
山茱萸CoHMGR基因的亚细胞定位和表达分析
2022-10-14 12:31王瑶瑶凡贞洁刘晓冉张佳琪胥华伟侯小改侯典云
中草药 2022年19期
王瑶瑶,凡贞洁,李 炯,刘晓冉,王 龙,张佳琪,胥华伟,侯小改*,侯典云*
山茱萸基因的亚细胞定位和表达分析
王瑶瑶1, 2,凡贞洁1, 2,李 炯3,刘晓冉1, 2,王 龙1, 2,张佳琪1, 2,胥华伟1, 2,侯小改1, 2*,侯典云1, 2*
1. 河南科技大学农学院,河南 洛阳 471023 2. 洛阳市道地药材繁育与创新利用工程技术研究中心,河南 洛阳 471023 3. 河南省农业农村厅中药材生产技术服务中心,河南 郑州 450000
研究山茱萸基因编码蛋白质的亚细胞定位以及不同浓度的外源激素对基因表达和活性成分含量的影响。农杆菌菌液侵染烟草叶片,激光共聚焦倒置显微镜观察CoHMGR蛋白的亚细胞定位。不同浓度的茉莉酸甲酯和乙烯利喷施山茱萸2年生幼苗,实时荧光定量PCR(quantitative real-time PCR,qRT-PCR)测定不同时期基因表达水平,高效液相色谱(HPLC)检测莫诺苷、马钱苷含量。激光共聚焦显微镜下观察到CoHMGR蛋白定位于内质网中。qPCR和HPLC结果显示200 μmol/L茉莉酸甲酯和250 mg/L乙烯利对目的基因表达水平和活性成分含量都有正向诱导效应。CoHMGR蛋白定位于内质网,外源喷施200 μmol/L茉莉酸甲酯和250 mg/L乙烯利能显著提高基因表达量和莫诺苷、马钱苷含量,为深入解析基因功能奠定基础。
山茱萸;;亚细胞定位;莫诺苷;马钱苷;表达分析
山茱萸Sieb. et Zucc.为多年生落叶乔木或灌木,其成熟去核干燥后的果肉入药,是多种中成药的主要成分,在河南、山西、甘肃、山东和浙江等省份都广有分布[1]。山茱萸含有环烯醚萜苷类、黄酮类、三萜类、多糖和鞣质等多种活性物质,其中,环烯醚萜类化合物是山茱萸的特征性物质[2]。山茱萸环烯醚萜化合物种类众多,具有代表性的是莫诺苷和马钱苷,越来越多山茱萸环烯醚萜类物质也在不断的被发掘[3]。现代医学研究表明,山茱萸具有多种药理功能,如从山茱萸中提取的总萜类物质可以有效降低血糖;山茱萸的环烯醚萜总苷既可以降低血糖和调节血脂,又能够起到抑制血栓形成的作用[4-6]。目前关于山茱萸的研究主要集中在活性成分的挖掘和药理作用的探究等方面,有关山茱萸萜类合成途径的研究十分有限,因此从分子生物学的角度解析山茱萸萜类生物合成途径关键基因的功能显得非常重要。
3-羟基-3-甲基-戊二酰辅酶A还原酶(3-hydroxy-3-methyl-glutaryl-CoA reductase,HMGR)催化萜类生物合成甲羟戊酸途径(mevalonic acid,MVA)中的第3步反应,HMGR为该途径中的第1个限速酶,是调控萜类次生代谢产物合成的关键酶[7]。近年来,越来越多来自不同植物的基因被克隆,并对其功能进行了初步验证[8]。徐晓腾等[9]克隆并分析了人参皂苷合成过程中的重要酶,从多个数据库进行了注释分析;郭思远等[10]克隆了东北雷公藤基因,并对该基因做出了生物信息学分析,预测基因编码的蛋白质位于内质网中。
