
Structure complexity is the primary driver of functional diversity in the temperate forests of northeastern China
2022-10-18 01:59ZhenghuLinJunWngChunyuFnKlusvonGdow
Forest Ecosystems 2022年4期
Zhenghu Lin, Jun Wng, Chunyu Fn, Klus von Gdow
a Research Center of Forest Management Engineering of State Forestry and Grassland Administration, Beijing Forestry University, Beijing, 100083, China
b Department of Forest Ecology, School of Ecology and Nature Conservation, Beijing Forestry University, Beijing, 100083, China
c Faculty of Forestry and Forest Ecology, Georg-August-University G¨ottingen, Büsgenweg 5, D-37077, G¨ottingen, Germany
d Department of Forest and Wood Science, University of Stellenbosch, Stellenbosch, 7600, South Africa
Keywords:Functional diversity distribution Forest types Forest management Random forest Stand structure attributes Structural equation modeling
ABSTRACT
1. Background
Changes in climate conditions pose major threats to biodiversity(Poorter et al., 2017). Such changes can directly and indirectly affect ecosystem processes and functions through abiotic and biotic factors(Ayres and Lombardero, 2000; Ciais et al., 2005; Allen et al., 2010).Community ecologists are increasingly realizing that a clearer understanding of community diversity-based traits may potentially provide additional insights into the processes and functions of ecosystems (He et al.,2019).One such measure is functional diversity,which quantifies the diversity of a series of core attributes in a community that are closely related to plant growth,reproduction and survival.Functional diversity is fundamentally linked to potential niche differences as species occupy different niches based on their traits. Furthermore, it can significantly affect ecosystem function and reflect ecosystem response to environmental change(Roscher et al.,2012;Chiu and Chao,2014;Thom et al., 2019). Consequently, functional diversity is a good proxy for drivers of ecosystem processes and resilience (Petchey and Gaston,2002).
A wide array of indices has been developed to quantify functional diversity (Schleuter et al., 2010). However, simply adding species traits to describe their impact on the ecosystem cannot be assumed,as multiple species may perform similar roles due to functional redundancy(Hooper et al., 2005; Flynn et al., 2009). Here, we focus on a distance-based aggregate information index, known as the Functional Hill Number.The Functional Hill Number is a single-parameter (q) index that incorporates the species' pairwise functional distances calculated from species traits. It can be utilized to quantify the “effective number of equally abundant and functionally distinct species”in an assemblage,and allows the focus to be either on rare or on common species in evaluating functional diversity. For example, as q increases, the diversity values become more sensitive to common species. When q = 0, species abundance is ignored and it's the functional richness. When q = 1 is the exponential Shannon entropy, all species are weighted by their abundance equally. And q = 2 is the inverse of the Simpson concentration,common species get more weight than rare species(for details see Chiu and Chao, 2014; von Gadow et al., 2021). The abundance weighting method allows us to consider whether or not the species contribute to the ecosystem functions in proportion to their abundance,as proposed by the mass ratio hypothesis(Grime,1998).
Ecosystem functions provided by forests are threatened by direct and indirect effects of global change drivers (Hisano et al., 2018). The average global air temperature rose in the period of 1880–2012 accounting for 0.85°C(Kasprzyk et al.,2016),while precipitation patterns also changed significantly(Niu et al.,2019).These rapid climate changes have influenced community composition and species interactions often leading to decreasing ecosystem functioning,such as biomass production(Ma et al., 2017). In the past few years, the phenomenon of rising temperatures has spread throughout northeastern China; especially serious was changes in daily maximum temperatures. How ecosystems will respond to these changes depends on the functional traits of constituent species(Liu and Ma,2015).
Northeastern China,which spans the cold temperate zone,the middle temperate zone and the warm temperate zone, is known for productive forests(Qian and Ricklefs,2000).However,the low biodiversity inherent and convergent trait synthesis in the ecotone of temperate and cold temperate ecotones may be particularly susceptible to climate change as many species are at the limits of their climatic range(Jiang et al.,2021).Uncertainty about future environmental change makes planning for forest management exceptionally difficult.Approaches that can maintain or improve the ability of ecosystems to respond and reorganize rapidly after disturbance events are urgently needed (Mina et al., 2021). Forest management focusing on functional diversity provides a promising strategy for this situation(Messier et al.,2019).However,little is known about the distribution characteristics and drivers of functional diversity in northeastern China, especially how different forest types respond to these drivers.
