
喜马拉雅旱獭(Marmota himalay ana)栖息地选择
2022-10-24 02:06米玛旺堆
西南民族大学学报(自然科学版) 2022年5期
张 娟,米玛旺堆
(西藏大学,西藏 拉萨 850000)
动物在进行栖息地选择会受到生存条件和环境因子等综合因素的共同影响[1-2],主要分为躲避捕食者[3-4]、配偶和资源的竞争[5]、抚育幼崽[6]等一系列与觅食相互冲突的需求[7].种间竞争、食物资源、隐蔽级和捕食风险对动物的生境选择均会造成影响[8].
旱獭的洞穴由主洞穴、辅助洞穴和冬眠洞穴组成,冬眠洞穴主要用来冬眠[9],其他动物也会使用旱獭的洞穴,例如它们会借用蒙古旱獭(Marmota sibirica)的洞穴作为避难所,这些动物主要是一些小型的哺乳动物(如沙鼠属Meriones spp),鸟(翘鼻麻鸭Tadorna tadorna)和爬行动物(变色沙蜥Phrynocephalus versicolor和丽斑麻蜥Eremias argus)[10-12].
旱獭是一种体型较大、居住在地面、穴居的松鼠科动物,其扩散能力较弱[13-14],全世界一共有着14种旱獭,我国分布着4种,它们分别是喜马拉雅旱獭(Marmota himalayana)、灰旱獭(Marmota baibacina)、蒙古旱獭和长尾旱獭(Marmota caudate),其中数量最多的是喜马拉雅旱獭,并且它的分布范围最广、造成的危害也最大[15].喜马拉雅旱獭广泛分布在高寒草甸草原,据估计,人类疾病中有61%左右是人与动物共患疾病,其中,与兽类有关的病毒性人与动物共患疾病中,有大约60%来自鼠类[16],喜马拉雅旱獭作为青藏高原鼠疫的主要贮存宿主[17],针对其相关研究主要为鼠疫监测[18-19],病毒组的提取与鉴定等[20-21],此外还有一些研究者对其生态学特征进行了初步的探究,例如其洞穴的特征[22]、若尔盖湿地[23]和甘肃地区[24]喜马拉雅旱獭生境选择以及生活习性和种群密度的变化特征[25].国外一些研究者也对旱獭属其他种旱獭的生境选择做了研究,例如López等学者研究了法国南比利牛斯山脉阿尔卑斯旱獭(Marmota marmota)的大生境选择,结果表明阿尔卑斯旱獭选择小范围的钙质土壤和大范围的草地,并且它的洞穴的密度与湿地植被的存在着一种正相关关系[26].Turnock等学者研究了又称蒙塔古岛褐旱獭(Marmota caligata)的生境特征,发现蒙塔古岛褐旱獭可能会选择有灌木的地区[27].Věra等学者调查了阿尔泰山脉南部灰旱獭亚种的生境偏好,结果表明,其生境利用与植被类型有很大的相关性[28].由于青藏高原的特殊性,已有研究表明1990~2013年青藏高原气温呈上升趋势,气温增长率约为每10年0.5℃[29],气候变化对高山生态系统的影响比其他地区要快,这反过来又会影响高山特有物种的物候和种群分布[30-31],喜马拉雅旱獭在青藏高原的生境特征仍有待研究,研究喜马拉雅旱獭的栖息地特征也可以了解是什么决定了它们在陆地上的分布,这对它们的保护和鼠疫的监测也至关重要,为鼠疫的防控、监测和策略制定提供理论基础.