本实验基于课题组前期获得的山茱萸转录组数据>c100572_g1[11],构建表达载体,根据基因编码蛋白在烟草叶片细胞中的荧光位置判断CoHMGR蛋白的亚细胞定位,同时探究了喷施不同浓度茉莉酸甲酯和乙烯利对山茱萸基因的表达模式及其活性成分含量的影响,为阐明山茱萸萜类物质分子合成途径提供参考。
1 材料与仪器
1.1 材料
山茱萸2年生幼苗种植于河南科技大学农学院试验田中,由河南科技大学戴攀峰副教授鉴定为山茱萸Sieb. et Zucc.,种植期间水肥管理一致,选取生长状态基本相同的植株作为外源激素喷施对象,喷施完成后摘取叶片为实验材料。选取本氏烟草为亚细胞定位的材料。
1.2 仪器
FV3000型激光扫描共聚焦显微镜(Olympus公司),5415 R型高速冷冻离心机(Eppendorf公司),紫外-可见光分光光度计来源,JY04S-3D型凝胶成像分析系统(君意电泳公司)等。
2 方法
2.1 山茱萸RNA提取和反转录
称取山茱萸叶片70~80 mg,装入标记好的灭菌1.5 mL离心管中,球磨仪研磨叶片成粉末,按照诺贝莱RNA提取试剂盒步骤提取山茱萸叶片总RNA,跑胶确认其完整度后,用微量分光光度计测定浓度,再使用艾德莱反转录试剂盒根据需要得到一部分用于PCR扩增的cDNA,一部分用于实时荧光定量PCR实验的cDNA,标记好保存于−20 ℃冰箱备用。
2.2 亚细胞定位表达载体构建
山茱萸基因开放阅读框全长1731 bp,共编码576个氨基酸。使用Primer Premier5.0软件设计引物,去掉开放阅读框的终止子,在终止子这端添加酶切位点,-F:5’-ATGGACGTTCGCAGGAGACAATCC-3’,- R-Ⅰ:5’-GCAGAGCACCCCTTGCT- TATATC-3’。以山茱萸总RNA反转录得到的cDNA为模板扩增出具有Ⅰ酶切位点的基因序列,反应条件:94 ℃、5 min;94 ℃、30 s,56 ℃、30 s,72 ℃、2 min,32个循环;72 ℃、5 min。将该序列连接到pMD18-T载体,转化至感受态的大肠杆菌DH10B中,选取单菌落摇菌,用上述引物进行菌液PCR扩增筛选阳性菌液送公司测序,序列比对正确后提取质粒备用。用于亚细胞定位实验的表达载体pCambia1300-GFP保存在大肠杆菌DH10B中,提取质粒备用。因为pMD18-T载体上也有Ⅰ酶切位点,因此采用Ⅰ单酶切的方法同时酶切连接到克隆载体上的序列和pCambia1300-GFP质粒,酶切过后会形成互补的黏性末端,在T4连接酶的作用下,形成新的重组载体pCambia1300- CoHMGR-GFP,按上述步骤转化到大肠杆菌DH10B中,筛选、测序比对正确后提取质粒进行后续实验。
构建好的重组质粒pCambia1300-CoHMGR- GFP转化到感受态农杆菌GV3101中待用;用软件在线预测CoHMGR蛋白位于内质网,选用ER-HDEL载体作为内质网标记(由南京农业大学张群教授惠赠)。将包含pCambia1300-CoHMGR- GFP质粒的农杆菌、空载pCambia1300-GFP和带有ER-HDEL标记的农杆菌分别扩大培养摇菌16 h以上,测定菌液的600在0.8~1.0内,就可以收集菌体,再用配制好的注射液重悬菌体,从叶片下表皮注射进烟草叶片内,暗培养两天后观察结果。
2.3 外源激素处理
