
甘肃兰州海石湾中侏罗世Coniopteris nikaensis 的大化石及原位孢子特征*
2022-11-02 04:04:54辛存林焦志鹏刘松欣狄国芸
地质科学 2022年4期
赵 涵 辛存林,2 焦志鹏 刘松欣狄国芸 张 博
(1.西北师范大学,地理与环境科学学院 兰州 730070;2.中国科学院南京地质古生物研究所,现代古生物学和地层学国家重点实验室 南京 210005)
蚌壳蕨科(Dicksoniaceae)的锥叶蕨属(Coniopteris)是 Brongniart 于 1849 年根据约克郡侏罗纪植物区系中的化石而建立(Doweld,2013)。该属地理分布广泛,主要分布于北半球,是构成中生代植物群的主要分子(Taylor et al.,2009;吴向午等,2019),特别是在中国、中亚和西伯利亚等地(Deng and Lu,2006)。全球范围内,锥叶蕨属植物化石已描述过的约130 余种(Taylor et al.,2009;吴向午等,2019),至今仍然有新种被报道(Cantrill and Nagalingum,2005;Kostina and Herman,2013;Scanu et al.,2015;袁修才,2018)。根据PBDB 数据库(The Paleobiology Database)资料,锥叶蕨的起源可以追溯到二叠纪时期,但只在《Geology of Korea》一书中有一处化石记录(Lee,1988),三叠纪时期在全球范围内零星出现,侏罗纪时期最为繁盛,其中中侏罗世时期达到顶峰,早白垩世晚期逐渐衰退。锥叶蕨属植物多生长于温带、亚热带地区,在热带地区也有个别记录,是温暖、湿润环境的指示分子(Vakhramee et al.,1991;Wang,2002)。
锥叶蕨属的研究已达一个半世纪,在国外,Harris et al.(1974)在20 世纪60 年代就系统研究了锥叶蕨属(Coniopteris)的原位生殖器官;Вахрамеев(孙革等,1990)曾研究了发现于布列亚盆地内的锥叶蕨属植物原位孢子的形态特征;Van Konijnenburg-van Cittert(1989)深入研究了英国约克郡植物群中锥叶蕨属植物化石原位孢子的形态特征;Balame(1995)系统总结了10 余种锥叶蕨属植物化石原位孢子的形态特征;Bystritskaya and Rychkova(2013)系统描述了发现于西西伯利亚的6 种锥叶蕨植物化石。在我国,该属植物化石主要发现于北方地区,包括辽宁北票、青海柴达木盆地、北京门头沟和甘肃窑街盆地、潮水盆地、宝积山盆地等,已经发现40 余种(叶美娜等,1986;邓胜徽等,2001;孙克勤等,2010)。国内学者对蕨类植物微细构造的研究不断深入,马清温等(1996)总结了锥叶蕨属植物的研究进展;邓胜徽等(1995,2001)详细探究了蕨类植物化石生殖器官的研究方法;张泓等(1998)总结了中国西北地区的真蕨类植物化石记录;邓胜徽等(2001)细致研究了中国东北地区早白垩世真蕨类植物化石的生殖器官;Zhou et al.(2021)系统研究了四川盆地三叠纪—侏罗纪过渡期真蕨植物的多样性变化和生态环境特征;Sun et al.(2021)描述了产自新疆准噶尔盆地白杨河地区的膜蕨型锥叶蕨原位孢子的形态特征。虽然对锥叶蕨的微细构造研究至今成果颇丰,但仍缺乏对部分锥叶蕨的原位孢子特征的研究。