1 研究地点
研究地点位于西藏自治区山南市措美县哲古镇(91°43′23″E,28°43′23″N)以及拉萨市墨竹工卡县各乡镇(92°23′71″E,29°69′24″N),研究地点样方分布见图1.墨竹工卡县以高原宽谷和山地为主,草地类型随海拔高度的升高呈垂直地带性分布;植被类型由山地灌丛向高寒草甸过渡,海拔均在4 500 m左右,墨竹工卡县气候属于温暖半干旱气候,全年日照时间长、日温差大、雨期集中,风大风频、冰雹多、霜期长[32].年平均气温4.9℃,年均日照时数3 012 h,平均日照百分率为67.3%,无霜期为约100 d[33].措美县位于西藏的南部、喜马拉雅山北麓,平均海拔4 500 m.研究区域以高寒草甸为主,主要草本优势种为高山嵩草(Kobresia pygmaea),主要伴生种为矮生嵩草(Kobresia humilis)和狼毒(Stellera chamaejasme Linn)、和棘豆(Oxytropis kansuensis)等,灌木主要有毛叶蔷薇(Rosa mairei levl.)、爬地柏(Sabina procumbens)、雪山杜鹃(Rhododendron aganniphum Balf.f.et K.Ward)、西藏锦鸡儿(Caragana spinifera Kom.)、拉萨小檗(Berberis hemsleyana Ahrendt)、高山柳(Salix obscura Anderss)和金露梅(Potentilmaola fruticose)等.喜马拉雅旱獭的捕食者有藏狐(Velpes ferrilata)、狼(Canis lupus)、藏棕熊(Ursus arctos pruinosus).
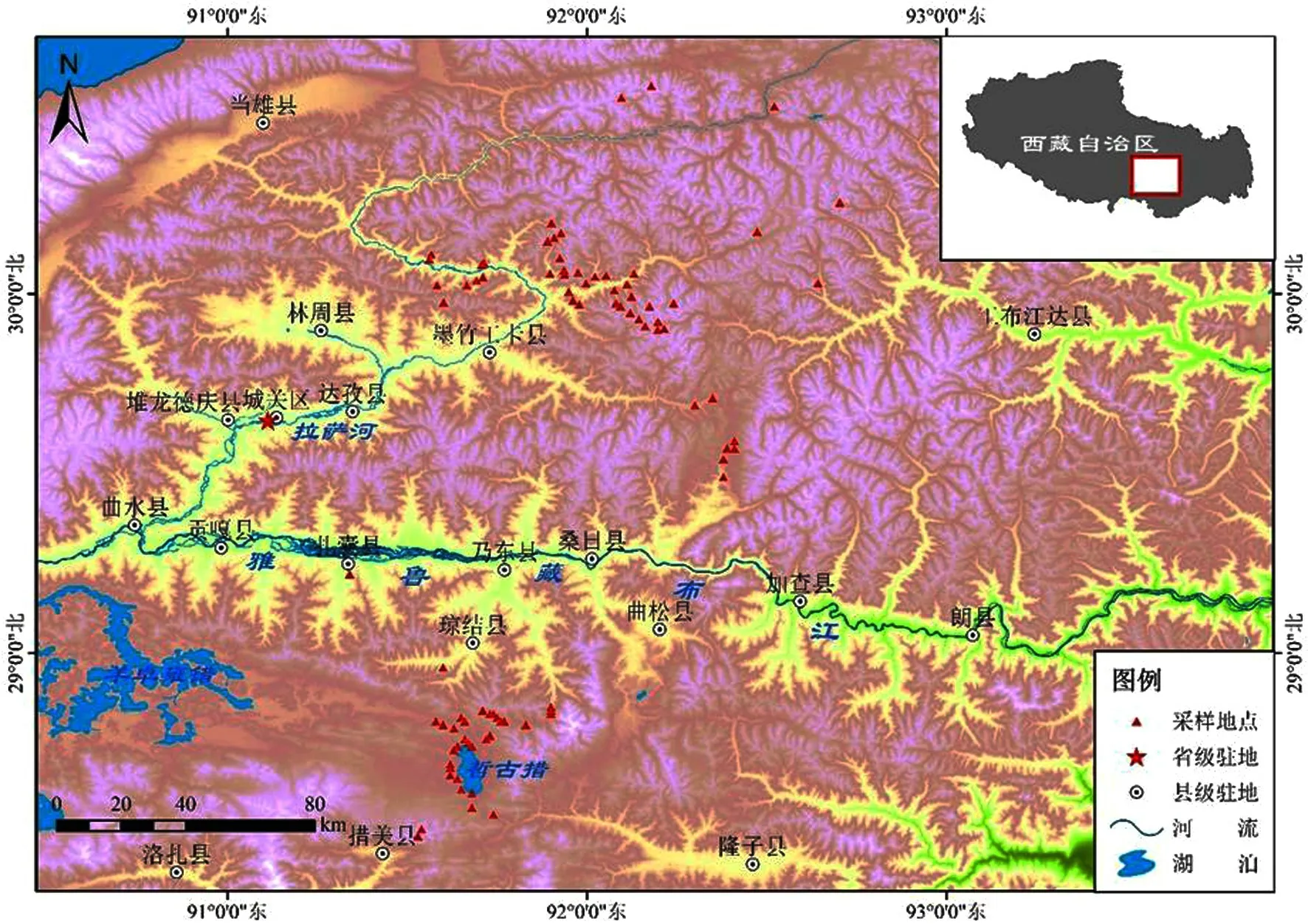
图1 研究样方分布示意图Fig.1 Distribution diagram of sample plot
2 研究方法
2.1 调查方法
野外数据采集时间为2021年9月-2021年10月,研究主要采用样方法,采取随机采样的方法,在采样地设置100 m×100 m的样方,每个样方间隔距离不少于2 km,在每个样方内再随机设置5个1 m×1 m的小样方来测量草本高度,在有喜马拉雅旱獭的区域设置利用样方,在没有喜马拉雅旱獭的区域设置对照样方,测定样方内的11种数值型生态因子和6种描述型生态因子,测定方法参照李路云[34-35]等的方法.测定的因子如下:
(1)海拔、经度和纬度.