分别配制100、200、400 μmol/L茉莉酸甲酯和250、500、750 mg/L乙烯利溶液。选取生长状态一致的山幼苗,分别喷施配制好的茉莉酸甲酯和乙烯利,喷施时间为17:30—19:30时,每个处理喷施量为500 mL,分别在喷施后0、1、3、5、7 d时取样,每株取若干叶片,将每个处理不同株上的叶片混合在一起,采集得到的叶片离体后立即用蒸馏水冲洗干净,分成3份装入材料袋,液氮处理后保存至−80 ℃冰箱。
2.4 山茱萸CoHMGR表达量检测
设计基因的特异性扩增引物用于实时荧光定量PCR实验:q--F:5’-TG- ACAAGAAGCCAGCAGCAGT-3’、q-- R:5’-GGCATTAAATCCACCAAGAGC-3’。以基因作内参,引物序列为:q-CoGAPDH-F:5’-TATCAAGGAGGAGTCAGAG- G-3’、q-CoGAPDH-R:5’-CCATTCGTTGTCAT- ACCAGG-3’。以“2.1”项方法获得激素处理后的叶片cDNA为模板,按照SYBR试剂盒说明书操作,每个反应3次重复,检测不同激素处理下基因随着时间变化的表达水平,根据相对表达量判断基因对茉莉酸甲酯和乙烯利的响应情况。
2.5 山茱萸叶片中莫诺苷、马钱苷含量测定[12]
称取激素处理后的样品0.5 g左右,每个样品3个重复,在装有研磨珠的10 mL离心管内研磨破碎山茱萸叶片成粉末,从研磨仪中取出离心管后迅速向管内加入80%甲醇,接着用超声波清洗机提取叶片中的化合物,提取条件为60 ℃、超声40 min,期间需要振荡2~3次,提取完成后,4000 r/min、4 ℃离心5 min,无菌针管吸取上清液过有机滤膜到棕色液相小瓶中,盖紧瓶盖置于4 ℃冰箱备用。
使用安捷伦高效液相色谱仪(Agilent 1260 Infinity)测量制备好的样品和标品,色谱柱为Hypersil GOLD-Agilent柱(250 mm×4.6 mm,5 μm)条件为:VWD检测器,流动相为甲醇-水(25∶75),每个样品的检测时间为40 min,检测波长240 nm,柱温箱30 ℃,进样体积10 μL,体积流量1 mL/min,按照上述条件测定样品及对照。方法学考察参照文献[12],RSD值均小于0.5%。
2.6 数据分析
利用Excel处理数据,整理得到激素处理后的CoHMGR表达量和莫诺苷、马钱苷含量,将数据输入GraphPad Prism 6作图,通过SPSS软件分析不同激素处理下每个时间点内基因表达量、代谢物含量的差异显著性,采用Pearson相关性分析探究基因表达量和莫诺苷、马钱苷含量之间的关系。
3 结果与分析
3.1 RNA提取和PCR扩增
按照“2.1”项提及方法获得的山茱萸叶片总RNA如图1所示,PCR扩增得到添加了Ⅰ酶切位点的基因序列(图2)。

M-Marker 1~3-总RNA
M-Marker 1、2-PCR扩增产物
3.2 CoHMGR蛋白的亚细胞定位
剪下注射完农杆菌菌液并且暗培养2 d后的烟草叶片,下表皮朝上放置于载玻片上,盖玻片轻轻盖住,排去多余气泡即可完成制片。将制备好的样品倒置放在激光共聚焦显微镜的载物台上,观察烟草叶片下表皮细胞中的荧光信号(图3),CoHMGR蛋白所发的绿色荧光和ER-HDEL标记的红色荧光重合,图片重叠后发出黄色荧光,表明CoHMGR蛋白位于内质网中,与软件预测结果一致。