长期以来,我国对尼勒克锥叶蕨化石的生殖器官研究得不多。张泓等(1998)首次在新疆伊犁盆地尼勒克吉仁台中侏罗统胡吉尔台组发现了Coniopteris nikaensis化石,对其进行了系统描述和命名,王晶菁(2020)于宝积山盆地发现了C. nikaensis,但都未对C. nikaensis的原位孢子进行研究。本文的C. nikaensis是在窑街盆地窑街组的首次报道。对窑街盆地古植物的研究至今已有近40 年的研究历史(孙柏年等,1986;阎德飞等,2004;张成君等,2004;辛存林等,2006,2010,2011,2012,2017a,2017b;Xin et al.,2018),该盆地中侏罗系广泛发育,并保存有精美的植物化石,是研究侏罗纪化石植物的理想地区之一。众多学者对窑街盆地的植物化石进行了较为细致的研究,如孙柏年(1986)总结报道了甘肃窑街盆地植物大化石的发现情况;阎德飞等(2004)研究分析了采集于窑街盆地内的穆雷似管状叶(Solenites murrayana);张成君等(2004)研究分析对比了窑街组银杏化石和现生银杏角质层的有机质组分,重建了古环境;辛存林等(2006)分析了窑街盆地的中侏罗世孢粉化石,建立了孢粉组合。迄今为止,甘肃窑街盆地已报道了7 种锥叶蕨化石(孙柏年等,1986;辛存林等,2006,2010,2011,2012,2017a,2017b;Xin et al.,2018),凸显了蕨类植物在该盆地的多样性特征。针对窑街盆地植物群的组成研究已久,但仍有新的蕨类植物在窑街盆地被发现,需要进一步进行研究,完善窑街植物群的组成,恢复窑街植物群面貌。
本文对采集于甘肃窑街盆地海石湾地区中侏罗统窑街组的C. nikaensis的裸羽叶和实羽叶进行了细致研究,完善了C. nikaensis的羽叶、孢子囊及原位孢子方面的数据。本次研究不仅为完善锥叶蕨属的系统演化提供了材料,同时为准确探索该属的自然地理分布、生境特征提供了有力证据。
1 研究区概况
红古区海石湾镇(北纬36°34',东经102°86')位于兰州市西北部,海拔约1 800 m,地处甘青两省的交界位置,距兰州市区约110 km,南部有京藏高速与京拉线经过,交通便利。境内分布有湟水、大通河、庄浪河等(图1)。海石湾地处窑街盆地东南部,窑街盆地在大地构造单元上属于中祁连隆起带,处于祁连褶皱山系与兰州古地块的交接部位,是中生代基底断裂活动所形成的一个内陆盆地。盆地内侏罗系地层发育,其中窑街组是一套中侏罗统的含煤地层,其沉积环境主要为河流—湖沼相,地层总厚度约235 m。窑街组自下而上可分为5 个岩性段:砂砾岩段(J2y1)、含煤岩段(J2y2)、铝质泥岩段(J2y3)、油页岩段(J2y4)、砂泥岩段(J2y5),大致相当于欧洲地区的阿林阶—巴柔阶。本研究的化石标本采于油页岩段(J2y4),该岩段含有丰富的植物化石(图2)。
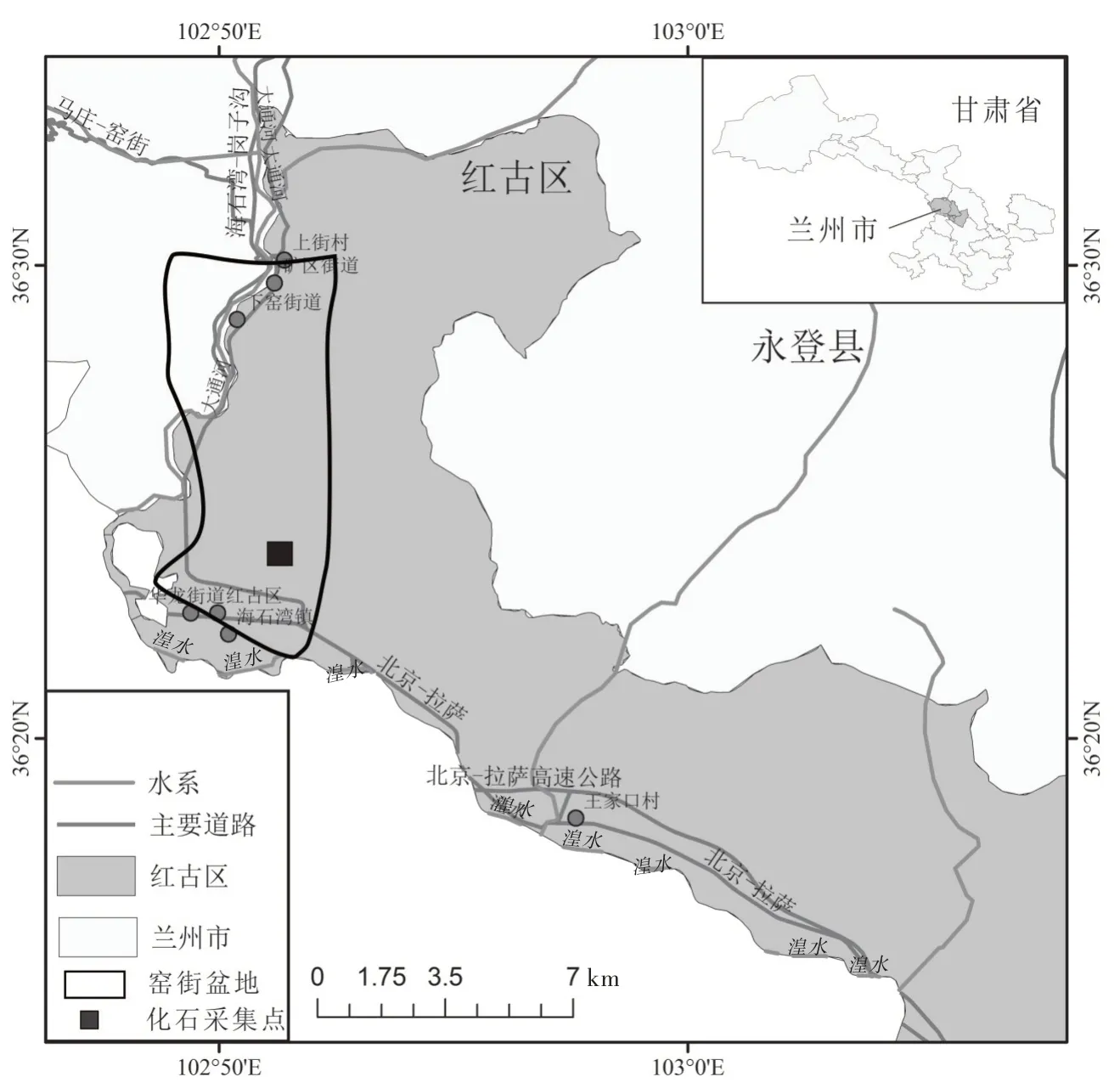
图1 研究区概况图Fig.1 Overview map of the study area
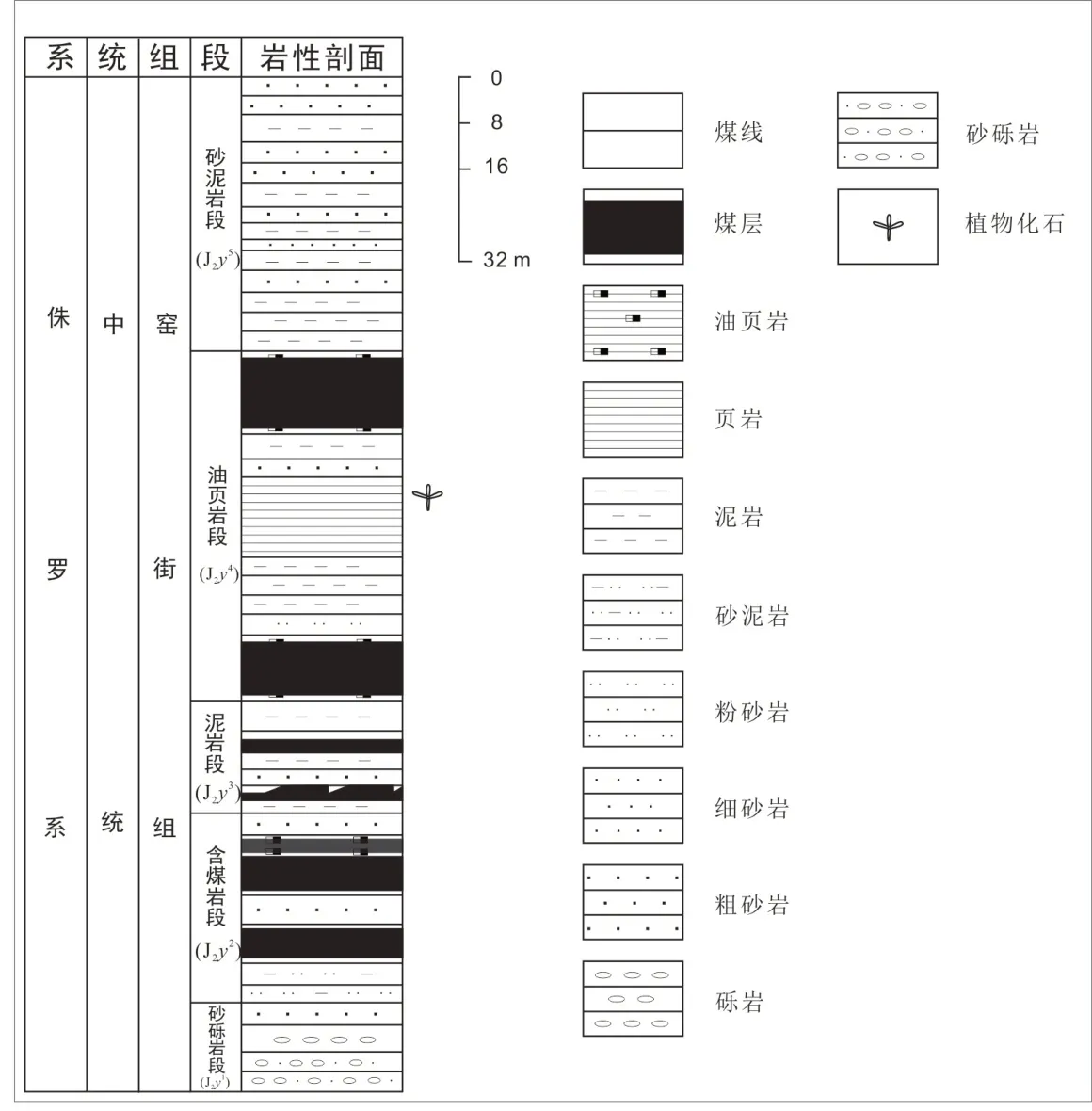
图2 地层柱状图(据孙柏年,1986 修改)Fig.2 Stratigraphic bar chart(modified after Sun,1986)
2 材料与方法
本文研究的锥叶蕨属植物化石材料,采自甘肃窑街盆地海石湾镇矸石堆。根据调查该矸石堆为海石湾煤矿所堆积,开采自窑街组油页岩段(J2y4),当前标本同时保存有营养羽片和生殖羽片的锥叶蕨化石标本,保存为压型化石,使用佳能650D 数码相机和NSZ-606 体视镜对标本及孢子囊群微细结构进行拍照留存。此外,在Nexcope NE900 显微镜下对孢子团块及孢子进行观察拍照。锥叶蕨植物化石标本、孢子薄片均保存于西北师范大学地理与环境科学学院地质标本室。利用Adobe Photoshop 及CorelDRAW 等软件进行了文中所有图片的绘制及图版的制作。