Differences in species functional traits and life strategies often show great variation across a large geographical region (Zhang et al., 2014;Wang et al., 2015; Thakur and Chawla, 2019; Wieczynski et al., 2019;Aros-Mualin et al., 2021). Regions with high functional diversity imply high insurance effects and resilience after disturbance (Wüest et al.,2018). Thus, the integration of functional diversity into forest management plans to improve the resilience of ecosystems to environmental change is increasingly encouraged (Messier et al., 2019). From large to local scale,climate,human activities,topography and soil factors jointly determine the distribution of functional traits (Liu and Ma, 2015).However, as an important component of biodiversity, the relationship between stand structures (for example, stand density, variations in tree sizes, basal areas, and canopy complexity) and functional diversity is rarely understood. This is surprising since it has long been recognized that tree stands with complex and diverse structures can provide better ecosystem services and functions when compared with tree stands containing single and even-aged species(Ali et al.,2016;Ali,2019).
In this study, we analyze the functional diversity of forests in northeastern China. Our main aims are as follows: a) Mapping the functional trait diversity distribution patterns throughout the study region; b)Quantifying the drivers of functional diversity.As our study area spans a large latitudinal gradient and is composed of temperate forests, we hypothesized that environmental filtering by climate determines the distribution of functional diversity. Following the stress dominance hypothesis, the effects of environmental filtering will be most distinct under harsh conditions, while stability selection causes only adaptive species with similar traits to be established.As environmental conditions become more favorable, competitive interactions play increasingly important roles in determining species establishment (Weiher and Keddy,1995;Coyle et al.,2014).More specifically,we speculate that the trait diversity distribution would show a distinct north-south gradient,with more diversity in the southern Longgang Mountains.In accordance with the mass ratio hypothesis, we further assume that the dominant species determine the bulk of functional diversity distributions.We also expect that our results will show only moderate variation when weighting functional diversity by different species aggregation levels.
2. Methods
2.1. Study area and forest plot network
The study area covers a total of eight mountain ranges and includes cold temperate coniferous forests, temperate mixed coniferous broadleaved forests, and warm temperate deciduous broad-leaved forests in northeastern China (latitude 39°42′48′′–53°19′21′′N, longitude 119°48′1′′–134°01′0′′E;Fig.S1).Vegetation zone was delineated around areas sharing similar climate,topography,and soil substrate conditions.The main precipitation period in the study area is from June to September,which belongs to the contemporary rain-energy climate.
A network of 442 0.1-ha circular(radius=17.84 m)forest plots were established in the region during the summer of 2017 (Fig. 1). All individual trees with DBH (diameter at breast height, 1.3 m above ground)greater than 5 cm in the sample plots were identified,geo-referenced and tagged.
2.2. Drivers of functional diversity
2.2.1. Climatic conditions
A growing number of studies have indicated the potential impacts of changes in climate conditions on ecosystem functional diversity(Lavorel and Garnier,2002;Thuiller et al.,2006;Ordonez et al.,2016;Wieczynski et al., 2019). Our plot-level climate variables, are four important indicators of the vegetation zones: mean annual temperature; maximum temperature during the warmest months;annual precipitation; and precipitation during the driest months(1970–2000 observation period),all derived from WorldClim data at a 1 km × 1 km spatial resolution (Fick and Hijmans,2017).
2.2.2. Topographical conditions
The topography was usually identified as an easily measured proxy for local environmental resource availability (Fl′avia et al., 2005).Topographical conditions may influence plant performance by modulating microhabitat resources(Balvanera et al.,2011).In this study,all of the topographic factors were measured in-situ.The altitude was recorded using a hand-held GPS device, and the slope and aspect were measured using a compass.
2.2.3. Soil conditions
The geographical distance between plots may form diverse soil properties. For example, the plots near the valleys generally had higher nutrient levels than the plots near the ridges(Segura et al.,2002;Gibbons and Newbery,2003).It is well known that low soil nutrient levels result in low functional diversity (Liu et al., 2014). We extracted three soil indicators: organic carbon content, base saturation and carbonate equivalent content from the world soil property estimates for broad-scaled modelling(WISE30sec;Batjes,2015).
2.2.4. Forest types
The northeastern forests contain China's largest percentage of forest coverage and are characterized by a high diversity of forest types(Zheng et al.,2001).In accordance with the volume ratios of the tree species,the sampling plots were divided into the following five types: pure broad-leaved forests; pure conifer forests; mixed broad-leaved forests;mixed conifer forests; and mixed conifer and broad-leaved forests (Wu et al.,2019).
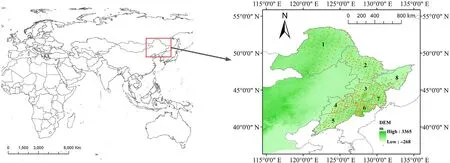
Fig. 1. Locations of the 442 examined forest plots in northeastern China, in which the study region covered eight mountain ranges indicated by the following numbers: 1, Great Khingan Mountains; 2, Lesser Khingan Mountains; 3, Zhang Guangcai Mountains; 4, Hada Mountains; 5, Longgang Mountains; 6, Changbai Mountains; 7, Laoye Mountains; 8, Wanda Mountains.
