
细胞焦亡在非小细胞肺癌中的作用及机制研究进展
2022-11-10 06:12:32张钰哲刘虹汝王甲正
昆明理工大学学报(自然科学版) 2022年5期
张钰哲,刘虹汝,袁 靓,王甲正
(1.大理大学 基础医学院,云南 大理 671000; 2.云南省滇西抗病原植物资源筛选研究重点实验室,云南 大理 671000; 3.云南省李云庆专家工作站,云南 大理 671000)
0 引 言
非小细胞肺癌(Non-Small-Cell Carcinoma,NSCLC) 是肺癌中最常见的组织学亚型,约占肺癌总数的80%~85%,位居我国男性恶性肿瘤第一位[1].2021年,全球新发肺癌病约235万例,其中死亡病例约有131万[2].因非小细胞肺癌较高的侵袭性等病理学特点,及缺乏明确有效的早期筛查方案,使我国多数非小细胞肺癌患者确诊时已是晚期,失去最佳手术治疗时机.因此,深入研究非小细胞肺癌的发生发展及诊疗的新策略十分重要.
细胞死亡主要分为由细胞凋亡、细胞焦亡等构成的程序性死亡和由细胞坏死等构成的非程序性死亡两大类.细胞焦亡(Pyroptosis)是依赖于半胱天冬氨酸蛋白酶-1(Caspase-1)介导的程序性细胞死亡方式,涉及Caspase-1介导的典型信号途径和Caspase-4、Caspase-5、Caspase-11介导的非经典信号途径[3].目前有研究直接或间接表明细胞焦亡为肝癌[4-6]、胃癌[7-8]、前列腺癌[9]、乳腺癌[10-12]、皮肤癌[13]等相关疾病的诊断与治疗提供了重要依据.近年来研究发现,细胞焦亡可通过多种途径参与非小细胞肺癌的发展过程,提示其可能成为非小细胞肺癌诊治的重要靶点.
本文将从细胞焦亡的角度综述当前细胞焦亡在非小细胞肺癌发生发展的最新研究进展,为非小细胞肺癌的治疗及预后提供依据.
1 细胞焦亡综述
1.1 细胞焦亡
细胞焦亡是一种由Caspase激活、炎性因子释放及GSDMD (Gasdermin D)裂解而诱发炎症发生的程序性细胞死亡方式[14-16].2001年,Cookson等[15]提出了细胞焦亡的概念,来自希腊语.“pyro”术语意为“发烧”;“ptosis”术语意为“跌倒、落下”,表明细胞焦亡是由炎症反应引起的细胞程序性死亡.2015年,Shi等[17]发现,细胞焦亡由革兰阴性菌内毒素脂多糖(Lipopolysaccharide,LPS)激活Caspase-4/Caspase-5/Caspase-11,活化的Caspase-4/Caspase-5/ Caspase-11对Gasdermin家族蛋白进行切割最终诱发细胞焦亡发生.2017年,Shi等[18]将细胞焦亡的定义重新修订为由Gasdermin家族蛋白介导的程序性细胞坏死.2018年,Galluzzi等[19]将细胞焦亡的定义再次修订为细胞焦亡是由Caspase激活且依赖于Gasdermin蛋白家族成员形成细胞质膜膜孔的可调控细胞死亡的炎症性活化结果.
1.2 细胞焦亡的形态学特征
细胞发生焦亡时细胞渗透性肿胀破裂且细胞膜失去完整性,这一形态学特征与其他形式的细胞死亡如坏死性凋亡、细胞坏死、细胞凋亡有着显著区别[20](图1和表1).虽然细胞焦亡与细胞凋亡都被认为是一种程序性细胞死亡形式,两者在诸多方面存在相关性:如在PS曝光、膜联蛋白V染色、TUNEL染色、DNA损伤、染色质凝聚、膜起泡、热解体和凋亡体直径(1~5μm)、Caspase-3/Caspase-6/Caspase-8/Caspase-9 激活途径中细胞凋亡与细胞焦亡十分相似[21-26];但在炎症、焦亡体、完整的细胞核、孔隙形成、细胞肿胀、渗透裂解、7-AAD染色、Caspase-1/Caspase-4/Caspase-5/Caspase-11激活、Gasdermin裂解的途径中二者则存在明显差异[18, 27-30].