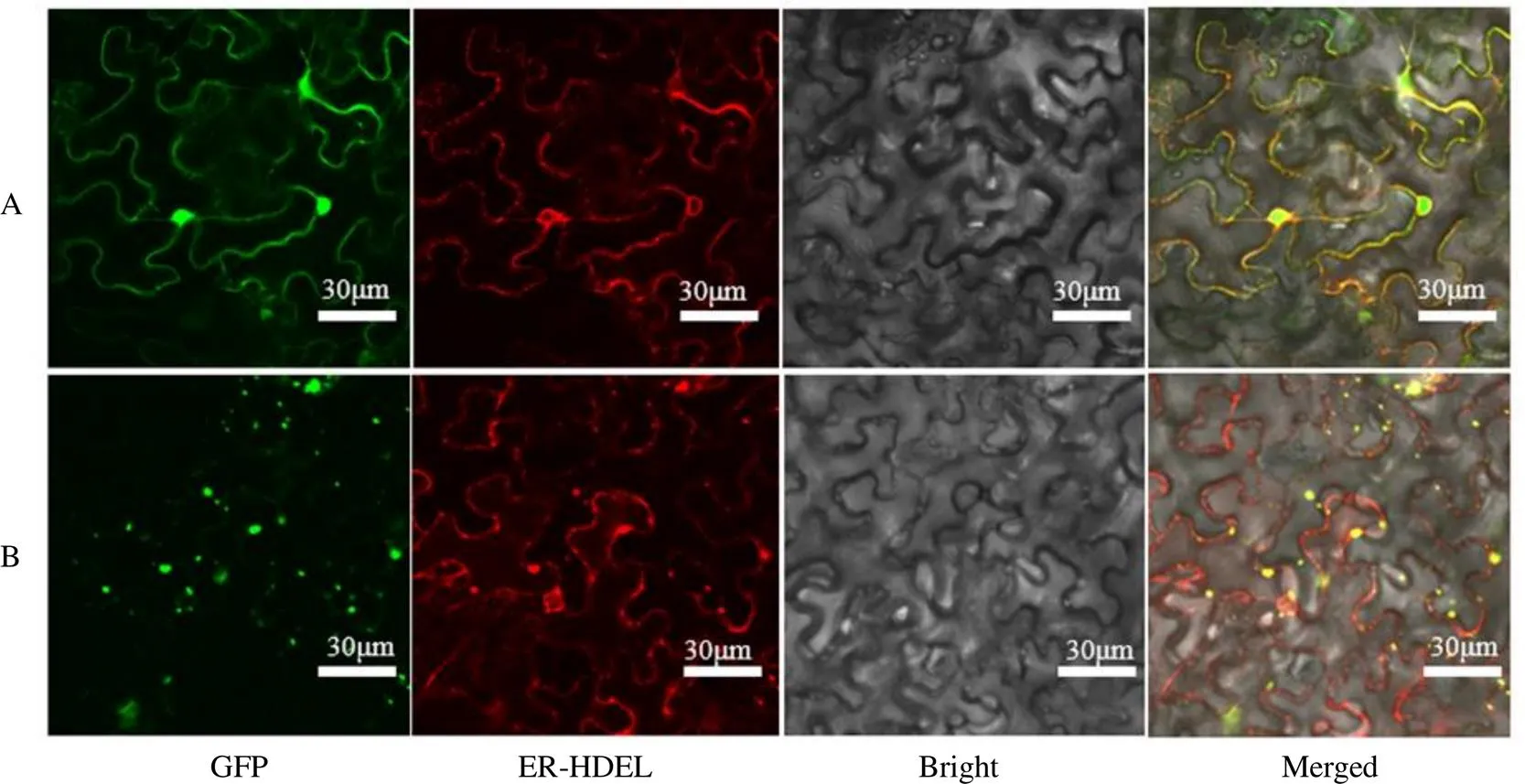
A-GFP载体 B-CoHMGR-GFP载体
3.3 茉莉酸甲酯、乙烯利对CoHMGR基因表达的影响
不同浓度茉莉酸甲酯和乙烯利处理后的基因表达水平受到了一定程度的影响,结果显示(图4),200 μmol/L的茉莉酸甲酯对基因的诱导效应强于100 μmol/L和400 μmol/L浓度,在处理后的第1天达到最高值比处理前提高了10倍左右,第3、5、7天基因表达量虽然有所下降,但是依然比处理前高出4倍;其余2个浓度的茉莉酸甲酯对基因没有明显的诱导作用。250 mg/L的乙烯利对基因的正向诱导作用最大,在处理后的第1天目的基因表达量提高了11倍,接下来几天其表达水平逐渐平稳,表达量基本是0 d的4倍;500 mg/L和750 mg/L的乙烯利喷施对基因的表达量没有明显持续的影响。以上结果证明200 μmol/L的茉莉酸甲酯和250 mg/L的乙烯利对基因都有正向且持续的诱导作用,显著提高了目的基因的表达水平。
3.4 山茱萸叶片中的活性成分分析
根据激素处理后山茱萸叶片中基因表达水平的检测结果,选取了200 μmol/L茉莉酸甲酯和250 mg/L乙烯利处理后的样品,通过高效液相色谱测定这2个处理后山茱萸叶片中的莫诺苷和马钱苷的含量。结果表明(图5),叶片中的莫诺苷含量在茉莉酸甲酯的处理后显著升高,茉莉酸甲酯处理后的第1天莫诺苷含量达到最大值,是处理前的2.28倍,接下来的几天内莫诺苷的含量虽然低于第1天,但是也显著高于处理前;乙烯利处理后叶片中的莫诺苷含量动态变化与茉莉酸甲酯处理下相似,先是在第1天就有显著提升,接着在第3天和第5天含量有所下降,但是在第7天莫诺苷含量达到了最大值,是0 d的1.99倍。激素处理对马钱苷含量的影响不同于莫诺苷,叶片中的马钱苷含量在处理后的7 d内呈现逐渐上升的趋势。茉莉酸甲酯和乙烯利处理后的马钱苷含量都在第7天达到了最大值,分别是处理前的3.21倍和3.17倍。综上所述,200 μmol/L茉莉酸甲酯和250 mg/L乙烯利对山茱萸叶片中莫诺苷和马钱苷的生成都有明显的促进作用。

CK-对照 M1、M2、M3-100、200、400 μmol·L−1茉莉酸甲酯处理 E1、E2、E3-250、500、750 mg·L−1乙烯利处理 不同小写字母表示处理间差异显著(P<0.05),下同

图5 激素处理后山茱萸叶片莫诺苷(A) 和马钱苷(B) 含量
3.5 CoHMGR表达量和莫诺苷、马钱苷含量相关性分析
对外源激素处理下的目的基因表达量和次生代谢产物莫诺苷、马钱苷含量进行皮尔逊(Pearson)关性分析,结果如表1所示,表达量和莫诺苷含量之间的相关系数为0.774,说明这两者之间呈强正相关性,即表达量越高,莫诺苷含量也就越高;但是表达量和马钱苷含量之间的相关系数仅为0.074,表明这两者之间相关性极弱;莫诺苷含量和马钱苷含量之间的相关系数为0.335,说明这两者之间存在较弱相关性。
表1 激素处理下CoHMGR相对表达量与莫诺苷、马钱苷含量的相关性
Table 1 Correlation between relative expression of CoHMGR and contents of morronin and loganin under hormone treatment