(2)坡度:利用坡度测量仪记录样方中心的坡度.
(3)草本盖度:测算样方内草本的覆盖面积占整个样方面积的百分比.
(4)灌木盖度:测算样方中灌木树冠覆盖面积与地表面积的比例,取百分比.
(5)草本均高:利用卷尺测量5个1 m×1 m样方内随机10株草本的高度取平均值.
(6)灌木均高:利用卷尺测量整个样方内随机10株灌木的高度取平均值.(若整个样方内的灌木少于10株,则测量整个样方内全部的灌木高度取平均值)
(7)距水源的距离:样方中心到水源的距离.
(8)距干扰的距离:样方中心到人为干扰的距离.
(9)距公路的距离:样方中心到公路的距离.
(10)生境类型:根据实地调查,将生境类型划分为灌丛、草甸和湿地.
(11)人为干扰类型:根据干扰的类型划分为放牧,住房、农田、工厂和寺庙五种类型.
(12)隐蔽级:在样方中心设立一根60 cm(旱獭站立时眼睛的大概高度)的杆,样地4个方向的不可视距离的平均值/60 cm取百分数,0%-30%记录为弱,30%-60%记录为中,60%-100%记录为强.
(13)优势灌木:记录样方内数量最多的灌木.
(14)优势草本:记录样方内数量最多的草本.
(15)坡向:根据洞穴所在的坡面朝向,将坡向分为四种类型.阳坡、阴坡、半阴半阳坡和无坡向.
2.2 数据处理方法
所有的数据均在Excel 2016和R语言4.1.1软件包中整理和分析.其中描述型生态因子有坡向、生境类型、干扰类型、隐蔽级、优势灌木、优势草本6种,对它们进行卡方检验,对除此之外的11种数值型生态因子进行K-S正态检验,对于服从正态分布的因子进行独立样本T检验,对于不服从正态分布的因子先进行对数转换再做K-S正态检验,对于数据转换后还不服从正态分布的生态因子用使用Mann-Whitey U来检验利用样方与对照样方的差异.对喜马拉雅旱獭生境选择因子进行逐步回归分析,选择AIC值最小的建立逐步回归方程.
3 结果
3.1 差异性检验
此次调查共采集了87个样方,包括42个利用样方和45个对照样方.对喜马拉雅旱獭生境选择的17种生态因子分别进行差异性检验,其中6种非数值型生态因子进行卡方检验,另外11种数值型生态因子进行K-S正态检验.
11种数值型生态因子正态分布检验结果如表1所示,单个样本K-S正态检验表明,海拔、坡度和草本均高服从正态分布,对不服从正态分布的数据先对数转换再进行K-S正态检验,发现对数转换后距人为干扰距离服从正态分布,其余生态因子仍不服从正态分布.

