
低温等离子体对粳稻表面霉菌的抑制作用及其品质的影响
2022-10-26 04:41王建峰刘兴泉
中国粮油学报 2022年9期
谢 辉, 王建峰, 曹 涌, 刘兴泉, 李 玲, 郭 俭
(浙江农林大学食品与健康学院; 现代粮食产业学院,杭州 311300)
稻谷是我国第一大粮食作物,其总产量与播种面积位居世界第一[1]。稻谷作为主要的粮食食用品种,供应着一半以上人口的口粮[2]。据联合国粮食及农业组织(FAO)2019年对全球农产品各环节粮食损失指数估计,全球约14%的粮食在从生产至零售环节之前被损失掉,约3.8亿t[3]。据2020年国家粮食和物资储备局数据显示,我国每年有约700亿斤的粮食在储藏、运输和加工等环节不断损失[3]。这对我国储粮工作提出了更高的要求。
为提高稻谷耐储性及品质,常用于稻谷储藏的技术有低温储藏技术、气调储藏技术和药剂熏蒸技术等。然而,低温储藏技术和气调储藏技术对仓房的气密性和隔热性要求严格,仓房的改造费用昂贵,稻谷储藏成本较高[4]。而使用药剂熏蒸会不断提高储粮害虫的耐药性,必须不断提高熏蒸药剂的浓度才能实现有效熏蒸[5]。
等离子体,又称为电浆,被认为是物质除固态、液态、气态的第四态,是原子或分子被电离后产生的离子化气体状态物质,可以分为高温等离子体和低温等离子体[6]。当电子温度远高于离子温度,体系处于非热平衡状态时,体系中的气体为轻度电离,放电温度接近室温,故被称为低温等离子体。低温等离子体是一种新型食品冷杀菌技术,通过电离空气产生活性物质作用于食品表面的微生物达到杀菌的目的[7],具有高效、快捷、安全无污染等优点。目前,该技术已在如肉类、果蔬和水产品等食品中有相关的应用研究[8-10]。徐慧倩等[11]证实低温等离子体能有效减缓南美白对虾的腐败变质,并且随着冷藏时间的变化,经过低温等离子体处理的南美白对虾的质构特性、色泽的保持效果都显著优于对照组。乔维维等[12]研究发现低温等离子体能够有效杀灭牛肉表面的微生物,降低牛肉的细菌残留量,并且较好保持牛肉的色泽。因此,低温等离子体在农产品保鲜和储藏上的应用也越来越广泛[13],但目前该技术应用于粮食储藏方向的相关报道较少。
前期课题组通过对仓储稻谷霉菌群落进行分析,选择亮白曲霉、黑曲霉、米根霉、草酸青霉4种稻谷储藏优势霉菌作为研究对象,研究低温等离子体对粳稻表面4种霉菌的抑菌效果与粳稻的品质变化之间的关系,为低温等离子体技术应用于粮食安全储藏领域提供参考。
1 材料与方法
1.1 稻谷样品
稻谷采自浙江省中央储备粮库中的粳稻谷,采用 3 层、5 点取样法,分别在粮仓东南角、东北角、西南角、西北角以及中央点作为取样点,每个取样点分别在距粮堆表层 0.5 m、中央层、距地面 0.5 m 取样。每仓 15 个取样点,样品放置在无菌采样袋中,于4 ℃保存。

图1 介质阻挡放电等离子体装置示意图
1.2 主要仪器与设备
介质阻挡放电等离子体装置,其示意图如图1,工作电压:20 kV。定制交流高压电源:GR-A-P30-150Ma型,MJ-系列恒温培养箱,T6新世纪可见分光光度计,JLG-Ⅱ砻谷机,LC-20A液相色谱仪,GCMS-QP201PLUS气相色谱质谱联用仪。
1.3 指标及测定方法