2.2.5. Stand structural complexity
The stand structures are typically characterized by variances in tree diameters at breast height (DBH) and the total tree heights (here the standard deviation) among and within the various species (Ali et al.,2016). Also, it was observed that the densities of the tree stand varied greatly among the different plot.The most common three stand structural attributes were obtained in-situ for each plot in the study region.
2.2.6. Anthropogenic factors
Climate change and human activities are two important drivers of ecosystem services and functions (Maeshiro et al., 2013). With the continuous warming of global climate, as well as the intensification of unreasonable use of land resources, such as overgrazing and land reclamation, ecosystem services and functions are undergoing dramatic changes (Huang et al., 2016). Four human footprint and roadless area indices (Table S1 for summary statistics) were derived from the human footprint database(Venter et al.,2016)and the global roadless database(Ibisch et al., 2016), with a resolution of 1 km, for the purpose of assessing their potential impacts on functional diversity.
2.3. Statistical analysis
Since functional traits are based on species level, more than three individual species were selected in order to make them representative.Ultimately, we collected 33,708 individuals from 54 species and measured their functional traits. The set of traits (Table S2) which was measured had been shown to be closely linked to growth and to have a significant impact on ecosystem functions (Chiang et al., 2016; van der Sande et al.,2017).
The functional diversity within each plot was quantified by the Functional Hill Numbers(Li,2018).The mapping of functional diversity distributions was completed using ordinary Kriging interpolation in ArcGIS 10.6. The spatial interpolation accuracy was evaluated by deriving the root-mean-square standardization of the predictions of the study plots. In addition, a random forest (RF) modeling approach was applied to evaluate the importance of functional diversity drivers. We derived 90% of the training set from all the obtained plot data using a bootstrapping method and evaluated the out-of-bag (OOB) errors with the remaining 10%. The model's goodness-of-fit (Table 1) was represented by three indicators: the coefficient of determination (R2), the root-mean-square error (RMSE) and mean absolute error (MAE). The relative importance of the variables was derived from 20 models. The random forest analyses were calculated using RandomForest Package(Liaw and Wiener,2002) in R 4.0.3(R Core Team,2020).
We constructed a conceptual structural equation model (Fig. S2)based on the importance ranking of variables. The first three most important variable groups were selected: stand structure, climate, and forest type(Fig.S3).In order to simplify the model path and improve the goodness of fit, PCA analysis was performed on four climate variables.Finally, climate variables are represented by the first axis of PCA(Fig.S4).Then,piecewise structural equation modeling was used to test whether the climate effect on functional diversity was mediated by forest structure.And we also examined whether or not the fixed factors(such as climate and stand structures)or random factors(such as the forest types)could explain the higher functional diversity variations.The Fisher C and p-values were used to evaluate the model-fit for each pSEM.Then,based on the conceptual model (Fig. S2), directional-separation tests were applied to check for any missing significant paths and whether or not unnecessary insignificant paths need to be excluded. The analysis of pSEM model was performed using piecewiseSEM packages (Lefcheck,2016)in R 4.0.3(R Core Team,2020).Linear SEM was applied to assess each forest type based on the same conceptual model.Each model-fit was evaluated using the following four statistical indexes: comparative fit index (CFI); goodness of fit index (GFI); root mean square residual(SRMR); and AIC. The scaled standardized path coefficients (ranging between-1 and 1)were then calculated and the significance(p <0.05)of the direct and indirect effects assessed(Fig.5;Fig.S6).The analysis of linear SEM was performed using Lavaan package (Rosseel, 2012) in R 4.0.3(R Core Team,2020).All of the above-described processes for three Functional Hill Numbers (q = 0, 1, 2) was calculated to determine the relationships between the predictors and the functional diversity changes as the weight of the abundant species increases.
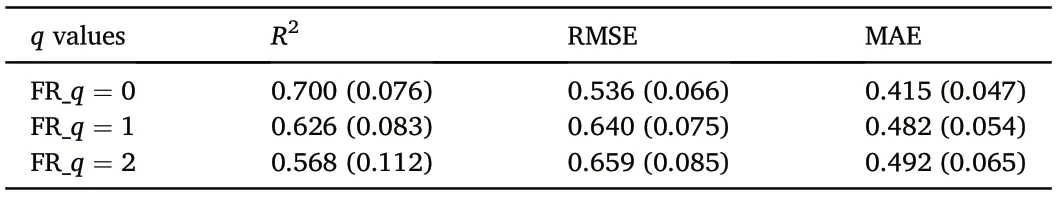
Table 1 Random forest model cross-validation results.Means and standard deviations(in parentheses) of each Hill numbers(q factor) are shown.