表1 11种数值型生态因子正态检验结果Table 1 K-S results of 11 numerical ecological factors
卡方检验结果表明,利用样方与对照样方在生境类型(X2=7.987 5,df=2,P=0.018 43<0.05)、隐蔽级(X2=11.389,df=2,P=0.003 364<0.01)、优势灌木(X2=22.317,df=6,P<0.01)和坡向(X2=9.428 6,df=3,P=0.0241<0.05)4个生态因子上差异显著,在优势草本(X2=9.099 1,df=7,P=0.261>0.05),干扰类型(X2=4.201 5,df=4,P=0.395 2>0.05)2个生态因子上没有差异性.T检验和Mann-Whitey U Test检验结果表明(见表2),利用样方与对照样方在海拔、距人为干扰距离、坡度和草本均高、纬度、草本盖度、灌木盖度、灌木均高上有极显著差异(P<0.01),在经度、距水源的距离和距公路距离上没有差异(P>0.05).

表2 喜马拉雅旱獭栖息地选择样方与对照样方11个数值型变量生态因子的比较Table 2 Comparison of 11 numerical variables of ecological factors between marmota himalayana habitat selection used sites and controlled plots
3.2 单因素分析和逐步回归分析
通过对喜马拉雅旱獭洞穴数和各个数值型生态因子之间进行单因素分析和逐步回归分析.单因素分析结果(见表3)表明,海拔、纬度、坡度、草本盖度、灌木均高、距水源的距离、距人为干扰的距离7个生境因子具有显著差异性(P<0.05),说明喜马拉雅旱獭对这7种生境具有选择性.对生境因子进行逐步回归分析(表4),根据逐步回归结果,选择AIC值最小的建立逐步回归模型:海拔+经度+纬度+坡度+草本盖度+灌木盖度,F统计量6.862,并得到了极显著支持(P<0.001).
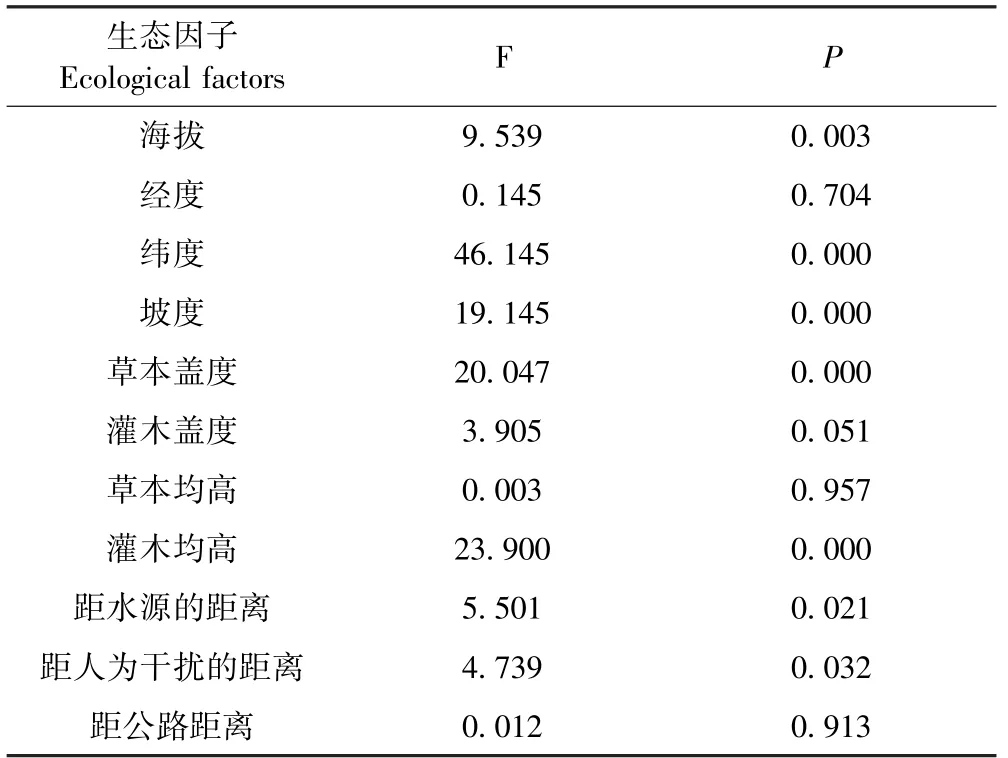
表3 喜马拉雅旱獭生境选择11种生态因子单因素分析表Table 3 Single factor analysis of 11 ecological factors for habitat selection of Marmota himalayana

表4 回归模型结果Table 4 Regression model results
4 讨论
本研究表明喜马拉雅旱獭秋季生境偏好适宜的海拔和纬度、坡度大约为10°~30°、有灌木覆盖、灌木高度大于70 cm(成年旱獭站立时的大概高度)的地带,距人为干扰较近、隐蔽级较高以及阳坡的区域,这与之前的研究[26,36]大致相同.