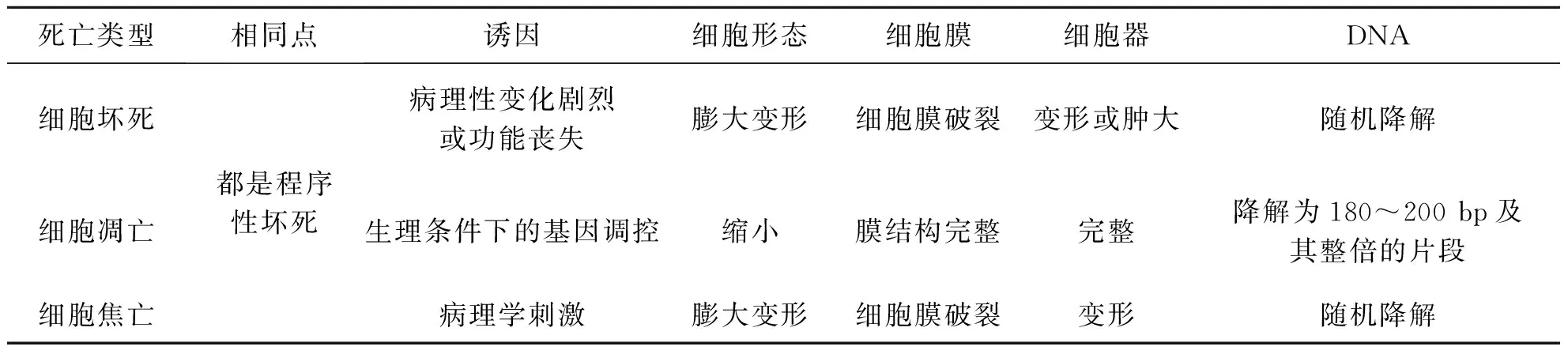
表1 不同细胞死亡形式的差异及相同点
1.3 细胞焦亡的分子途径
细胞焦亡的发生包括经典的炎症小体通路和非经典的炎症小体通路[28, 31].其中,经典的焦亡炎症小体通路又称为Caspase-1依赖途径,Caspase-1的激活是由炎症小体介导的,经典的焦亡炎症小体由胞质模式识别受体蛋白(Pattern Recognition Receptor,PRR),包括核苷酸结合寡聚化结构域样受体(Nucleotide- binding domain Leucine-rich Repeats,NLR)家族(NLRP1、NLRP3、NLRC4和AIM2)和PYHIN(PYRIN和HIN结构域蛋白)家族]、接头蛋白(ASC)和效应体半胱天冬酶-1前体(Pro-Caspase-1)三部分组成[32].炎症小体可与Caspase-1结合,将Caspase-1前体裂解成具有催化活性的Caspase-1,活化的Caspase-1切割焦亡执行关键蛋白GSDMD,切割产生的GSDMD-N在细胞包膜形成孔隙并破坏细胞内外的渗透压平衡,使包膜上离子失衡,导致细胞肿胀、裂解并死亡[33]. 此外, 活化的Caspase-1也可切割并加工IL-1β前体、IL-18前体,并将促炎症形式的成熟体IL-1β和IL-18分泌至细胞外,介导细胞焦亡发生[34](图2).
非经典的焦亡炎症小体通路又称为非Caspase-1依赖的细胞死亡方式,其主要由Caspase-4、Caspase-5、Caspase-11介导,不仅通过由胞质中LPS激活Caspase-4、Caspase-5、Caspase-11后切割GSDMD-N 端引起的细胞焦亡,还可通过刺激缝隙连接蛋白-1(Pannexin-1)通道开放,释放ATP后致K+外排后启动细胞焦亡进程[35].同时,也可通过 NLRP3/ Caspase-1途径介导IL-1β和IL-18的成熟和分泌[36-37].除了以上两种焦亡途径之外,Wang[24]和Sarhan等[38]还发现Caspase-3、Caspase-8也可介导细胞焦亡的发生(图2).