3. Results
3.1. Functional diversity hotspots in middle temperate forests
Our spatial analysis revealed that the temperate coniferous and broadleaved mixed forest zone includes several functional diversity hotspots, with Changbai Mountains being the most prominent. The average effective numbers of tree species were 8.4 (q = 0), 5.9 (q = 1),and 4.9 (q = 2) across the zone. Conversely, the Great Khingan Mountains were functional diversity "coldspots", where the average effective numbers of tree species were only 3.1(q=0),2.8(q=1),and 2.7(q=2).Interestingly, the average effective numbers of tree species in the warm temperate deciduous broad-leaved forests ranged between the above two extremes at 7.0 (q = 0), 5.2 (q = 1), and 4.7 (q = 2). Overall, with increasing q factor,the effective number of tree species decreased across the entire study region(Fig.2).The root-mean-square standardized of the spatial interpolations across all the sampling plots was 0.95(q=0),0.93(q=1), and 0.93(q= 2).
3.2. High correlation between forest structure and functional diversity
For each Functional Hill Number, the average value of 20 models showed that the five most important variables that explained the variations in functional diversity were the tree height and DBH diversity,the annual precipitation, annual average temperatures, and forest types(Table 2; Fig. S3). Slight shifts in the importance and ranking of some explanatory variables were found as the q factor increased.Overall,forest structure (36.6%–44.7%) was the most important predictor group for explaining the functional diversity variations. That was followed by climate (25.1%–30.9%), forest type (5.2%–10.8%), topography (8.7%–9.7%),human activities(7.3%–9.1%),and soil type(4.6%–5.9%;Fig.3).
3.3. The response of functional diversity to environment varies with forest types
The pSEM explained 0.18%(q=0),0.15%(q=1),and 0.13%(q=2)of the marginal variances(explained by climate and stand structure)and 0.31%(q=0),0.31%(q=1),and 0.26%(q=2)of the random variances(forest types as random effects) of the functional diversity, respectively(Fig.4).More specifically,functional diversity increased with increasing stand density(β(q=0)=0.27;β(q=1)=0.18;β(q=2)=0.15),tree height(β (q= 0)= 0.21;β (q=1) =0.20;β (q=2) =0.19),and DBH diversity (β (q = 0) = 0.22; β (q = 1) = 0.23; β (q = 2) = 0.21),respectively.However,stand density had significantly negative effects on the variations of tree height(-0.13)and DBH(-0.37).
The annual mean temperature and annual precipitation were found to have significantly positive effects on the functional diversity. Their effects on height diversity were significantly negative. However, we did note that the forest types explained the higher variances of the stand density (0.04), tree height (0.15) and DBH (0.26) variations, which in turn may potentially affect the functional diversity(Fig.4).
In order to determine whether the relationships tested in the pSEM(Fig.4)were dependent on the forest types,we assessed the linear SEMs across five temperate forest types(Fig.S5;Tables S4–S8).Compared with the pSEM results,the stand density and tree DBH variations had negative yet insignificant but direct effects on the functional diversity in the pure conifer forests. However, positive direct effects were observed in the other forest types. Furthermore, with the exception of the height variations of the mixed conifer forests (q = 1 and q = 2), which had insignificant negative effects on functional diversity, the other forest types experienced positive direct effects.
Consistent with the pSEM results, climate had positive direct effects on the functional diversity across all of the forest types. However, in comparison with the pSEM, climate had negative effects on the stand density in broad-leaved forests and mixed conifer and broad-leaved forests, with the changes ranging from significant to insignificant effects.But climate had positive effects on the stand density in the conifer forests (including pure and mixed conifer forests), including significant and insignificant effects (Fig. S5). The effects of climate on tree height diversity change from significantly negative to insignificant effects across all the forest types, which was consistent with the pSEM results. In addition,we noted that the direct effects of the predictors on functional diversity were higher than the indirect effects across the various forest types(Fig.5;Fig.S6).Moreover,the direct and indirect effects showed a variety of patterns across the forest types, with the exception of the effects of climate (both negative) on the functional diversity in mixed conifer forests (q = 1 and q = 2). The boxplots showing the differences amongst the fixed factors (for example, the predictors) across the five temperate forest types are presented in Fig.6.

Fig.2. Observed functional diversity distribution across northeastern China:(a)Functional richness (q = 0); (b) Exponential Shannon entropy (q = 1); and (c)Inverse of the Simpson concentration(q=2).Kriging interpolation was used to derive the overall distribution of Functional Hill Number based on the seven traits of 54 species in 442 plots;Three different q factors were used to derive the effects on the functional diversity of the different species abundance weights.