项目Pearson相关系数 CoHMGR莫诺苷含量马钱苷含量 CoHMGR1.000 莫诺苷含量 0.774**1.000 马钱苷含量 0.0740.3351.000
**< 0.01
4 讨论
关于HMGR蛋白的亚细胞定位研究的报道较少,拟南芥HMGR蛋白的亚细胞定位结果显示,基因编码的蛋白质一开始在内质网中合成,然后通过不同的方式运输至细胞内的不同囊泡状结构,再发挥具体作用[13]。王淋等[14]克隆了杜仲基因,通过瞬时转化烟草下表皮细胞,观察重组蛋白在细胞内的具体荧光位置,判断EuHMGR蛋白质位于内质网中。本课题组构建了pCambia1300-CoHMGR-GFP重组载体,农杆菌介导转化进烟草细胞内,激光共聚焦显微镜观察到基因编码蛋白质所发的绿色荧光和内质网标记蛋白ER-HDEL所发的红色荧光重合,因此初步判断CoHMGR蛋白位于内质网。
基因受多种外界因素的调控,研究发现温度、盐胁迫处理均可以影响人参基因的表达量[15-16];生物诱导物如大肠杆菌能够提高火索麻悬浮培养物中基因和基因的表达水平,从而上调薯蓣皂苷元的产量[17];对铁皮石斛幼苗施加内生真菌菌液后,铁皮石斛基因表达量明显上调[18]。
植物生长调节剂通常用于促进植物的生长发育,但是最近的研究发现部分植物生长调节剂也可以诱导植物体内萜类物质途径关键基因的表达和萜类物质的生成,这可能是提高药用植物特征活性成分的有效途径[19]。茉莉酸甲酯和乙烯利都是激活植物防御系统的的重要激素,外源施加这2种激素能够提高药用植物的有效成分产量。
银杏是我国特产的珍贵树种,据报道,不同的外源激素处理银杏幼苗,会诱导甲羟戊酸途径内基因表达和银杏内酯生成,结果显示茉莉酸甲酯、乙烯利都显著提升了银杏、、的表达量,乙烯利对、基因也有正向诱导作用,这2种激素都显著提升了处理后叶片中银杏内酯的含量[20-22]。目前,关于茉莉酸甲酯诱导药用植物次生代谢产物产生的研究比较广泛,已经报道的有茉莉酸甲酯诱导灵芝三萜物质的合成[23],提高罗汉果皂苷及其中间产物含量[24]等,乙烯利在促进药用植物活性成分生成方面的研究较少。
山茱萸生长周期长,生长年限达到7~10年才能结果,盛产期则需要等20年之久,这给在山茱萸本体中研究关键基因造成了不小的困难,因此本课题组选用了山茱萸2年生幼苗作为激素喷施的实验对象[25]。本研究中,设计了不同浓度的茉莉酸甲酯、乙烯利分别处理山茱萸幼苗叶片,综合实时荧光定量PCR结果和高效液相色谱结果发现,200 μmol/L茉莉酸甲酯和250 mg/L乙烯利处理都正向调控了基因表达量和山茱萸叶片中莫诺苷、马钱苷含量,这也在一定程度上说明基因在山茱萸萜类物质合成中起重要作用,对目的基因表达量和莫诺苷、马钱苷含量的相关性分析结果表明:表达量与莫诺苷含量之间有较强的联系,和马钱苷含量的相关性则不大,为后续研究基因功能、解析山茱萸萜类物质合成途径以及使用外源激素促进山茱萸活性成分生成提供参考。
利益冲突 所有作者均声明不存在利益冲突
[1] 叶小风, 范祥祯, 应智琦, 等. 山茱萸的资源分布及开发应用现状 [J]. 农技服务, 2021, 38(3): 84-87.
