株相似性为97.1%~98.9%。如表4所示,与经典毒株CV777的N氨基酸序列相比较,5株PEDV流行株在10个位置发生了相同氨基酸的置换,即G84A、K123N、A142T、N205K、H242L、L381P、L395Q、Q397L、H398N、E400D;HNHB、HNZY、HNHC和HNNY株在4个位置发生相同氨基酸置换,即R241K、K252R、A408L和V412S,而HNXC仅在408位置氨基酸发生置换(A408V);HNXC、HNHC和HNNY株在4个位置发生相同氨基酸置换,即F309I、V910C、E311R、G313S;此外,HNHB株在2个位置发生氨基酸置换(N255I和N258Y),HNZY株在3个位置发生氨基酸置换(D240E、N255S和P285L),HNXC株在1个位置发生氨基酸置换(N416Y),HNHC株在2个位置发生氨基酸置换(N255S和P409S),HNNY存在2个位置氨基酸发生置换(D240E和N255S)。

图3 PEDV N氨基酸进化树分析

表4 PEDV N基因推导氨基酸与经典毒株CV777比较发生氨基酸置换
3 讨论
自从2010年开始,我国大部分地区暴发PEDV疫情,许多猪场的新生仔猪的病死率接近100%,造成了严重的经济损失[6]。经过许多研究结果证实,与经典疫苗毒株CV777相比, 2010年后国内流行的PEDV株出现氨基酸突变、插入及缺失等形式变异,从而导致疫苗免疫猪仍然可以被感染。虽然PEDV仅有一个血清型,但在疫苗免疫、环境和药物作用等压力因素下,PEDV在流行过程中极有可能发生变异。因此,及时掌握猪群中流行的PEDV的遗传变异情况对于该病的防控具有重要的指导意义。本研究扩增5株豫南地区的PEDV流行株S1部分基因、ORF3基因和N基因,并进行核苷酸序列测定,通过对其推到氨基酸序列的分子遗传变异分析,可为该地区制定科学的PEDV综合防控措施提供指导。
PEDV的S蛋白是由1 383个氨基酸(aa)组成,能促进病毒与宿主细胞的融合,刺激宿主细胞产生中和抗体等。已有研究证实,S蛋白具有4个中和表位,分别为COE(499~638 aa)、SS2(748~755 aa)、SS6(764~771 aa)和2C10(1 368~1 374 aa)[16],其中S1部分包含有3个中和表位(COE、SS2和SS6),S2部分有1个中和表位(2C10)。已有研究证实,PEDV变异毒株S蛋白氨基酸序列存在氨基酸片段的插入、缺失及突变等变异方式,是导致疫苗免疫失败的重要原因[17-21]。本研究中,根据S1部分氨基酸序列进化树分析,5个PEDV流行株分布在G2a群,与CV777为代表的2010年前流行毒株组成的G1群亲缘关系较远。氨基酸变异方面,与CV777毒株相比较,5个流行株在COE区域存在7个相同氨基酸的置换,在SS6区域存在1个相同氨基酸的置换,此外,每个PEDV流行株在不同位置氨基酸发生置换,表现出不同毒株变异的多样性。上述结果表明,本地区流行的PEDV毒株均为变异毒株,且与疫苗毒株CV777存在抗原位点氨基酸变异,推测其实导致疫苗免疫效果不佳的一个重要原因。
ORF3是PEDV重要的非结构蛋白,对PEDV适应体外培养宿主细胞发挥重要作用。已有报道证实,ORF3是否发生17个氨基酸缺失是区别野生型和弱毒株的关键分子特征,其与病毒的毒力与致病性有密切关系。本研究中,基于ORF3氨基酸序列进化树分析结果表明,5株PEDV流行株与CV777均为属于G2群,但分布在不同亚群,编码224个氨基酸,与3个ORF3氨基酸缺失株组成G1群(JQ023162、JX560761和KC210146)亲缘关系较远。与疫苗毒株CV777相比,5株流行株存在5个相同氨基酸置换位点,与周兵强等[22]报道结果相一致。此外,还存在不同程度的单个氨基酸突变位点。上述分析结果表明,5株PEDV流行株具有野生型毒株的分子遗传特征,并表现出多样性变异的特点,与国内报道相一致[23-25]。
相比较其他蛋白,PEDV N蛋白相对比较保守,其在病毒基因组RNA复制、诱导机体特异性免疫反应及协助病毒逃避免疫反应方面发挥重要作用。董波等[26]报道,8株闽西地区PEDV流行株与CV777亲缘关系较远,N蛋白氨基酸序列存在14个位点相同氨基酸置换。本研究中,5株流行株与2010年后流行参考毒株组成G2群,与疫苗毒株CV777所在G1群亲缘关系较远,结果与上述报道相一致。5株流行株存在10个相同氨基酸置换位点,其中有9个位置氨基酸变异与董波等[26]报道相一致,有1个位点不相同(H398N)。此外,闽西地区PEDV流行株还存在5个氨基酸位点的变异,而在豫南地区流行株中未发生相应位置氨基酸变异,但在其他位置发生多个位点不同氨基酸位点的突变,呈现出分子遗传多样性特征。
猜你喜欢
数学物理学报(2022年5期)2022-10-09
科学大观园(2022年2期)2022-01-23
天津市教科院学报(2021年5期)2021-11-10
生物学通报(2021年9期)2021-07-01
河北画报(2020年8期)2020-10-27
中国猪业(2017年11期)2017-12-11
中国医学创新(2017年7期)2017-03-31
江苏农业科学(2016年8期)2017-02-15
浙江大学学报(工学版)(2016年2期)2016-06-05
特产研究(2014年4期)2014-04-10