4.1 海拔和纬度对喜马拉雅旱獭栖息地选择的影响
喜马拉雅旱獭在生态学上不同于世界上所有其的旱獭,喜马拉雅旱獭的活动范围在海拔3000 m以上[37].全世界共有14种旱獭,全部分布在北半球,纬度对喜马拉雅旱獭造成的影响可能跟种间竞争有关,在我们的实地调查中喜马拉雅旱獭集中分布在墨竹工卡县,而在措美县分布较少,这可能和降水量有关.墨竹工卡县年降水量(522.1 mm)远大于措美县(286.5 mm).根据我们的野外调查发现在喜马拉雅旱獭聚居的地方几乎没有发现其他啮齿类动物的洞穴,如高原鼠兔(Ochotona curzoniae)等,在没有喜马拉雅旱獭的地带多为高原鼠兔的聚集地(图2),这可能和种间竞争有关,喜马拉雅旱獭和高原鼠兔都采集禾本科、菊科和玄参科植物[38].

图2 未发现喜马拉雅旱獭的样地(多为高原鼠兔的聚集地)Fig.2 No transects of Marmots himalayan were found(mostly the gathering places of plateau pika)
4.2 隐蔽级和灌木对喜马拉雅旱獭栖息地选择的影响
喜马拉雅旱獭对隐蔽级的偏好高>中>低(见表5),并且喜马拉雅旱獭的栖息地都有灌木覆盖或存在较大的岩石的区域.这可能是因为旱獭的栖息地偏好表现为进化上的保守,亲缘关系密切的物种倾向于占据相似的环境[39].Svendsen等学者的研究发现黄腹旱獭(Marmota flaviventris)的各种生境都有一个共同的特征就是有足够大的岩石提供遮蔽,并且旱獭洞穴在岩石下面,这些岩石被用来进行瞭望或者晒太阳[40],并且旱獭相对偏好草原与岩石混合生境,特别是在旱獭到达一个新领域的时候[41],这与我们的野外调查结果一致.喜马拉雅旱獭的洞穴也常在岩石下面(见图3),另外我们的野外调查还发现,喜马拉雅旱獭也偏好在灌木根下挖洞(见图4),这些也能说明喜马拉雅旱獭喜欢隐蔽级较高的栖息地.除此之外灌木可以积累更深的雪,而这额外的绝缘层可以增加冬天冬眠时的温度,提高冬眠存活率[26],并且这些雪在春季和夏季融化的更早[42],因为5月中旬,它们从冬眠中醒来,会立即在无雪的附近搜寻区域各种草本植物食用[43].我们的研究观察到喜马拉雅旱獭在灌木丛旁觅食,这种行为在蒙塔古岛褐旱獭也有被观察记录到[27],旱獭在觅食时可能会利用灌木来躲避捕食者[27].动物的反捕食根据能量的消耗程度可以分为三类:隐蔽、逃避、自卫[44],而喜马拉雅旱獭选择在岩石附近和灌木根下挖洞,可以有效地躲避捕食,提升存活概率.喜马拉雅旱獭几乎不选择没有毛叶蔷薇的地方(见表6),毛叶蔷薇是蔷薇科的一种矮小灌木,高1-2 m,密被针刺,这可能也是喜马拉雅旱獭选择它的原因,针刺有利于躲避捕食者,且灌木郁闭度较大,隐蔽级更高.喜马拉雅旱獭是藏棕熊、藏狐和狼等的食物来源之一,选择隐蔽级较高的区域可以有效躲避天敌,提高存活率.