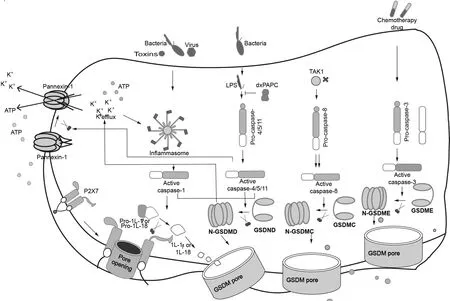
图2 细胞焦亡的分子途径Fig.2 Molecular pathways of pyroptosis
2 细胞焦亡与肿瘤疾病
研究表明,细胞焦亡可通过抑制肿瘤细胞的增殖、迁移和侵袭来治疗肿瘤,延缓病情进展.在恶性肿瘤等相关疾病的发生发展中发挥极其重要的作用[39-42].石新丽等[43]发现,在SMMC-7721肝癌细胞中加入鼠双微2(MDM2)拮抗剂可活化Caspase-1,并激活抑癌基因p53诱导肝癌细胞发生焦亡,发挥抗肝癌作用.Tang等[44]研究表明,雌激素的高低可调控MAPK通路,进而影响NLRP3炎症小体的表达,诱导乳腺癌细胞焦亡.此外,有研究发现,宫颈癌HeLa细胞被沙眼衣原体感染后,可导致胞内K+外流,活性氧ROS水平升高,从而使NLRP3炎性小体激活Caspase-1诱导宫颈癌细胞焦亡[45].上述研究证明了细胞焦亡与肿瘤的相关性.因此,深入探究与细胞焦亡密切相关的Caspase家族蛋白、GSDM家族蛋白、炎症小体及相关信号通路中的作用机理,对于我们深刻理解非小细胞肺癌发生发展的机制是十分必要的.
3 细胞焦亡与非小细胞肺癌
3.1 细胞焦亡与非小细胞肺癌研究进展
随着研究的不断展开和深入,人们发现细胞焦亡与非小细胞肺癌的进展存在一定的关系.GAO等[46]研究表明,在非小细胞肺癌中GSDMD呈现高表达与其侵袭性特征有关,当沉默非小细胞肺癌中GSDMD时,表皮生长因子受体信号减少、Caspase-3分解增加,并通过线粒体途径促进非小细胞肺癌细胞焦亡,从而可抑制肿瘤生长.Wang等[47]研究指出,辛伐他汀可通过激活过激活 NLRP3后使Caspase-1激活,从而介导经典途径诱导非小细胞肺癌细胞发生焦亡.Xi等[48]发现,GSDMD有助于肺鳞状细胞癌和肺腺癌中细胞毒性T淋巴细胞的杀伤.Lu等[49]研究不仅发现敲除DFNA5/GSDME可通过Caspase-3诱导非小细胞肺癌细胞从凋亡到焦亡的转换,还发现DFNA5/GSDME基因的缺失可促进非小细胞肺癌的耐药性,而DFNA5/GSDME过表达时会导致药物敏感性增加.大量研究表明,Caspase家族蛋白、GSDM家族蛋白、炎症小体等相关信号通路介导的细胞焦亡在非小细胞肺癌的发生发展中起着重要作用.
此外,细胞焦亡既能抑制肿瘤细胞增殖,又能提供肿瘤细胞生长的适宜环境,从而促进肿瘤细胞增殖.在非小细胞肺癌的进展中,细胞焦亡同样扮演着双重角色的作用,Liu等[42]发现,下调LncRNA-XIST致癌基因时,可激活NLRP3炎性小体,使Caspase-1、IL-1β表达增加,进而诱导肿瘤细胞焦亡,最终抑制非小细胞肺癌的发生发展,敲除LncRNA-XIST可使miR-335/SOD2/ROS信号通路中ROS的水平增高,进一步诱导细胞焦亡产生抗肿瘤的作用;而LncRNA-XIST过表达时则可促进非小细胞肺癌细胞增殖.Gao等[46]研究表明,硫氧还蛋白互作蛋白(thioredoxin-in-teracting protein,TXNIP)与NLRP3结合,通过TXNIP的过表达启动细胞焦亡,促进肺相关慢性炎症反应;而抑制TXNIP的表达时则可减少炎症小体的形成,抑制肺部相关慢性炎症反应.Wang等[47]研究证明,NLRP3炎症体的激活能够通过Caspase-1依赖性或非依赖性焦亡途径介导IL-18和IL-1β的释放来促进肺腺癌A549 细胞的增殖和迁移;而阻断IL-18和IL-1β信号传导,可减轻NLRP3炎性体对肺腺癌 A549 细胞增殖和迁移的影响,进而抑制肺癌的进展.因此,细胞焦亡相关因子有促进和抑制非小细胞肺癌发生发展的双重作用机制,但目前与之相关的研究相对较少,这可作为新的研究方向去深入探讨细胞焦亡在非小细胞肺癌中的双重作用,为寻求治疗靶点奠定基础(图3).