Table 2 Relative variable importance for explaining the variations in functional diversity.Relative importance indicates the relative contribution of each variable explaining functional diversity (adding up to 100%); the values were averaged across 20 models for each Hill Number(q factor);standard deviations are shown in parentheses (see Table S1 for the summary statistics of the explanatory variables).
4. Discussion
4.1. Environmental filtering is less important than forest structure
We found that there were obvious regional differences in the distribution of functional diversity.The lowest was found in the Great Khingan Mountains, where coniferous forests dominate, while the highest was found in the Changbai Mountains, where coniferous and broad-leaved mixed forests are dominant. Forest structure was the main driver of functional diversity distribution at regional scale. Our pSEM analysis showed that the indirect effect of climate on functional diversity was only mediated by height diversity,but was negative.The strong evidence of a direct correlation between forest structure and functional diversity was surprising. These results challenge our initial expectation that environmental filtering determines the distribution of functional diversity in the temperate forests in northeastern China. As described by the stressdominant hypothesis, functional diversity in temperate forests is generally low at the local scale compared to random simulations. This is because adaptive species with convergent trait syndromes are selected for community assembly by abiotic factors in harsh environments.Divergent trait syndromes,however,result from competitive interactions in affluent environments.Consequently,our results challenge the stressdominance hypothesis.As expected,the greater Khingan Mountains have the lowest regional functional diversity,while Changbai Mountains(not Longgang Mountains), have the highest.
In pSEM analysis,the high variance interpretation(random effect)of forest type on functional diversity verified this pattern. The unique climatic conditions in northeastern China have resulted in a variety of forest types. Although there were no significant differences in structural attributes among different forest types, it was obvious that the functional diversity of coniferous and broad-leaved mixed forests was significantly greater than that of coniferous and broad-leaved pure forests(Fig.6).The essence of this difference is niche differentiation. In contrast to monocultures, mixed-species forests with complementary architectural traits provide increased functional diversity due to the full use of light resources (Pretzsch, 2014). For instance, the canopy expands greatly and the understory light transmission increases in the mature forest stage,which is conducive to the growth and development of a sub-canopy layer and understory vegetation. The full utilization of ecological niche space by species is likely to cause an increase in functional diversity (Shen,2001). Although environmental filtering is less important than forest structure,its modest indirect effect on functional diversity suggests that climate may modify the structural development of forests.
The dynamic development process of forest structure,however,is not only affected by climate, but also by management practices and natural disturbances. In the past century, forests in northeastern China have undergone three historic management changes: harvesting without cultivation (1860–1945), timber utilization after the founding of the People's Republic of China (1950–1977) and timber utilization and the birth of ecological construction(1978–1998), followed by the period of ecological conservation and construction (Yu et al., 2011). Natural disturbances are an integral part of forest ecosystem dynamics and play an important role in shaping forest structural complexity (Janda et al.,2017). For instance, the Greater Khingan Mountains is known to be an ecotone of temperate and cold temperate climate zones and belongs to a transition region between monsoon and non-monsoon conditions.Due to the inter-annual instability of monsoon conditions, the agriculture and forest ecosystems in the region are often severely affected by such events as flooding, drought, frost, and cold damage during the growing season(Jiang et al., 2021). Small scale disturbance can promote structural complexity by creating favorable conditions for the growth of understory trees (Franklin et al., 2002; Willim et al., 2019). However, large-scale disturbances, such as fires or storms, can simplify the forest structure(Stephens and Gill,2005).Obviously,the large-scale disturbance effect is stronger in this region.

Fig.3. Relative importance of the stand structure,climate,forest type,topography,soil,and human activities for functional diversity:(a)Functional richness(q=0);(b)Exponential Shannon entropy(q=1);and(c)Inverse of the Simpson concentration(q=2).Boxplot shows the median(gray circles),10 percent(red bar),and 90 percent(blue bar)quantiles of the density distribution of the variable importance for each category;The three panels weight the different species abundance effects on the drivers of functional diversity. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
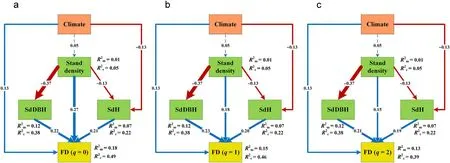
Fig. 4. Piecewise structural equation models (pSEMs) for linking the climate, stand density, tree height, and diameter variation and the functional diversity in the temperate forests of northeastern China,while considering the forest types as random effect:(a)Functional richness(q=0);(b)Exponential Shannon entropy(q=1);and (c) Inverse of the Simpson concentration (q = 2). Model-fit: a) Fisher's C= 1.289, p = 0.525, AIC = 41.289; b) Fisher's C = 1.289, p = 0.525, AIC = 41.289; c)Fisher's C=1.289,p=0.525,AIC=41.289.Blue and red arrows represent significantly positive and negative paths,respectively(p <0.05),while the dashed arrows indicate insignificant paths (p >0.05); Climate is the first principal component axis representing annual average temperature and annual precipitation (Fig. S4); for each path,the standardized regression coefficient is shown(See Table S3 for detailed statistics);the three panels weight the different species abundance effects on the drivers of functional diversity. (For interpretation of the references to colour in this figure legend, the reader is referred to the Web version of this article.)