图4 喜马拉雅旱獭洞穴在灌木根下Fig.4 Marmota himalayana burrow under shrub roots
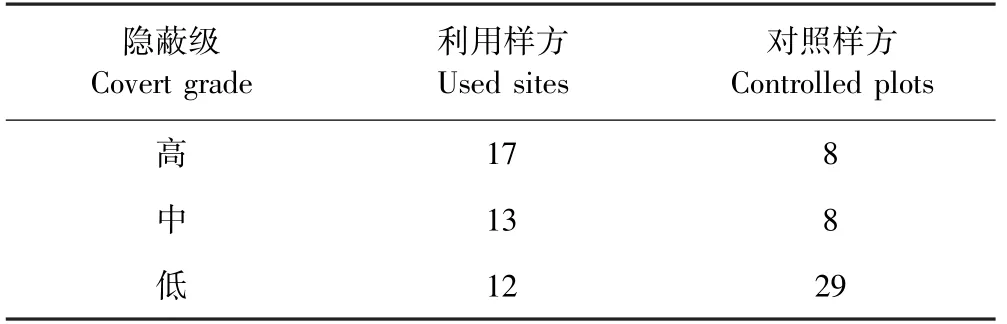
表5 利用样方和对照样方隐蔽级对比Table 5 The covert grades at used sites and controlled plots
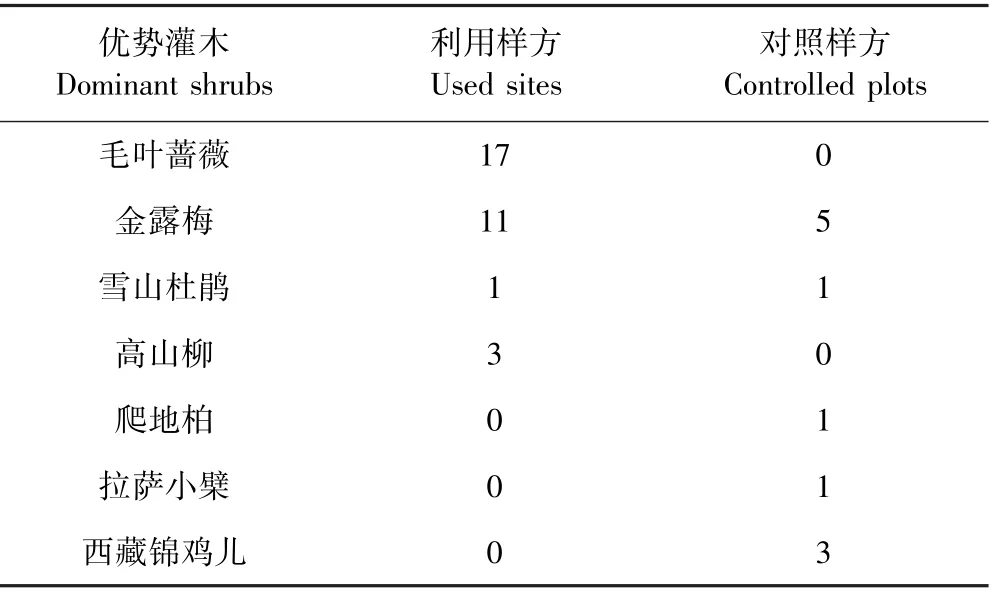
表6 利用样方和对照样方优势灌木对比Table 6 The dominant shrubs at used sites and controlled plots

图3 喜马拉雅旱獭洞穴在巨大岩石正下方Fig.3 A burrow of Marmota himalayana is directly beneath a huge rock
4.3 坡度和坡向对喜马拉雅旱獭栖息地选择的影响
喜马拉雅旱獭倾向于选择阳坡(见表7)以及有一定坡度的区域(见表8).喜马拉雅旱獭不选择坡度较低的区域是因为低坡会积累更深的雪[45],而太深的积雪会减少繁殖和同窝仔畜数,且旱獭也需要一些区域来提供冬眠醒来后的牧草[43,46-49],积雪的深度和积雪融化时间存在着显著正相关关系[50].喜马拉雅旱獭偏好阳坡区域这一点和阿尔卑斯旱獭很相似,因为阳坡是冬雪最早融化的地方[51],积雪的融化时间对旱獭的生存率和繁殖率有很大的影响.