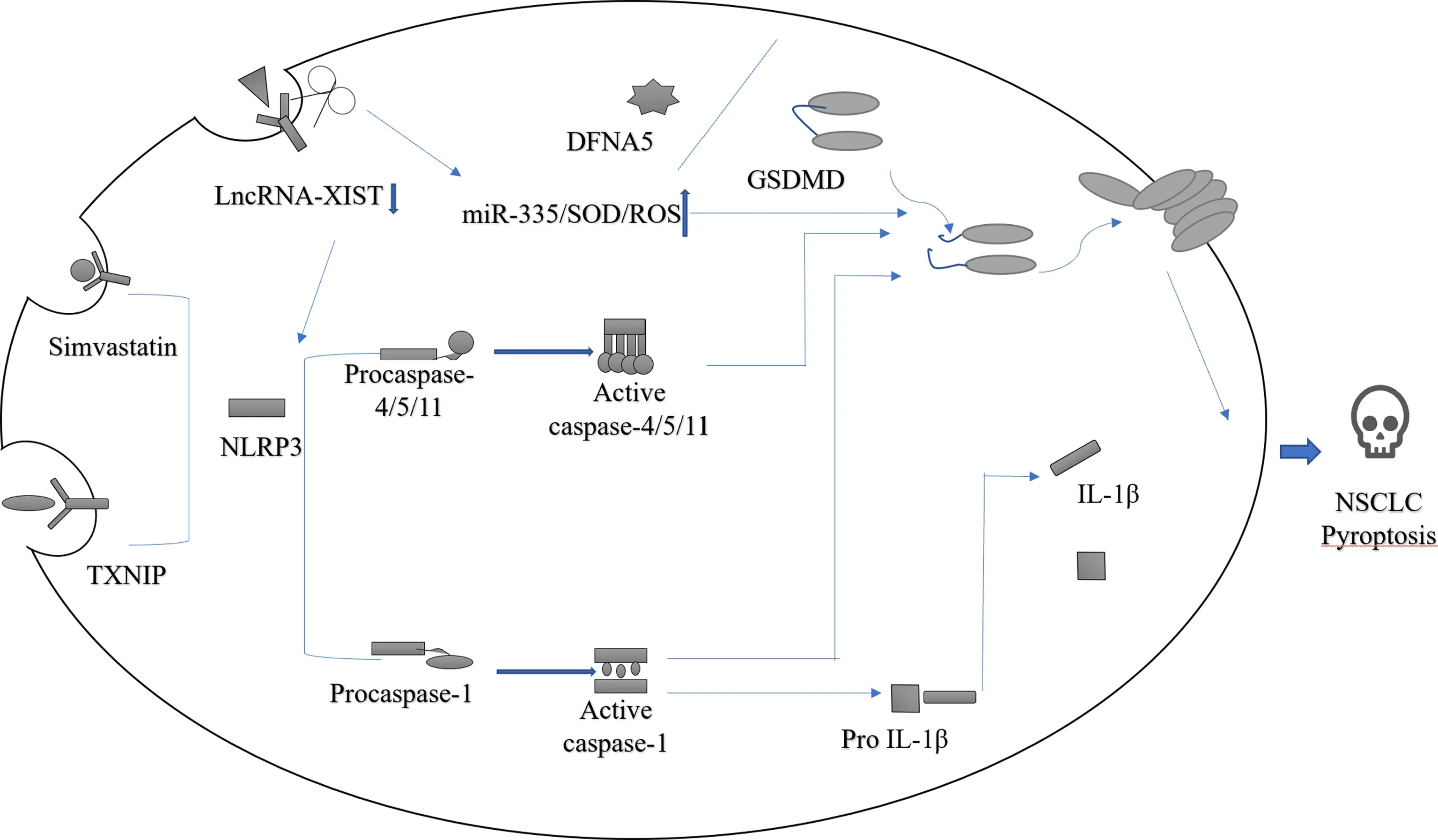
图3 细胞焦亡与非小细胞肺癌相关信号通路图Fig.3 Pyroapoptosis versus non-small cell lung cancer-related signaling pathway map
3.2 基于细胞焦亡的非小细胞肺癌早期诊断
目前非小细胞肺癌早期诊断的研究热点聚焦于细胞焦亡途径的相关分子,如:Caspase家族、Gasdermin家族、炎性小体、转录因子NF-κB和线粒体等相关蛋白.
3.2.1 Caspase-1及Gasdermin家族与非小细胞肺癌早期诊断
半胱氨酸裂解的Caspase-1不仅会导致细胞死亡,还会裂解促炎细胞因子,如IL-1β和 IL-18以及GSDMD[33,50].白花蛇舌草加半枝莲通过激活NLRP3/NF-κB/MAPK信号通路诱导细胞焦亡进而抑制非小细胞肺癌细胞的增殖和迁移[51].顺铂通过激活Caspase-3/GSDME诱导非小细胞肺癌A549细胞焦亡[33].因此,Caspase-1及GSDMD的激活在诱导细胞焦亡方面起着重要作用,可作为非小细胞肺癌的早期诊断的重要标记物.