原位孢子的处理(Homes et al.,2015):首先,在化石标本上选择含有清晰的孢子囊群位置,将选中的几个孢子囊群用解剖刀剥离,再将样品装入10 ml 的塑料离心试管中,然后通过在工业级乙醇中漂洗,将孢子囊与孢子团分离;静置待乙醇蒸发后,再用30%的HCl 去除样品中钙质杂质;然后把样品洗至中性,再加入40%的HF 冷处理24 h,去除样品中的硅质和粘土;洗至中性,再置于6%的H2O2中澄清至少3 小时。吸取部分样品观察,若发现碳质碎片变黄,孢子或孢子团块清晰可见,则停止处理。用苯酚甘油凝胶制片,观察并照相,其余部分孢子样品保存备用。
3 化石标本描述
真 蕨 类 植 物 Filices(Brongniart)Braun, 1864; 蚌 壳 蕨 科 Dicksoniaceae Engler,1902;锥叶蕨属ConiopterisBrongnart,1849;尼勒克锥叶蕨Coniopteris nikaensisZhang et Sun, 1998;(1998Coniopteris nikaensis张 泓 等 , 页 272, 图 版 13, 图 1.; 2020Coniopteris nikaensis王晶菁,页14,图版Ⅰ,图f.)。
(1)特征描述
蕨叶及羽轴:末二次全羽片。羽轴宽1 mm,有竖纹。羽叶宽约10 cm。羽片长约6 cm,基部宽约1 cm,尖部宽约6 mm,两羽片羽轴之间距离约1 cm,相邻羽片相互接触或略叠覆(图3a)。
裸羽片:羽片互生,与羽轴呈45°~50°夹角,羽片呈披针形,自基部向顶端逐渐变窄。小羽片呈卵圆形至菱形,密集排列,叶脉清晰,楔羊齿型,稀疏,中脉与侧脉等粗,终止于裂片边缘顶端在,侧脉分出角度20~30°,不分叉。小羽片最宽处多在基部,边缘深或浅裂,顶端钝圆,裂片5~7 片,裂片钝圆(图3b、图3c)。
实羽片及孢子囊群:实小羽片叶膜略微显退缩。实羽片同样为二次羽状分裂,末次实羽片长约5 cm,基部宽约7 mm,尖部宽约4 mm,轴宽约1 mm,于约50°夹角着生于叶轴之上,两实羽片羽轴间距约8 mm。孢子囊生长成群,着生于叶的裂片顶端,直径约1 mm,呈卵圆形,囊群盖为杯状。每一小羽片上有2~3 对孢子囊群,部分顶部裂片也发育囊群而至使囊群数目变成奇数(图3d~图3h)。

图3 尼勒克锥叶蕨Coniopteris nikaensis Zhang et Sun 蕨叶形态及其素描图Fig.3 Leaf morphology of sketch Coniopteris nikaensis Zhang et Sun
孢子:在光学显微镜下观察,C. nikaensis的原位孢子基本特征可概括为:孢子三缝,赤道轮廓亚三角形,三边微凹,顶角钝圆,直径平均约35 μm。射线较窄且平直,具微弱发育的唇,长度约为孢子体半径的3/4 或更长。外壁薄,表面光滑(图4)。
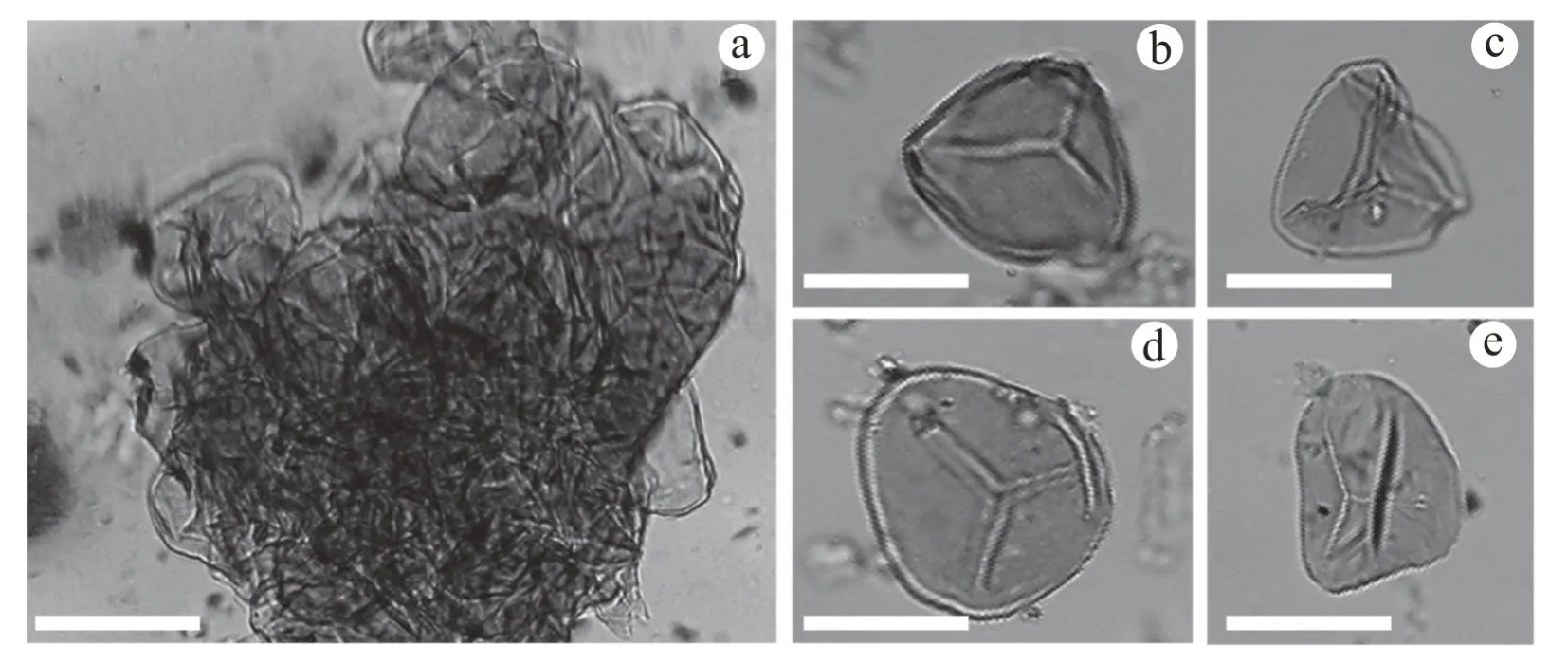
图4 尼勒克锥叶蕨Coniopteris nikaensis Zhang et Sun 原位孢子Fig.4 In situ spores of Coniopteris nikaensis Zhang et Sun
(2)产地层位
海石湾窑街组油页岩段(J2y4)(图2)。
4 比较与讨论