[2] Dong Y, Feng Z L, Chen H B,.:A review of chemical constituents and pharmacological activities [J]., 2018, 13: 34.
[3] He J, Xu Y, Xu J K,. Cornusglucosides A and B, two new iridoid glucosides from the fruit of[J]., 2019, 16(11): e1900421.
[4] 相聪坤. 山茱萸资源及活性成分研究进展 [J]. 河北医药, 2016, 38(12): 1886-1889.
[5] Huang J, Zhang Y W, Dong L,. Ethnopharmacology, phytochemistry, and pharmacology ofSieb. et Zucc [J]., 2018, 213: 280-301.
[6] 刘薇, 朱晶晶, 徐志猛, 等. 山茱萸总萜对KKay糖尿病小鼠的治疗作用研究[J]. 药物评价研究, 2016, 39(6): 947-952.
[7] 郝爱平, 国会艳, 薛巨坤. 31种药用植物HMGR蛋白的生物信息学分析 [J]. 湖北农业科学, 2017, 56(14): 2761-2767.
[8] 赵志新, 李秀秀. 药用植物中HMGR的生物信息学分析 [J]. 陕西农业科学, 2018, 64(4): 43-46.
[9] 徐晓腾, 李翔宇, 林彦萍, 等. 人参皂苷合成关键元件HMGR基因生物信息学分析 [J]. 生物技术, 2016, 26(2): 163-168.
[10] 郭思远, 闫琦, 李佳. 东北雷公藤DXR HMGR基因克隆及生物信息学分析 [J]. 中国现代中药, 2019, 21(11): 1482-1488.
[11] Hou D Y, Shi L C, Yang M M,. De novo transcriptomic analysis of leaf and fruit tissue ofusing Illumina platform [J]., 2018, 13(2): e0192610.
[12] 王瑶瑶. 山茱萸CoHMGS、CoHMGR基因的克隆及表达分析 [D]. 洛阳: 河南科技大学, 2021.
[13] Leivar P, González V M, Castel S,. Subcellular localization of3-hydroxy-3-methylglutaryl- coenzyme A reductase [J]., 2005, 137(1): 57-69.
[14] 王淋, 杜红岩, 乌云塔娜. 杜仲MVA和MEP途径相关基因的亚细胞定位与表达分析 [J]. 植物研究, 2017, 37(1): 52-62.
[15] 孙亚静. 温度对人参转录因子及皂苷合成酶基因表达能力的影响 [D]. 长春: 吉林大学, 2021.
[16] 崔悦. 盐胁迫对人参转录因子及皂苷合成酶基因表达能力的影响 [D]. 长春: 吉林大学, 2021.
[17] Shaikh S, Shriram V, Khare T,. Biotic elicitors enhance diosgenin production inL. suspension cultures via up-regulation ofandgenes [J]., 2020, 26(3): 593-604.
[18] 王进, 李俊峰, 张婷婷, 等. 内生真菌对铁皮石斛多糖和生物碱合成关键酶基因表达的影响 [J]. 中草药, 2019, 50(23): 5838-5846.
[19] 马琳, 郜玉钢, 臧埔, 等. 植物生长调节剂对药用植物次生代谢物积累的影响 [J]. 中南药学, 2016, 14(8): 834-837.