3.2.2 NLRP3与非小细胞肺癌早期诊断
炎症小体NLRP3不仅可被多种病原体及致病分子识别激活[52],也可通过经典细胞焦亡途径来激活Caspase,并释放促炎因子IL-1β诱导细胞焦亡.Ma等[53]发现,红景天苷 (SAL)通过AMP活化蛋白激酶 (AMPK)[adenosine 5′-monophosphate (AMP)- activated protein kinase,AMPK]依赖性抑制NLRP3炎性小体进而诱导细胞焦亡发生,抑制肺癌细胞A549增殖和迁移.而杜卫华等[54]指出,NLRP3过表达时非小细胞肺癌A549细胞的增殖、迁移和侵袭能力也随之增强.另外,Wang等[55]发现,炎性小体AIM2不仅作用于固有免疫细胞,还可在平滑肌细胞及血管内皮细胞等细胞中产生应答激活,其羧基末端的HIN-200结构域可与DNA结合发生构象变化,激活Caspase-1的活性,释放下游炎性因子IL-1β和IL-18,最终诱导细胞焦亡.Yu等[56]研究表明,敲低AIM2的表达,可促使非小细胞肺癌A549细胞和NCI-H460细胞中Caspase-1激活和IL-1β成熟,降低G2/M期的细胞活性,抑制A549和NCI-H460细胞的迁移和侵袭能力,而AIM2过表达时则作用相反.此外,Zhang等[57]证明,上调AIM2的表达时,AIM2将以炎性依赖方式通过AIM2/IL-1β/STAT3 信号通路促进非小细胞肺癌细胞的生长.由此推断,调控焦亡的炎症小体NLRP3和AIM2可能成为非小细胞肺癌早期诊断的新靶标或潜在调节因子.
3.2.3 NF-kB与非小细胞肺癌早期诊断
转录因子NF-κB的激活与癌症的发展密切相关.Resibufogenin等药物可通过ROS介导的NF-κB抑制Caspase-1依赖性细胞焦亡的发生,延缓非小细胞肺癌病程进展[58].据报道,Polyphyllin VI和Casticin 可激活NF-κB和Caspase-1依赖性细胞焦亡的发生[59].说明转录因子NF-κB不仅有抗癌作用,还可激活细胞焦亡促进肿瘤的生长和侵袭.因此,调节NF-κB活性可能成为非小细胞肺癌早期诊断的新靶标[60-60].
3.2.4 线粒体与非小细胞肺癌早期诊断
Huang等[62]研究发现, LPS刺激内皮细胞中GSDMD的裂解将导致 GSDMD-NT在线粒体上的浓度改变,推测可能会穿透线粒体膜,线粒体上的GSDMD-NT孔形成将mtDNA释放到胞质中,可被细胞质DNA传感器cGAS检测到.在与mtDNA结合后,cGAS产生二级信使cGAMP,由接头蛋白STING检测.通过cGAMP结合激活STING抑制了YAP1易位进入细胞核,阻止了促进细胞增殖的基因的转录上调.另有研究指出,致病性耶尔森氏菌(Yersinia)不仅可抑制线粒体活化蛋白激酶(MAPK),也可被TLR4识别,TLR4通过RIP1发出信号,形成由 RIP1、Caspase-8和FADD组成的细胞死亡复合物,通过Caspase11驱动GsdmD切割和激活进一步导致细胞焦亡[38].此外,线粒体醛脱氢酶2(ALDH2)过表达可减少活性氧物种(ROS)的产生,并通过抑制细胞中NLRP3激酶和 Caspase1的激活来缓解高糖引起的细胞焦亡[63](图4).以上研究表明,线粒体可诱导细胞焦亡,将为确定非小细胞肺癌早期诊断的新靶标带来潜在的研究价值.
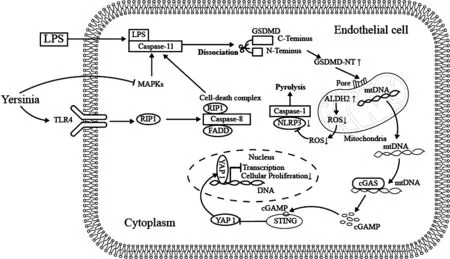
图4 线粒体诱导细胞焦亡途径Fig.4 The pathway of Mitochondria-induced pyroptosis
综上,细胞焦亡途径中的Caspase家族、Gasdermin家族、NLRP3和AIM2炎性小体、转录因子NF-κB和线粒体等相关蛋白有望成为非小细胞肺癌的早期诊断的有效切入点.