当前化石实羽片退缩,孢子囊群着生于叶的边缘,这一特征仅见于蚌壳蕨科(Dicksoniaceae),因此当前化石应归入蚌壳蕨科。该植物化石的羽片互生,以45°~50°夹角着生于轴上,羽片呈披针形,自基部向顶端逐渐变窄;小羽片呈卵圆形至菱形,密集排列,边缘深或浅裂,分裂2~3 次;叶脉细,楔羊齿型,楔羊齿型,稀疏,中脉与侧脉等粗,终止于裂片边缘顶端在,侧脉分出角度20°~30°,不分叉;实小羽片叶膜略微退缩;卵圆形孢子囊群着生于实小羽片顶端。将这些特征与蚌壳蕨科各属的特征进行对比(表1),发现当前植物化石形态特征与锥叶蕨属(Coniopteris)最为相似。虽然蚌壳蕨属(Dicksonia)的一些特征与当前化石也较为相似,叶脉都为楔羊齿型,裸小羽片披针形,囊群顶生,但蚌壳蕨属的中轴较宽,宽2 mm,叶脉较为稀疏,且实羽片与裸羽片同形,而当前化石中轴宽1 mm 左右,实羽片较裸羽片叶膜略显退缩。相较爱博拉契蕨属(Eboracia),叶脉和裸小羽片都为栉羊齿型或枝脉蕨型,而当前植物化石的叶脉为楔羊齿型,同时中轴宽度相比爱博拉契蕨属较细。屈囊蕨属(Gonatosorus)的蕨叶是2 次羽状分裂,中轴较细,但叶脉和裸小羽片都为枝脉蕨型,且实羽片是混生型。同样,刺蕨属(Acanthopteris)是两次羽状分裂,但中轴粗,可达5~8 mm,裸小羽片为栉羊齿型,且实羽片为独立或混合型,着生于蕨叶的上部或顶部。综上所述,从形态学特征上可将当前标本明确为锥叶蕨属植物化石。

表1 蚌壳蕨科相关属蕨叶形态特征对比Table 1 Comparison of morphological characteristics of fern leaves in related genera of Dicksoniaceae
小羽片的分裂程度是鉴定锥叶蕨属中不同植物化石种的重要依据之一。当前化石的小羽片分裂程度较高(正常小羽片分叉2~3 次)。根据已有报道的中国境内化石材料的描述和图版,发现共有12 种锥叶蕨属植物化石的小羽片分裂程度相对较高;将当前植物化石与这12 种锥叶蕨属植物在蕨叶、羽片、叶脉、裸羽片、实羽片、生殖型等特征方面进行了详细对比(表2)发现,当前标本与新疆伊犁盆地尼勒克吉仁台胡吉尔台组的Coniopteris nikaensis最为相似(张泓等,1998),C. nikaensis是张泓等于1998 在《中国西北侏罗纪含煤地层与聚煤规律》一书中建立的新种。当前标本与C. nikaensis的羽叶形态均为二次羽状分裂,羽轴竖直,具纵沟,叶脉细,楔羊齿型;二者小羽片都为菱形,小羽片浅裂和半深裂,两侧对称,分叉2~3 次,有5~7 个小裂片,末次羽片基部的小羽片最大,其中C. nikaensis相邻羽片相互接触或略叠覆这一特征与当前标本最为相似,但C. nikaensis尼勒克锥叶蕨羽轴相较当前标本的羽轴稍宽,同时C. nikaensis的
实小羽片未退化,当前标本的实小羽片略微退化。当前标本与C. nikaensis的孢子囊群直径都为1 mm,都单生于实小羽片上侧半深裂的长圆裂片顶端。因此,将当前化石归为C.nikaensis是适宜的。

ilar sp eciesits sim比对的征ika en sis an d特态形类iopteris.n种etween C on似相其ikaensis 与iopteris.non ological characteristics b 2 Corph表rison of m paTable 2 C om献文等等等等等考,1984芬娟, 1 9 8 8, 2 0 0 1徽徽, 2 0 0 1芬,1984参陈佩胜胜邓李邓陈型型型型型型实殖合实合实混生全混全m位圆群椭m 1.5 m 群椭位圆m 柄片圆囊群片囊横 子囊 囊群 群椭 裂至2 m子囊1.5 m囊群 的子群羽于形孢,片狭孢囊实孢,小1×子囊,端位圆,端1×羽缩顶瘦为孢,1.5×群椭。