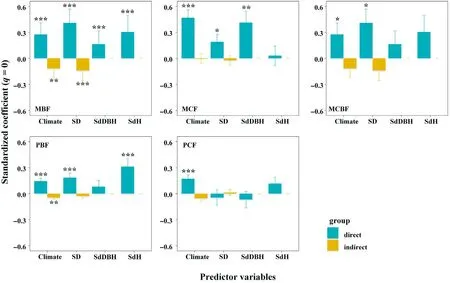
Fig. 5. Comparison of direct and indirect effects derived from the SEMs (see Fig. S6 and Tables S4–S8) of the climate, stand density, tree height, and tree diameter variations on the functional diversity within each temperate forest type in China.MBF is the abbreviation of the Mixed Broadleaved Forest;MCF is the abbreviation of the Mixed Conifer Forest; MCBF is the abbreviation of the Mixed Conifer Broadleaved Forest; PBF is the abbreviation of the Pure Broadleaved Forest; and PCF is the abbreviation of the Pure Conifer Forest; Data shows standardized coefficient ± standard error.
Our analysis was based on field surveys at specific points in time,however, time series data were needed to examine the relationship between functional diversity and stand structure across forest development(Pretzsch et al., 2016; Brandt et al., 2017; Meigs et al., 2017; Ali, 2019).Especially in the context of ongoing climate change,functional diversity is likely to decline with biodiversity loss due to their inherent correlations(Naeem and Wright,2003).However,the positive effect of temperature on stem biomass may(Reich et al.,2014)also contribute to forest structural complexity. In this case, the relationship between forest structure and functional diversity may be complex and context-dependent.Exploring the relationship on a time scale may go beyond our findings.
In addition,the results of this study showed that soil contributed the lowest to the importance of functional diversity,which may be due to the poor precision match between soil attribute values evaluated by broadscaled modelling approach and the actual plot. Soil attribute measurements from field may get different results, so future research is worth paying attention to.
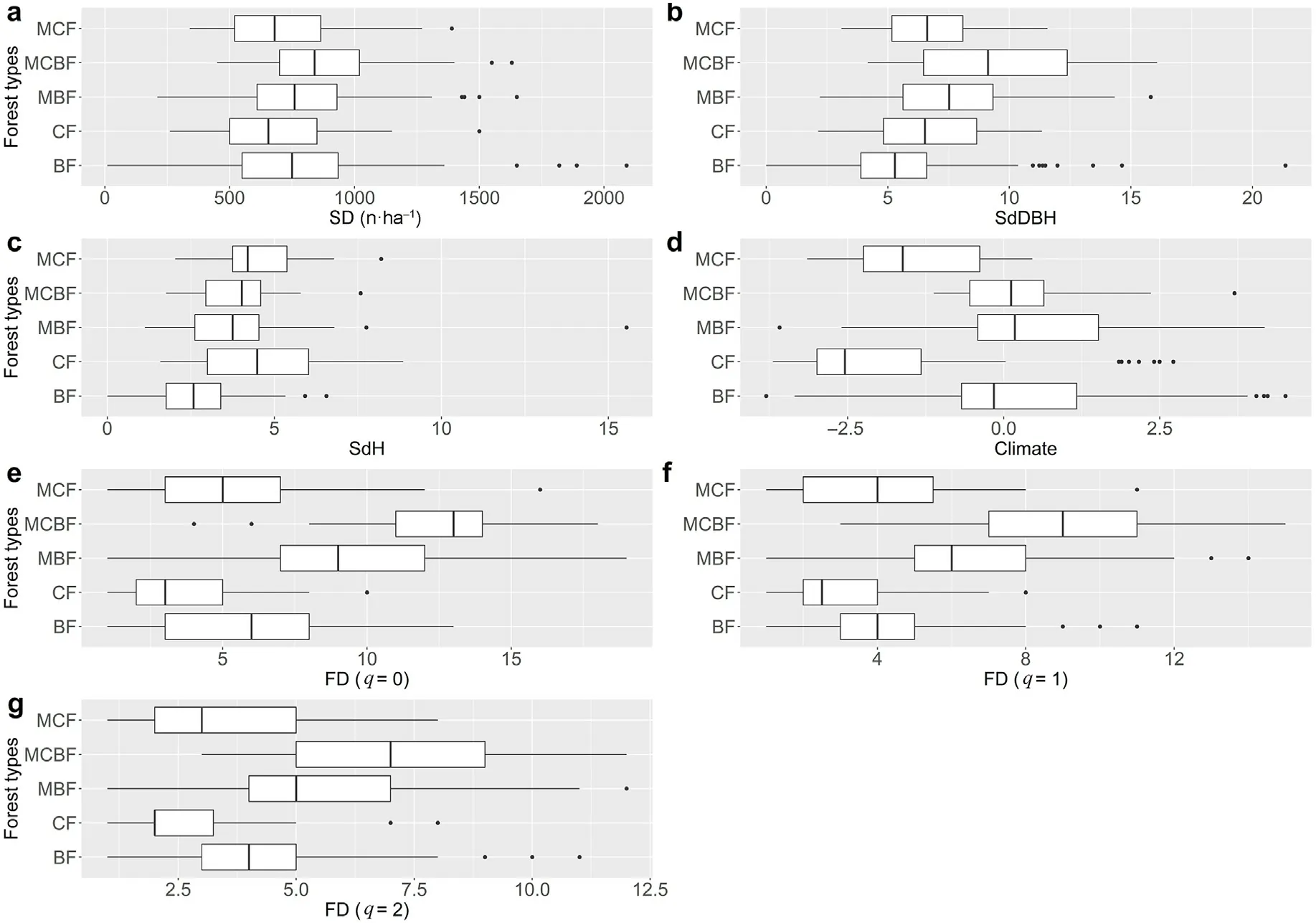
Fig. 6. Boxplots of the predictor and response variables used in structural equation models across the temperate forest types. The medians are significantly different where the boxes (95% confidence interval of the median) do not overlap.
4.2. Abundant species affect most of the functional diversity
All of the results had remained robust across the three q factors in the study region. It was found that the effective numbers of functionally of distinct species decreased with the increases in the q factors by up to 39%(Fig.2).Specific to each forest type(with the exceptions of the coniferous and broad-leaved mixed forests),the declines in the effective numbers of functionally of distinct species were even more subtle. Obviously, the drivers and distribution patterns of functional diversity showed only slight divergences across the species abundance weightings (Fig. 2;Fig. S3). Those results support the predictions of the mass ratio hypothesis (Grime, 1998), in which rare species have only slight effects on functional diversity, while abundant species determine most of the functional diversity.This was indeed found to be the case in northeastern China.