3.3 基于细胞焦亡的非小细胞肺癌药物治疗
基于文献检索,将细胞焦亡激活途径中所涉及的Caspase-1、NLRP3、GSDMD、IL-1β、IL-18和 κB等分子靶点在非小细胞肺癌药物治疗中的作用进行分析(表2).
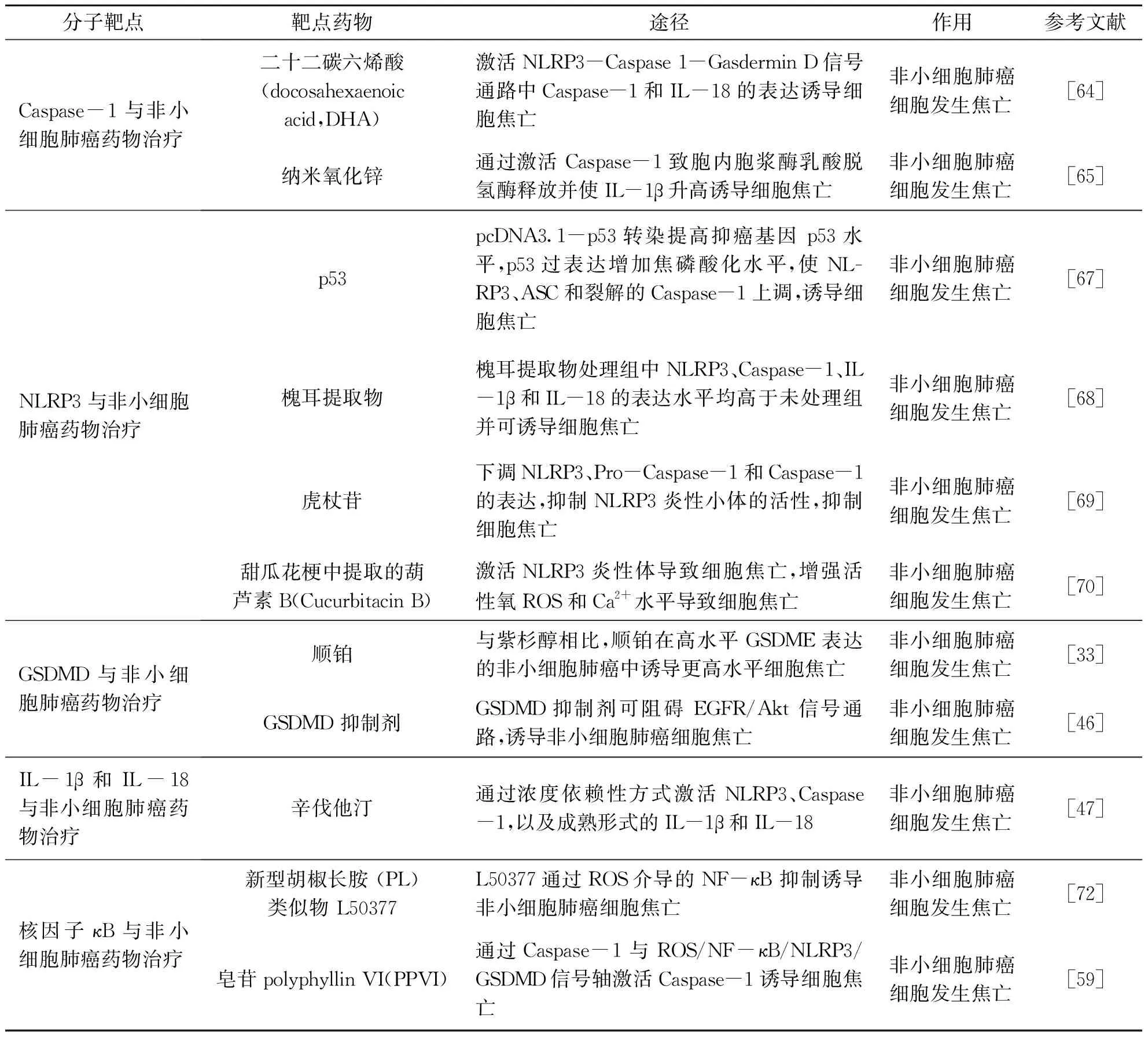
表2 细胞焦亡相关靶点及其机制与非小细胞肺癌药物治疗的关系
3.3.1 Caspase-1 与非小细胞肺癌药物治疗
Caspases是一组存在于细胞质中具有类似结构的蛋白酶家族,参与先天免疫应答、调节细胞死亡和炎症反应[29].黄辉[64]发现,增加二十二碳六烯酸(Docosahexaenoic acid,DHA)可激活NLRP3-Caspase-1 -Gasdermin D 信号通路中Caspase-1和IL-18的表达,诱导非小细胞肺癌细胞发生焦亡,抑制肿瘤的进展.Song等[65]观察到,纳米氧化锌与非小细胞肺癌A549 细胞接触后,通过激活Caspase-1致胞内胞浆酶乳酸脱氢酶释放并使IL-1β升高,诱导非小细胞肺癌细胞发生焦亡.说明Caspase-1参与非小细胞肺癌中的炎症反应,并促进了细胞发生焦亡.以上研究提示,Caspase-1在细胞焦亡激活途径中起着调控炎症小体的组成和下游相关基因的重要作用,可作为非小细胞肺癌临床诊断与治疗的的靶点.