全小 ,, 大 缩顶为 , 成囊群形缩端 小实小烈裂,为(1~1.5)×0.8 mm缩端为 缩退片退顶 大形子囊退顶小大强于退不片, , 不片大 膜退片孢,膜位圆膜裂 膜于叶于形 椭 叶位形 ,端膜裂,圆叶叶群状顶°,,片顶略裂,小,第,,裂羽微不,尖部 小微,叶40羽片,片缩为3~7 个顶端钝圆裂片深片尖自°~小羽收离,裂离钝基分钝,部裂延羽生分,变下小50微裂°~或,小态或钝不°着,或 角从右分或,锐生基浅分°着微尖具剧对片3~态5 个或触端,40片°左m,部形°着度35急一态60蕨变生微锐延裂尖裂不尖35触°~接顶延间尖下 间片下 生部态基40端。并第变之端缩 片羽呈着顶以之羽缩 ,片顶收152m裂以钝深着以接,片小收缩部小裂 顶片,片态,端,羽片部为1~尖 片收基生羽,部对半羽变生部变距端叶于形小羽基生 羽生顶延部片一小羽互片下具,到互小离基 渐间 互小小基羽m,=0°,长形宽互下1 m,裸,m,长长30 6 m宽=m,∶°~6状宽形针m,宽宽=0∶8 m 披4 mm,形楔0.5 m长= 2∶1生∶8 m,m,8∶小 ,形m,以着或长宽∶5长长宽大,m,2∶1宽形3~0.3 m= 1纵宽3∶2,或菱2 m 卵长宽∶形4 m 形m,∶圆 长卵形5 m 长,,长卵,细近消菱侧脉型弱等很,片锐侧大,近度叉羽 齿显纤,处 与侧分中细不片 以,细长缘其脉分,显半侧出歧到脉角分小 羊明 脉,或裂脉 ,出粗叉2~3 次,直达裂片端型明一 中粗前与伸二展叉1~3 次楔不 细伸脉, 尖脉脉,脉齿 不 的 , 等1 次 一1 条每含脉脉,延很轴中羊脉膜状近叉,等,侧弱中 脉自和叶楔中叶叶中侧小并羽脉分叉 中角脉相生极互散端致对.8;m 约 以距顶细0°着10 m ,间互m,间生形1m末长,;,1 cm ,间羽片,°~66~;m 针细片40角约形生以片近<0紧形纤羽较约或约三宽m 互m,羽披生3 m生宽次轴; , 末大线互轴羽°40,羽 或轴或列宽距1 m到形羽°着片形1 m间2~ ;较形形次 针次70形以片次°着约 线约针角排生距长生mm,羽 末40角末至宽尖披末°~ 三片;50披,形轴距次2 cm,针羽末片片,m;羽生2 m羽渐三;羽生以羽生35披次~5,m;;°~,长,,m;裂7 mm m分,m;m裂裂上m;m裂5 m 2 m裂7 mm<分宽 分宽 分80 m羽m,约2 m 4 m分叶状状宽2~mm,宽状m 以0 m状0~宽 宽羽, 羽m,蕨轴<宽轴宽轴次0°着次羽状二24 m二不轴 次10次轴次明 羽0 m三长羽10宽羽 二40 m羽长150.5 m羽sis羽长长ylloid heen C.bella C.beijin 1.5 m种二25 es gensis na名C.concin olinC.hu分宽C.hymenoph
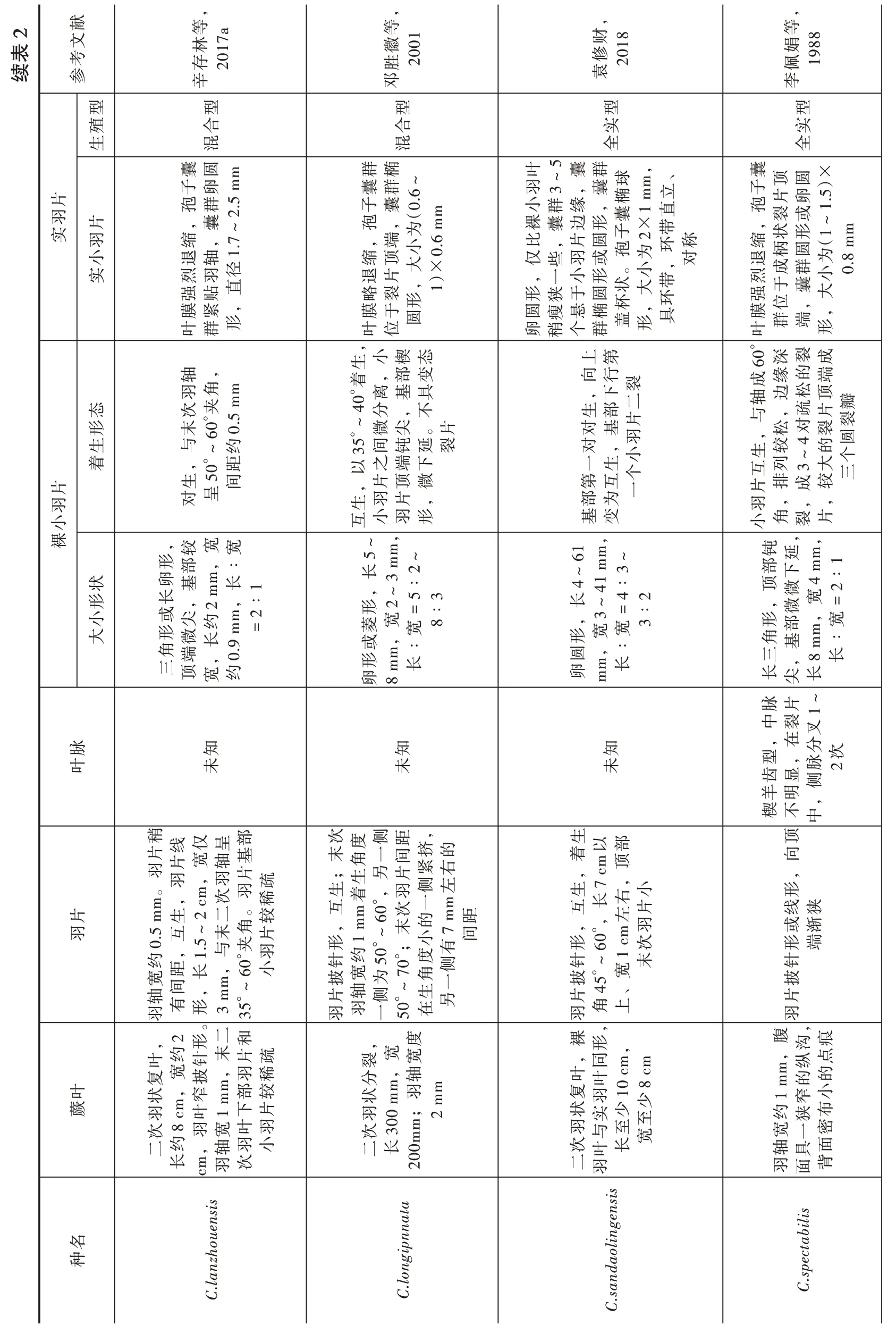
2献等等财等文林徽修娟考, 2 0 0 1,2018, 1 9 8 8参存, 2 0 1 7 a胜佩辛邓袁李型型型型型实殖合实混全生合全混5 m囊圆 叶羽3~囊群球m,、孢群.5 m囊子卵囊群顶圆,囊椭片立子囊群椭片卵小群缘,囊1 m羽直,囊 (0.6~1)×0.6 mm 孢,裂或片实裸囊边形 ,缩, 比,片圆为孢为环称羽,端小 子2×带 缩状形1~1.7~2退轴小大仅些羽或缩顶退。小,对 柄圆(为实成群退片, ,一小形 烈1.5)×0.8 mm烈羽径 状大带 小强强贴直于囊略裂 杯,环形狭于圆膜紧, 形位,大圆叶群形 圆瘦悬椭盖形具膜于膜群端,叶位 形上第60子,卵稍个群生小楔态向行羽角m轴,°成缘的端裂深裂成,下40°着,部变态末0°夹次二离基具0.5 m轴边松顶瓣与,疏片°~微尖。