低温等离子体装置操作:将需要处理的样品放到培养皿中,再将其放到如图1所示的DBD处理室中,打开电源,将电压调到20 kV,处理相应的时间。
真菌的活化:草酸青霉(PenicilliumoxalicumACCC 32574)用马铃薯葡萄糖琼脂培养基(PDA),28 ℃培养5 d,5 ℃储藏。米根霉(RhizopusoryzaeCICC 3010)和黑曲霉(AspergillusnigerCICC 2041)用PDA培养基, 28 ℃培养5 d,5 ℃储藏。亮白曲霉(AspergilluscandidusACCC 30347)用014马铃薯葡萄糖琼脂,28 ℃培养5 d,5 ℃储藏。真菌的活化可参考《菌种活化操作方法》[14]。
稻谷常见真菌活性的测定:将用1%次氯酸钠溶液处理过的无菌干燥稻谷分为9组。取5 g无菌稻谷浸泡到等量含有真菌的生理盐水中,使其充分接触。再将9组稻谷分别进行0~8 min低温等离子体处理,然后放入装有45 mL生理盐水的蓝口瓶中超声洗去稻谷表面的真菌。选取合适梯度的稀释液100 μL到凝固好的平板培养基,按涂布法涂匀后用封口膜密封于28 ℃的MJ-系列霉菌培养箱中培养。记录菌落数量,计算抑菌率。
稻谷中蛋白质组分的测定:称取分别经过0~8 min处理的米粉各0.1 g,共9组,参考杨静等[15]的连续提取法,依次用蒸馏水、2%氯化钠、75%乙醇和0.04 mol/L氢氧化钠4种溶剂对清蛋白、球蛋白、醇溶蛋白和谷蛋白进行分离提取,然后采用考马斯亮蓝法依次对各组分蛋白含量进行测定。
稻谷中淀粉含量的测定:称取经过等离子体处理0~8 min的米粉各2 g,共5组,参照 GB 5009.9—2016《食品安全国家标准 食品中淀粉的测定》中的酸水解法进行测定。
稻谷脂肪酸组分的测定:分别取经过0~8 min处理过的米粉各2.5 g,共5组,放置于50 mL离心管中。加入25 mL石油醚,在42 ℃,300 W的条件下超声37 min后以4 000 r/min离心10 min。取上清液至圆底烧瓶中,于旋转蒸发仪上50 ℃旋蒸至无石油醚,烘干至恒重。加入0.5 mL 5 mg/mL十一烷酸甘油三酯标液(125 mg十一烷酸甘油三酯加25 mL甲醇)和25 mg焦性没食子酸。加入2 mL正己烷于圆底烧瓶中并转移到10 mL的具塞试管中,加入2 mL 0.5 mol/L的KOH-CH3OH溶液,充分振荡后于70 ℃水浴15 min,随后加入4 mL 0.5 mol/L的H2SO4-CH3OH在70 ℃水浴锅中水浴5 min,冷却至室温,加入2 mL正己烷和2 mL蒸馏水,加0.2 g NaCl,充分振荡后静置,取上层液体过膜,上机分析。
1.4 数据分析
采用Excel处理数据计算平均值和标准差,使用SPSS24和Origin2019b软件进行单因素分析和作图。

图2 不同处理时间下低温等离子体对对粳稻中4种霉菌的影响
2 结果与分析
2.1 稻谷真菌抑菌率