3.3.2 NLRP3 与非小细胞肺癌药物治疗
NLRP3炎性小体是固有免疫的重要组分,在多种疾病过程中都发挥了关键作用[66].Zhang等[67]发现,通过pcDNA3.1-p53转染提高抑癌基因p53水平,p53过表达增加焦磷酸化水平,使mRNA和蛋白质水平上的NLRP3、ASC和裂解的Caspase-1上调,促使非小细胞肺癌细胞焦亡.Xie等[68]的研究表明,槐耳提取物处理的肺癌H520细胞和H358细胞中NLRP3、Caspase-1、IL-1β和IL-18的表达水平均高于未处理组,并可诱导非小细胞肺癌细胞焦亡.Zou等[69]研究了虎杖苷以剂量依赖性方式下调非小细胞肺癌细胞中NLRP3、Pro- Caspase-1和Caspase-1的表达,通过抑制NLRP3炎性小体的活性诱导细胞焦亡来抑制非小细胞肺癌细胞的增殖和转移.Yuan等[70]发现,甜瓜花梗中提取的葫芦素B(Cucurbitacin B)不仅与Toll 样受体4 (TLR4) 结合以激活NLRP3炎性体导致肿瘤细胞焦亡,也可增强非小细胞肺癌肿瘤细胞中线粒体活性氧(ROS)的产生和Ca2+导致非小细胞肺癌细胞焦亡.因此,NLRP3炎性小体可能成为非小细胞肺癌治疗的新策略.
3.3.3 GSDMD与非小细胞肺癌药物治疗
GSDMD是炎性Caspase诱导细胞焦亡的直接作用分子,GSDMD家族蛋白的N端结构域具有在膜上打孔进而诱发细胞焦亡的作用[27].有研究表明,化疗药物顺铂与紫杉醇相比,顺铂在非小细胞肺癌A549细胞中诱导更高水平细胞焦亡,表明顺铂可能在治疗具有高水平GSDME表达的非小细胞肺癌中具有额外的优势[33].此外,研究显示非小细胞肺癌患者的肺癌组织中GSDMD蛋白表达水平高于癌旁组织,给予GSDMD抑制剂可阻碍EGFR/Akt信号通路,诱导非小细胞肺癌细胞焦亡抑制肿瘤增殖[46].因此,在非小细胞肺癌的治疗中,抑制 GSDMD相关的信号传导,可诱导非小细胞肺癌细胞发生焦亡,最终延缓病情发展.
3.3.4 IL-1β和IL-18与非小细胞肺癌药物治疗
在细胞焦亡中,Caspase-1可切割和加工IL-1β前体和IL-18前体,并将成熟体IL-1β和IL-18分泌至细胞外,介导细胞焦亡发生. Wang等[47]发现,辛伐他汀通过浓度依赖性方式激活NLRP3、Caspase-1,以及成熟形式的IL-1β和IL-18来诱导细胞焦亡抑制非小细胞肺癌肿瘤的生长;还发现辛伐他汀的抗癌特性有可能会提高非小细胞肺癌的治疗效果,减少其他药物的剂量,起到减毒增效的作用.这些表明IL-1β和IL-18参与的细胞焦亡与非小细胞肺癌的治疗存在一定联系,因此,可通过调控炎症介质为突破口,如过表达或抑制Caspase的活性、阻滞IL-1β受体来诱导非小细胞肺癌细胞焦亡,有助于开发新颖的非小细胞肺癌诊疗标志物.