片形裂片对基分,不生,°~6间钝延裂生部对,约35羽 ,松的圆一生 生较与生50着小大叶以之端下距个,片顶微 互列3~三第互片生羽片,间对呈个片排成较基变羽部为 羽,,,互小羽形一孢小角裂片小,较宽裸卵基m,∶61m,钝延m,状长顶微宽= 2∶1部下形部,宽= 2∶4 对裂长5~8 mm,宽2~3 mm,形2~8∶3或尖长,= 5∶2 m,微4~菱 宽形形微长约4 m1,3~= 4∶形部长角基,0.9 m 41 m小m,形宽宽或∶大圆角端 长三,8 m 3~3∶2∶三顶卵mm,∶形长长长尖长脉片脉卵知知中裂,在分知宽约叶未,未型,未齿显脉侧,楔不羊明中片线仅呈部顶次度侧距,叉1~2 次向稍;生另片紧右片宽轴基 末角一间挤的,羽着,生,m。羽生次羽稀 生互m 着°, 羽 侧,形互长右小形1 m线狭二。较顶生,°, 羽0.5 m互次 一m 左距羽,1.5~2 cm,羽片疏片末角片形60或渐60与1 cm 左片针约50°;小有, °~末的7 m间形端披宽为70次距长7 cm 以部约m,0°夹 羽小末侧有形3 m°~6针针°~宽度宽间,披一披45角另片轴侧°~生轴35片 、羽角上片羽羽羽一50。羽,在,2 m,纵点,同形二和 ,次8 cm,窄,叶约针末片疏叶裂宽宽复宽披叶叶状羽的的分m,轴状裸形复羽m,羽稀部较8 cm叶1 m羽0 m 1 m状实少蕨少腹沟痕次30m;下片约狭布至羽二约羽宽叶羽窄小二长0m 10 cm,度 2 m m羽与至宽 宽一密次叶长长cm,轴羽小20二羽羽次轴具面羽面背a taipnn sisaoling名C.la nzhouensis C.long nd m,宽en种C.sa bilisC.specta表续

2献等等等文芬徽泓文考,1984, 2 0 0 1,1998本参陈胜邓张型型型型实实殖实全全实全全囊顶生囊扁子生裂宝退其囊圆片小子、为子片 孢单深端 显, 状孢形 子卵微小孢裂羽杯m元片,或,圆 ,或 化,m,半侧顶大孢呈略很实为羽圆缩群形 形缩片 退1 m 上片 形,膜片裂梯椭1.5 m盖退囊圆 退羽小叶羽不约片 圆大数1×群圆烈小群实 烈,片小生群囊囊强短 长强于羽实着囊,羽直小少膜较小群实膜位 ,,的小。端,形形顶群叶柄 部 实缩,,,叶群 实囊于生羽宽对下侧部,,圆形侧0°着 小 较裂钝生分、。菱两°~4或急较片,部片长,侧即上上0°着 微 尖 宽片径羽浅片明基裂态,裂°~6或裂触端部裂间片,变裂对浅深30不,态深,深不或半小浅称,称裂裂接顶基态距锐变形缘生以间尖具50半侧缘或以之羽圆具 变或对片 缘 边3 次和两全裂片端不,片 ,片小或不片2~羽全两:,;生羽着羽裂;边深生羽,尖互小离钝顶互小浅称侧边小边羽裂的侧小片小分裸m,部伸宽菱末片宽片下渐密约m,1 2 m 长。羽叶逐m,=1∶1宽1 cm,,长小mm,羽m,=间蕨上状,0.5 m= 4∶宽形先的,,2~3 m 的10 m ∶和出小长1形片3 cm,距长菱羽约轴= 2∶宽菱式部10∶形宽∶形小宽行基长中长m,2∶ 至,宽羽1约宽之在向大长形长化圆,m,部约大形长长卵、下片, 长菱形次最7 m 圆生形,羽大m,,变卵长2 m羽形5 m卵集6 cm,片离∶近两菱宽型齿,型楔羊知知未未叶细齿羊楔脉为脉脉叶粗细50近羽触型°末50次°~距羽于相覆于上形间,宽蕨生 等具 ,,。叠小叶 互;以片 中上角生纹上略相生m,羽m 轴形0°着叶呈片互竖轴或羽角 羽m,互°~接45片形0.1 m 次末4 m叠三覆,有于触,2~ 轴羽羽形或,形次三50,1 m主邻末在 略或针m,生接°~8°着互生生m,针线宽 披针约 ;相披形约与相呈披宽°着 ,;50形宽°~轴对1 m片以片片,长轴70羽羽细纵相纹交羽叶轴45邻,m ,主纹裂m,;主,裂裂竖,直m;1 m分m, 分0 m 10 m裂有上m度分1 m羽20 m蕨叶状<分m,沟状10m 以1 m纵轴状宽 次大50 m状m,羽2 m羽于 宽次长宽具羽2 m二长宽轴三羽次宽羽二轴次宽二轴en sta sis en名sis种tu ng C.venu C.ta,竖C.nika表续