由图2可知,随着低温等离子体处理时间的增长,根霉、黑曲霉、草酸青霉和亮白曲霉的抑菌率总体上呈上升趋势。根霉在处理时间到达3 min后,其抑菌率与对照组相比显著提高(P<0.05);在处理时间到达8 min时,抑菌率可达到72%。这说明低温等离子体对米根霉的抑菌效果显著(P<0.05)。黑曲霉在处理时间到达3 min后,其抑菌率与对照组相比显著提高(P<0.05);在6 min之后,抑菌率基本保持不变趋向于平缓;在处理时间到达8 min时,黑曲霉的抑菌率可达到79%。这说明低温等离子体对黑曲霉的抑菌效果显著(P<0.05)。草酸青霉在处理时间到达2 min后,抑菌率与对照组相比显著提高(P<0.05);在处理时间到达8 min时,其抑菌率可达到64%。这说明低温等离子体对草酸青霉的抑菌效果显著(P<0.05)。亮白曲霉在处理时间到达2 min后,抑菌率与对照组相比显著提高(P<0.05);在处理时间到达8 min时,对亮白曲霉的抑菌率可达到66%。这说明低温等离子体对亮白曲霉的抑菌效果显著(P<0.05)。
随着低温等离子体处理时间的增长,低温等离子体对4种霉菌的抑菌率均有显著提高(P<0.05)。Niemira等[16]利用等离子体处理苹果时,也发现处理组的杀菌率随处理时间的增加而增加。Misra等[17]利用等离子体抑制樱桃表面的细菌,结果发现随着处理时间的延长,抑菌率逐渐提高。
2.2 粳稻蛋白质组分
稻谷中蛋白质含量与品种有关,含量高有利于大米的吸水膨胀和糊化,增加大米黏性,改善口感[18]。由图3可知,清蛋白、球蛋白和醇溶蛋白的含量一开始都先随处理时间而提高。清蛋白含量在2 min后便趋于平衡,不会随处理时间发生显著变化。而球蛋白含量在处理5 min后又随处理时间而增多。醇溶蛋白含量在2 min时达到最大值1 642.2 μg/g后又趋于平稳,直到7 min。这可能是因为低温等离子体中的高能电子和活性氧等物质破坏了稻谷细胞壁组成成分里的共价键实现对稻谷表面的蚀刻,使稻谷表面出现凹陷和裂痕[19,20],从而导致稻谷内部的清蛋白、球蛋白和醇溶蛋白更容易被提取出来。

图3 不同处理时间下低温等离子体 对粳稻蛋白质组分含量的影响
谷蛋白含量随处理时间先减少,到1 min后谷蛋白含量随处理时间趋于平缓,无显著变化。有研究表明,水稻种子的谷蛋白主要由分子质量为36 000和22 000两种亚基通过双硫键结合形成的[21],而低温等离子体对双硫键可能会有破坏作用[22]。虽然,谷蛋白含量也可能因为低温等离子体对稻谷表面的蚀刻作用而提高,但是低温等离子体降解谷蛋白的数量大于谷蛋白由于蚀刻作用而多提取出来的数量,总体上谷蛋白含量是下降的。醇溶蛋白含量在 7 min后降低(P<0.05),也可能是这一原因所造成的。另外,等离子体对二硫键的破坏作用,可以在一定程度上提高米饭的黏度,提升米饭的食用品质[23]。
2.3 粳稻淀粉含量
淀粉是稻谷中的主要成分,是影响稻谷食用品质的重要因素之一。图4为不同处理时间下粳稻的淀粉含量变化,与对照组相比,处理时间为4 min的淀粉含量无显著变化,而处理时间为6、8 min时,与对照组相比,粳稻的淀粉含量有所下降(P<0.05),这可能是因为处理时间过长导致部分淀粉被氧化分解。这说明适当的低温等离子体处理对粳稻的淀粉含量无显著影响,但是处理时间过长会导致粳稻淀粉下降(P<0.05)。并且结合抑菌率实验,可以发现跟对照组相比,在处理时间为4 min时,4种真菌的抑菌率均已显著提高(P<0.05)。所以只要控制合适的处理时间,就可以在不影响稻谷的淀粉含量的前提下,有效抑制稻谷霉菌生长。虽然高强度的等离子体处理会使淀粉含量降低,但同时也能改性淀粉的某些功能。比如Zou等[24]发现,等离子体可以使淀粉的羟基极化,在不破坏淀粉主体结构的情况下促进淀粉的交联。Thirumdas等[25]研究发现,经等离子体处理后,大米的蒸煮时间有效缩短,并且其吸水率提高。