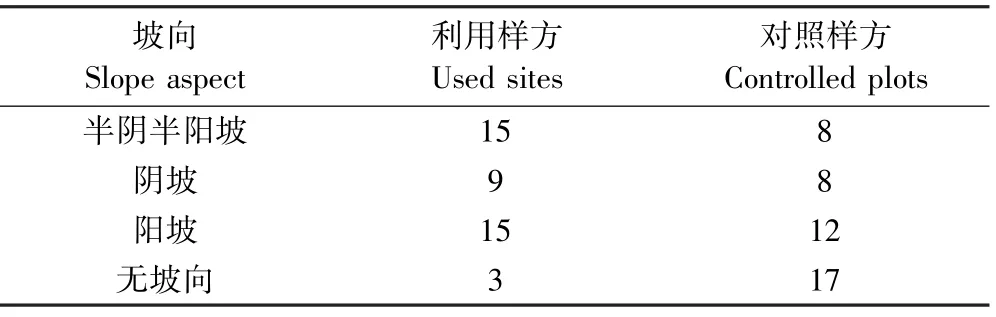
表7 利用样方和对照样方坡向对比Table 7 The slope aspect at used sites and controlled plots
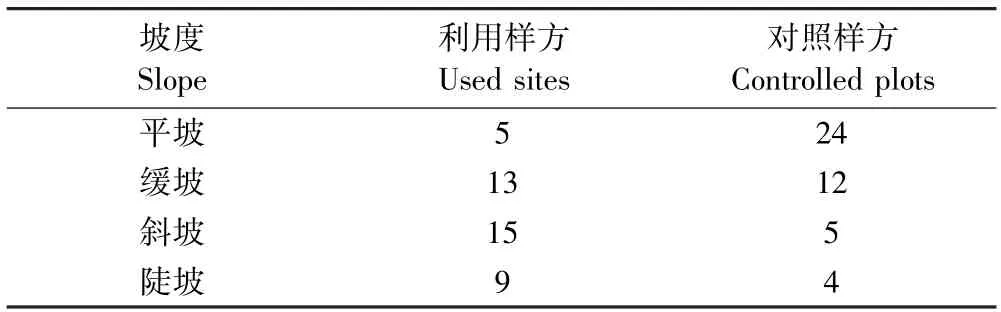
表8 利用样方和对照样方坡度对比Table 8 The slope at used sites and controlled plots
4.4 人为干扰和植物组成对喜马拉雅旱獭栖息地选择的影响
人为干扰对喜马拉雅旱獭的栖息地选择没有明显影响.Mainini等测验了在五种类型的徒步旅行活动下阿尔卑斯旱獭的行为,发现当只有徒步者时,以及狗(牵绳)和徒步者一起时旱獭不会躲在洞穴里,而狗不牵绳时,旱獭会躲在洞穴里,说明移动可预测性和潜在威胁是阿尔卑斯旱獭对不同形式徒步旅行反应的决定性因素[52].喜马拉雅旱獭偏好距人为干扰较近栖息地的原因可能为西藏主要以放牧为主,它们的生境又会和放牧地重叠,所以对人为干扰有了可预测性.此外郭程等人记录比较了若尔盖湿地三个同一生境类型下,但受到不同人为干扰的喜马拉雅旱獭种群,发现人类活动会提高喜马拉雅旱獭的繁殖率,因为受到人类活动的影响,旱獭会挖一些临时洞来作为避难所[53].国外研究者研究了黄腹旱獭的食物组成,发现黄腹旱獭偏好摄取禾本科植物(禾草、莎草和灯心草),尤其是三叶草(Trifolium andersonii)[54].Garin等研究者对阿尔卑斯旱獭的食性进行了研究发现它们更偏好豆科、菊科、百合科、车前花科和伞形花科的植物[55].国内的研究者对滇西北部喜马拉雅旱獭的食物组成进行了研究,发现喜马拉雅旱獭偏好摄食禾本科、杜鹃花科植物,其次为菊科、豆科、为菊科、豆科、龙胆科、百合科、莎草科等[36],在我们的研究样方中记录到的植物主要以莎草科、禾本科、菊科、豆科、蔷薇科、车前花科为主,说明西藏地区的喜马拉雅旱獭可能偏好摄食这些科的植物.
猜你喜欢
意林·少年版(2022年9期)2022-06-14
青年文学家(2021年16期)2021-08-05
小哥白尼(野生动物)(2021年10期)2021-02-12
少儿美术(2020年8期)2020-11-06
科普童话·百科探秘(2020年9期)2020-09-06
少年文艺(2020年6期)2020-06-11
好日子(2019年4期)2019-05-11
小哥白尼(野生动物)(2018年6期)2018-09-12
现代园艺(2017年13期)2018-01-19
小说月刊(2014年1期)2014-04-23