当前海石湾标本的原位孢子在形态上可与表3 中10 种锥叶蕨原位孢子相比较,孢子形态均为四面体形,除C. bella的孢子极面观近于圆形外,其余9 种锥叶蕨的孢子极面观都为亚三角形或圆三角形。与当前标本孢子赤道轮廓最为相似的为C. hymenophylloides,C. lanzhouensis和C. simplexz三种锥叶蕨的原位孢子。与当前孢子直径最为接近的为C. hymenophylloides,C. lanzhouensis和C. longipnnata三种锥叶蕨的原位孢子。其中C. hymenophylloides和C. longipnnata与本文的原位孢子十分相似,前两者三射线直而窄,体积均为30~40 μm,不具唇状边缘;不过,当前C. nikaensis孢子三射线平直且较窄,其长度约为半径的3/4,相较前两者较短,从而与前两者区别开。

表3 锥叶蕨属孢子形态对比Table 3 Contrast of spore morphology of Coniopteris
该种在建立之初,仅描述了孢子囊群的特征(张泓等,1998),并未描述其原位孢子特征。本次研究补充了C. nikaensis的原位孢子特征,为尼勒克锥叶蕨的深入研究提供了新材料。
5 古地理分布及古环境意义
植物大化石是良好的气候及环境指示剂。不同植物有不同的习性,特定的植物一定是生长在特定的环境中,比如羽片有无滴水叶尖等特征都能反映各自所处的环境(袁修财,2018)。现生蚌壳蕨科是树蕨,植株不高,对湿度的敏感程度高于温度,对水的利用效率很低,并且不能长时间处于干旱环境中,一般生活在潮湿、阴暗的密林中心区(Passioura, 1982; Franks and Farquhar, 1999; Dignan and Bren, 2003; Sack and Holbrook,2006;Volkova et al.,2011)。根据“将今论古”的原则,通过分析现生蚌壳蕨科植物的生境,可推测出中生代的蚌壳蕨科植物生长区域的气候同样为温暖湿润。
根据植物化石的植物体保存状况、植物体分选情况、方位类别及围岩特性,可将植物化石埋藏类型分为原地埋藏、亚原地埋藏以及异地埋藏3 个类型(表4)。
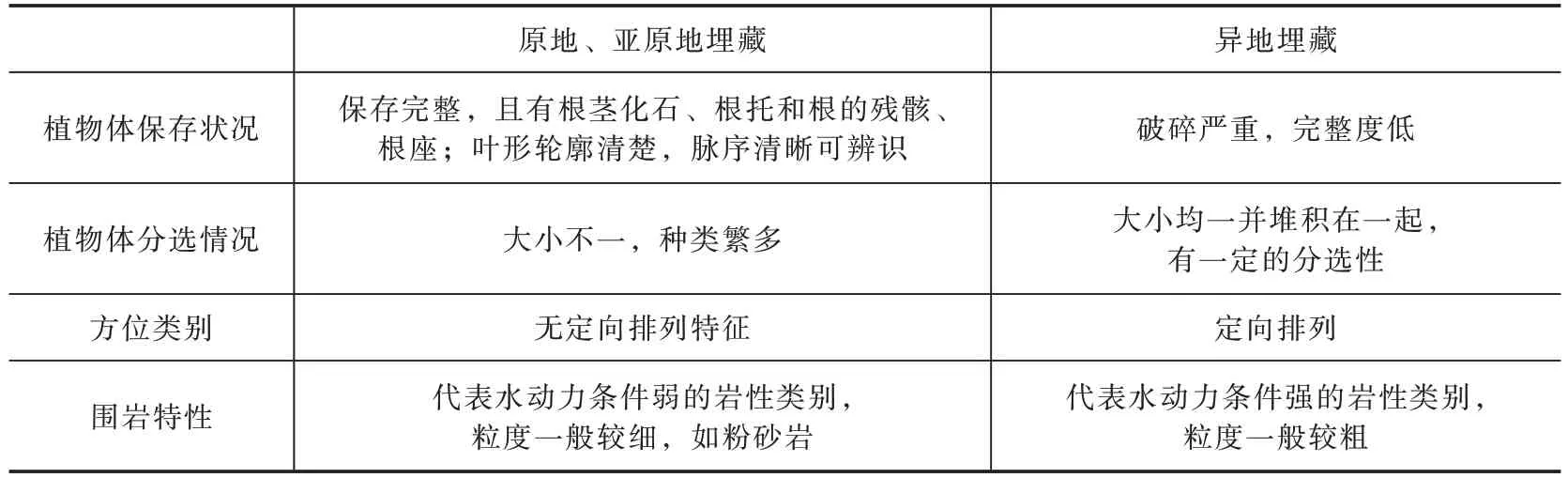
表4 植物化石埋藏类型判断标准(据张英芳等,2006 修改)Table 4 Judgement standard of plant fossils buried type(modified after Zhang et al.,2006)
将与当前标本一起采集到的其他植物化石标本进行埋藏学分析,发现都保存完整,且部分保存有根的残骸等,植物叶形轮廓清楚,叶脉清晰,植物种类多样,大小不一,并且化石赋存的岩性为炭质泥岩或砂泥岩,岩石表面具有较细的平行层理构造。通过分析表明,本文所采集的化石主要为亚原地埋藏和原地埋藏,因此该植物群组合的特征可以作为反映古生态环境的依据。
C.nikaensis在窑街盆地的发现属于该化石的第三次报道,它首次发现于新疆伊犁盆地,在伊犁盆地的整个侏罗系地层中均有发现(张泓等,1998),后于宝积山盆地中侏罗统中发现(王晶菁,2020)。根据辛存林等(2019)对窑街植物群植物地理归属的划分,发现宝积山盆地、窑街盆地与伊犁盆地都位于北方植物群中西北部一个地块单元中,3 个盆地的古环境具有一定的相似性,但在锥叶蕨属植物的物种繁荣度上存在一定的差异。