In this context, we speculate that there are varying degrees of functional redundancy within the study region,due to regional differences in the distribution of functional traits.For instance,the Changbai Mountain region is resistant to biodiversity loss,and that random loss of species will reduce the functional traits diversity to a fairly small degree, since suitable environmental conditions can support the same values and varieties of functional traits (Fonseca and Ganade, 2001; Mayfield et al., 2010).Our study only considered growth-based traits, but recruitment and survival are also important components of demographic performance.Studies have shown,for example,that climate change is already having an impact on the regeneration and survival of species(Shao et al.,2019).Thus, the set of traits related to recruitment or survival may lead to different Hill numbers and thus different conclusions.
4.3. Forest resilience management - a strategy aiming at functional diversity
Functional diversity is intrinsically associated with potential niche differences. Forest management aiming to maintain functional diversity may thus ensure a sustainable provision of multiple ecosystem goods and services(Messier et al.,2015;Sasaki et al.,2015).Forest management to increase functional diversity should focus on three related options.First,the contribution of species to ecosystem functioning is related to their stress tolerance and biological characteristics(individual species traits).In addition, functional groups, groups of species with a combination of biological characteristics that affect ecosystem function in similar ways,can be used to evaluate ecosystem functioning. Last but not least,structural complexity is a simple and easily manipulated measure that can be used directly to improve the functional diversity.
Our results show that the Greater Khingan Mountains region has a low trait diversity and it is more sensitive to climate change (Fig. 2;Fig.S5).This means that the region's ecosystems may be less resilient to and recover from disturbance. Developing appropriate forest management strategies based on functional diversity is a matter of urgency(Oliver et al., 2015, 2016; Hodgson et al., 2015; Ingrisch and Bahn,2018).