表1 不同处理时间下低温等离子体对粳稻的脂肪酸组分的影响/%
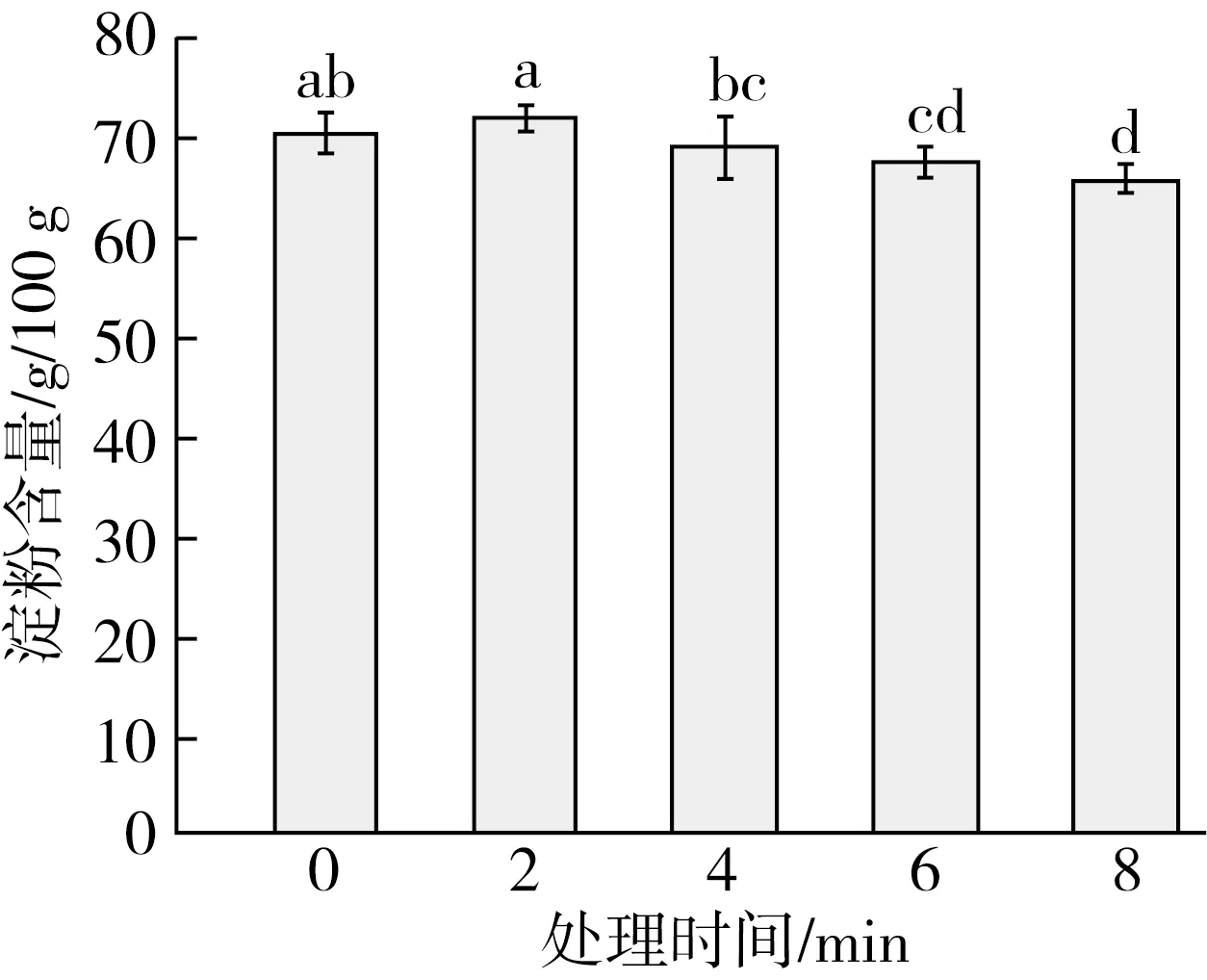
图4 不同处理时间下低温等离子体 对粳稻淀粉含量的影响
2.4 稻谷脂肪酸组分
稻谷自身的呼吸作用和一些环境因素的影响会使稻谷籽粒中的脂类物质水解而产生游离脂肪酸,这些游离脂肪酸不稳定很容易发生酸败现象从而影响稻谷品质,因此根据脂肪酸组分的变化可以衡量稻谷品质是否受到影响。并且不饱和脂肪酸对人体健康具有重要作用,如α-亚麻酸和二十碳五烯酸具有促进大脑发育、保护视力、降血脂和预防心血管疾病等多种生理作用[26]。对处理0、2、4、6、8 min时间里的稻谷进行脂肪酸组成成分的鉴定,经过谱库检索匹配,整理出了相似度较大的11种脂肪酸,如表1所示。经过低温等离子体处理4 min后,只有十五烷酸的含量与对照组相比略有提高(P<0.05)。但是随着处理时间的继续延长, 十五烷酸含量无显著变化。而低温等离子体处理对其余10种脂肪酸均无显著影响。尽管低温等离子体中存在羟自由基、超氧阴离子自由基和过氧化氢等物质,会对不饱和脂肪酸产生氧化作用[27],但是由表1可知,粳稻的亚麻酸、反-9-十八碳烯酸和反-11-二十碳烯酸这些不饱和脂肪酸的含量并未减少,甚至反-11-二十碳烯酸含量还会升高。产生这种现象的原因可能是等离子体在一定程度上促进脂肪的水解,但无法对粳稻内部的不饱和脂肪酸产生氧化作用。
3 结论
采用低温等离子体技术对根霉、黑曲霉、草酸青霉和亮白曲霉4种稻谷储藏中常见霉菌进行处理,4种霉菌的抑菌率随处理时间而显著上升(P<0.05)。粳稻经过低温等离子体处理后,除了十五烷酸,其他脂肪酸含量与对照组相比无显著变化;淀粉含量则有所下降(P<0.05);实验组的蛋白质组分中的球蛋白、醇蛋白和清蛋白相比对照组均略有上升,只有谷蛋白的实验组比对照组显著下降。适量时间的低温等离子体处理既能有效地抑制根霉、黑曲霉、草酸青霉和亮白曲霉等4种常见真菌的生长,并且能够较好地保证稻谷的品质,而长时间的处理则会破坏稻谷的食用品质。本实验只是初步探究了低温等离子体技术应用于稻谷霉菌控制领域的可行性。未来对于低温等离子体的抑菌机理还需更加深入的研究,如该技术对稻谷风味以及感官评价的影响,不同处理电压、不同氧气比例对真菌毒素的降解效果等仍需进一步的研究。
猜你喜欢
中国稻米(2022年5期)2022-10-11
汽车实用技术(2022年11期)2022-06-20
今日农业(2021年15期)2021-10-14
农民致富之友(2020年32期)2020-12-03
农村百事通(2018年24期)2018-02-22
农村百事通(2017年8期)2017-04-26
农村农业农民·B版(2017年2期)2017-03-11
科学启蒙(2016年11期)2016-12-09
国外科技新书评介(2014年8期)2014-12-05