将窑街植物群中锥叶蕨属植物种类与甘肃宝积山植物群和新疆伊犁盆地胡吉尔台组植物群(以下简称胡吉尔台植物群)中的锥叶蕨属植物对比分析(表5)。锥叶蕨属植物在宝积山植物群中有10 种,占植物群总种数量的15.1%,而在窑街植物群与胡吉尔台植物群中的占比相近,分别为12.5%和12%。虽然两植物群中锥叶蕨属植物占总数的比例仅相差0.5%,但在种数上相差较大,窑街植物群有8 种Coniopteris而胡吉尔台植物群中仅有3 种Coniopteris,仅从锥叶蕨的物种数量上看窑街植物群与宝积山植物群更相近。

表5 三大植物群中锥叶蕨属的对比Table 5 Comparison of the Coniopteris in three major flora
王晶菁(2020)将宝积山植物群中的真蕨类植物归入了低地蕨类—苏铁类植物群落,该群落的真蕨类植物多样性最高,这一群落往往分布于盆地内部地形较低缓,土壤相对潮湿的山坡低地,推断出宝积山地区中侏罗世的古气候温暖而潮湿。张泓等(1998)将胡吉尔台植物群归入了锥叶蕨顶峰带,该组合带中锥叶蕨属植物不仅种类多样,而且分异度高,又根据沉积构造推断侏罗纪伊犁盆地内低洼分布广泛,具有丰富的水源补给,属于常湿温暖气候区。通过对比,发现在古气候方面3 地相近,都属常湿温暖气候区,但在古环境方面,窑街盆地与宝积山盆地存在更利于Coniopteris的生长繁殖的环境,以至于与胡吉尔台植物群的Coniopteris物种数量相差较远。
6 结 论
(1)当前植物化石蕨叶为末二次羽片,羽片互生,以45°~50°着生于轴上,叶形呈披针形。小羽片呈卵圆形至菱形,叶脉清晰,楔羊齿型,稀疏,中脉与侧脉等粗,终止于裂片边缘顶端在,侧脉分出角度20°~30°,不分叉;小羽片边缘深或浅裂,分裂2~3 次,裂片钝圆。相邻羽片之间相互接触或略微叠覆。
(2)孢子囊群着生于实羽片裂片顶端,直径约1 mm,呈卵圆形,囊群盖为杯状。孢子三缝,赤道轮廓亚三角形,三边微凹,顶角钝圆,直径平均约35 μm。射线较窄且平直,具微弱发育的唇,长度约为孢子体半径的3/4 或更长。孢子外壁薄,表面光滑。
(3) 通过与已报道的锥叶蕨属植物化石形态特征的对比研究,发现当前化石与Coniopteris nikaensis特征基本一致,因而将当前化石标本归为C. nikaensis是合理的。对本次采集到的标本进行埋藏学分析,具原地—亚原地埋藏特征。将窑街植物群、宝积山植物群和胡吉尔台植物群中的Coniopteris种类对比,仅从Coniopteris种类上来看,在古环境方面,窑街盆地与宝积山盆地都存在更为适宜Coniopteris生长繁殖的环境。
猜你喜欢
幼儿100(2024年19期)2024-05-29 07:43:34
美文(2021年12期)2021-07-01 14:30:45
美文(2021年10期)2021-05-24 04:46:43
美文(2021年10期)2021-05-24 04:46:43
美文(2021年10期)2021-05-24 04:46:43
自然保护地(2020年2期)2020-06-25 11:31:50
种子(2019年7期)2019-01-06 22:12:38
天然产物研究与开发(2018年5期)2018-06-13 03:23:54
贵州工程应用技术学院学报(2016年5期)2016-12-17 06:15:40
黄山学院学报(2016年5期)2016-03-16 01:31:34