[20] Meng X X, Song Q L, Ye J B,. Characterization, function, and transcriptional profiling analysis of 3-hydroxy-3- methylglutaryl-CoA synthase gene () towards stresses and exogenous hormone treatments in[J]., 2017, 22(10): E1706.
[21] Rao S, Meng X X, Liao Y L,. Characterization and functional analysis of two novel 3-hydroxy-3- methylglutaryl- coenzyme A reductase genes (and) from[J]., 2019, 9(1): 14109.
[22] Chen Q W, Yan J P, Meng X X,. Molecular cloning, characterization, and functional analysis of acetyl-CoA-acetyltransferase and mevalonate kinase genes involved in terpene trilactone biosynthesis from[J]., 2017, 22(1): E74.
[23] 任昂, 缪志刚, 赵明文. 茉莉酸甲酯诱导灵芝三萜生物合成的机制研究[A] // 第十届全国全国食用菌学术研讨会论文汇编[C]. 北京: 中国菌物学会, 2014: 68.
[24] 张凯伦. 茉莉酸甲酯对罗汉果皂苷生物合成途径的影响及罗汉果中bHLH转录因子的初筛 [D]. 北京: 北京协和医学院, 2016.
[25] 史淑红. 山西山茱萸形态解剖学与主要有用成分含量变化规律研究 [D]. 太原: 山西大学, 2017.
Subcellular localization and expression analysis ofgene from
WANG Yao-yao1, 2, FAN Zhen-jie1, 2, LI Jiong3, LIU Xiao-ran1, 2, WANG Long1, 2, ZHANG Jia-qi1, 2, XU Hua-wei1, 2, HOU Xiao-gai1, 2, HOU Dian-yun1, 2
1. College of Agricultural, Henan University of Science and Technology, Luoyang 471023, China 2. The Luoyang Engineering Research Center of Breeding and Utilization of Dao-di Herbs, Luoyang 471023, China 3. Chinese Medicinal Materials Production Technology Service Center of Department of Agriculture of Henan Province, Zhengzhou 450000, China
To study the subcellular localization of the protein encoded by thegene of, and the correlation of spraying different concentrations of exogenous hormones on the expression ofgene and the content of active ingredients.Agrobacterium infects tobacco leaves, and the subcellular localization of CoHMGR protein was observed under a confocal inverted laser microscope. Using different concentrations of methyl jasmonate and ethephon to spray two-year-old seedlings of, quantitative real-time PCR (qPCR) was used to determine the expression levels ofgene at different periods after treatment, high performance liquid chromatography (HPLC) was used to detect the content of morroniside and loganin.The CoHMGR protein localization in the endoplasmic reticulum was observed under the laser confocal microscope. The results of qPCR and HPLC showed that 200 μmol/L methyl jasmonate and 250 mg/L ethephon had a positive induction effect on the target gene expression level and active ingredient content.The CoHMGR protein is located in the endoplasmic reticulum. The exogenous spraying of 200 μmol/L methyl jasmonate and 250 mg/L ethephon can significantly increase the expression ofand the content of monoside and loganin, laying a foundation for in-depth analysis of the function ofgene.
Sieb. et Zucc.;; subcellular localization; morroniside; loganin; expression analysis
R286.12
A
0253 - 2670(2022)19 - 6174 - 06
10.7501/j.issn.0253-2670.2022.19.023
2022-03-06
国家自然科学基金资助项目(U1404829);中央本级重大增减支项目(2060302);河南省自然科学基金项目(202300410151);河南省科技攻关项目(202102110156);河南省中药材产业技术体系建设专项资金资助;河南省中药材产业科技特派员服务团资助
王瑶瑶,硕士研究生,主要从事中药资源评价与利用方面的研究。E-mail: 18438616133@163.com
侯典云,教授,博士生导师。Tel: (0379)64282340 E-mail: dianyun518@163.com
[责任编辑 时圣明]
猜你喜欢
河南农业·教育版(2022年3期)2022-04-23
保健与生活(2021年6期)2021-03-16
家庭医学(2020年10期)2020-11-20
家庭医学(2020年8期)2020-09-02
意林·全彩Color(2019年11期)2019-12-30
意林彩版(2019年11期)2019-11-22
下一代(2019年3期)2019-09-10
阅读(低年级)(2019年5期)2019-07-18
当代工人·精品C(2019年1期)2019-04-29
戏剧之家(2016年1期)2016-02-25