3.3.5 NF-κB与非小细胞肺癌药物治疗
NF-κB在细胞的炎症反应、免疫应答等过程中起到关键性作用,可使IL-1β前体和IL-18前体分泌和活化,也可促进NLRP3的表达进而合成炎症小体诱导细胞焦亡的发生[71].Teng等[59]揭示了皂苷polyphyllin VI(PPVI)可通过Caspase-1与ROS/NF-κB/NLRP3/GSDMD信号轴激活Caspase-1来诱导肺癌A549和H1299细胞焦亡抑制非小细胞肺癌的进展.Li等[72]研究表明,新型胡椒长胺 (PL) 类似物 L50377可在2.5μmol/L的浓度下诱导了大量的ROS,通过处理ROS抑制剂(NAC)可以降低L50377的活性,表明L50377 通过ROS介导的NF-κB抑制诱导非小细胞肺癌细胞焦亡.这提示NF-κB参与了非小细胞肺癌细胞焦亡,这可作为一个新的研究方向去深入探讨NF-κB在非小细胞肺癌的治疗中的作用.
上述结果提示,从细胞焦亡途径密切相关的Caspase-1、GSDMD、NLRP3、IL-1β、IL-18和NF-κB等相关信号通路入手,靶向针对细胞焦亡可挖掘出非小细胞肺癌治疗的新靶点.
4 结语与展望
综上所述,细胞焦亡是一种由Caspase激活、炎性因子释放及GSDMD裂解而诱发炎症发生,以膜穿孔、细胞肿胀、膜破裂及细胞内容物释放为特征的程序性细胞死亡方式.细胞焦亡作为一种促炎性程序性细胞死亡方式与非小细胞肺癌的发生发展及诊治密切相关.本文综述了与细胞焦亡密切相关的Caspase家族蛋白、GSDM家族蛋白、炎症小体及线粒体等相关信号通路在非小细胞肺癌中发生发展中的作用和机制,并分析了细胞焦亡激活途径中所涉及的Caspase-1、NLRP3、GSDMD、IL-1β、IL-18和NF-κB等关键靶点的药物如二十二碳六烯酸、纳米氧化锌、槐耳提取物、虎杖苷、葫芦素B、GSDMD 抑制剂、辛伐他汀等在非小细胞肺癌治疗中的作用.发现细胞焦亡途径中的任何一个环节的调控均可在非小细胞肺癌的发生发展过程中发挥重要作用,可为非小细胞肺癌由慢性炎症诱发细胞癌变提供理论基础,将与细胞焦亡关联的靶点作为非小细胞肺癌治疗新策略提供有效依据.
虽然已有研究显示细胞焦亡与非小细胞肺癌的发生发展及诊疗有着一定的关系,但其研究还处于初级阶段,仍有诸多亟待解决的问题.首先,细胞焦亡在非小细胞肺癌发生发展过程中相关的上下游信号转导通路蛋白,及其详细的作用机制尚未明确,有待进一步深入探究.其次,细胞焦亡不仅可引起慢性肿瘤细胞坏死,加速肿瘤的发展最终恶化病情;也可诱导急性炎症增强免疫反应抑制肿瘤发展进而延缓病情.因此,如何平衡细胞焦亡的发生值得我们更深入细致地研究.最后,目前细胞焦亡引发非小细胞肺癌的靶向治疗药物虽在细胞和动物模型中被不断证实,但应用于临床的较少.如果在非小细胞肺癌的治疗过程中促进肿瘤细胞发生焦亡,同时抑制正常细胞及免疫细胞死亡,便可起到清除癌细胞和保护正常组织的作用,进而控制并延缓病情的发生发展,最终提高疾病的治愈率.因此,进一步深入研究细胞焦亡的信号通路、调控机制和病理意义,这对非小细胞肺癌的早期诊断及治疗预后提供扎实的理论依据.
猜你喜欢
世界科学技术-中医药现代化(2022年3期)2022-08-22 00:24:52
医学综述(2022年7期)2022-04-19 12:31:12
昆明医科大学学报(2020年12期)2021-01-26 00:44:42
中华养生保健(2020年9期)2021-01-18 03:12:32
世界科学技术-中医药现代化(2020年2期)2020-07-25 02:06:26
中国组织化学与细胞化学杂志(2017年1期)2017-06-15 20:27:43
中外医疗(2016年15期)2016-12-01 04:25:44
中华老年多器官疾病杂志(2016年9期)2016-04-28 08:52:49
西南军医(2015年3期)2015-04-23 07:28:32
国际心血管病杂志(2015年5期)2015-02-27 12:11:34