Resistance is related to the ability of a species or function to resist change in the face of disturbance, which is both related to survival and the ability of a species to continue to perform its function under stress(Oliver et al., 2015). Resistance occurs during disturbances, so individual-level traits that control species sensitivity to disturbances are important for developing resistant ecosystems (Gladstone-Gallagher et al., 2019). Consequently, according to the regional species pool, the tree species that can adapt to the corresponding environmental pressure are selected as the target trees for management. For example,cold-tolerant species such as Betula platyphylla Suk., Quercus mongolica Fischer ex Ledebour and Populus davidiana Dode are known to resist low temperature disturbances in northeastern China. Disturbance generally does not occur unilaterally. For example, low temperature is usually accompanied by frost, and high temperature may cause drought or a large number of diseases and insect pests to occur and spread. Furthermore, mixing different functional groups provides a powerful way to improve the sustainable trait diversity (T¨ornroos and Bonsdorff, 2012;Villn¨as et al.,2018).In addition,rapid recovery after disturbance is also one of the important goals for the development of a resilient ecosystem.Population attributes that control colonization at the landscape scale are important determinants of rapid recovery (Ingrisch and Bahn, 2018).Species with a range of traits associated with growth, reproduction and survival can achieve this goal.For instance,high resprouting ability after disturbance, fast juvenile growth or low light compensation points are desirable species characteristics for resilient ecosystems.
The application of the proposed measures in practice may still be challenging. The maintenance of ecosystem functions at different scales(stand or landscape scale) is limited in different contexts (such as the spatial range, frequency and intensity of disturbance). As a result, definition of these restrictions in the future will be beneficial to forest managers (Derose and Long, 2014). Fostering structural complexity is a straightforward way to improve functional diversity without much ecological knowledge.Based on our study,species mixing,and increasing the structural complexity of coniferous and broad-leaved mixed forest,has a strong positive effect on functional diversity.This can be achieved through a series of silvicultural methods applied in "near natural" and"continuous cover"forest management systems(Schütz,2002).A variety of niches created by a mixed forest structure depends on related physiological traits such as shade tolerance and crown plasticity in different functional groups and thus drives functional diversity. Our results may encourage forest managers in the region to focus on ecologically meaningful and silviculturally achievable objectives.
5. Conclusion
Changes in climate conditions often pose major threats to biodiversity. How ecosystems will respond to these changes depends on the functional traits of constituent species.Moreover,functional diversity is linked to potential niche differences as species occupy different niches based on their traits.Thus,it can significantly affect ecosystem function and reflect ecosystem response to environmental change(Roscher et al.,2012;Chiu and Chao,2014;Thom et al.,2019).Consequently,functional diversity is a good proxy for drivers of ecosystem processes and resilience(Petchey and Gaston, 2002). However, little is known about the distribution characteristics and drivers of functional diversity in northeastern China, especially how different forest types respond to these drivers.These gaps in knowledge hinder forest management based on functional diversity to improve the resilience of ecosystems to environmental change.
Here,we focus on a distance-based aggregate information index,and allow the focus to be either on rare or on common species in evaluating functional diversity, known as the Functional Hill Number. Our study deals with a topical issue: Assessing the spatial pattern of functional diversity and identifying its main drivers in the temperate forests of northeastern China. We found that although environmental filtering by climate is important, stand structure explains most of the functional variation of the forest ecosystems in northeastern China. We also indicated that several abundant species determine most of the functional diversity,which supports the mass ratio hypothesis.Based on our study,the mixing of species with different functional groups is a more general approach to increase functional diversity. Particularly,a strong positive effect on functional diversity can be expected when coniferous and broad-leaved tree species are mixed. As a result, fostering structural complexity is a straightforward way to improve functional diversity without much ecological knowledge.
The main finding of our study was the evidence that stand structure based on forest types drive the variation of functional diversity.Ehbrecht et al. (2021) speculated that the climate-structure relationship they found in primary forests might be controlled by the relationship between climate and functional traits.Therefore,it is also worth paying attention to whether there is an inverse relationship between structural complexity and functional diversity in future studies.
Funding
This work was supported by the Program of National Natural Science Foundation of China (No. 31971650), and the Key Project of National Key Research and Development Plan (No. 2017YFC0504104), and Beijing Forestry University Outstanding Young Talent Cultivation Project(No. 2019JQ03001).
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Authors' contributions
JW designed the study; ZL analyzed data, and wrote the initial manuscript; and KG and CF provided comments and other technical support;JW revised the manuscript.All authors contributed critically to the manuscript and gave final approval for publication.
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Acknowledgements
We are grateful to colleagues who survey data together and Weixue Luo for assistance in data analysis.
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://do i.org/10.1016/j.fecs.2022.100048.

- Forest Ecosystems的其它文章
- Prediction of the global potential geographical distribution of Hylurgus ligniperda using a maximum entropy model
- The 2/3 scaling of twig nitrogen to phosphorus in woody plants
- Monitoring the abundance of saproxylic red-listed species in a managed beech forest by landsat temporal metrics
- Trade-offs among fine-root phosphorus-acquisition strategies of 15 tropical woody species
- Evaluating and quantifying the effect of various spruce budworm intervention strategies on forest carbon dynamics in Atlantic Canada
- Consistent response of nematode communities to management of coniferous plantations