
饲粮添加硫辛酸对肉鸡生长性能、屠宰性能、脂肪代谢和肠道菌群的影响
2022-10-12 03:23李金燕聂存喜南珊珊牛俊丽李佳橙
动物营养学报 2022年9期
李金燕 聂存喜 南珊珊 牛俊丽 李佳橙
(石河子大学动物科技学院,石河子832000)
硫辛酸是一种类维生素物质,兼具脂溶性和水溶性特性,可清除引起慢性疾病和衰老的自由基和活性氧,增强生物体肠道内消化酶活性,调节肠道菌群,进而维持肠道健康状态,起到促进动物健康生长的作用[1]。研究表明,饲粮中添加硫辛酸后可在一定程度上增强肉鸡抗氧化能力,并可提高其免疫功能[2]。El-Senousey等[3]研究发现,与维生素C和维生素E相比,肉鸡饲粮中添加500 mg/kg硫辛酸具有更好的抵抗地塞米松诱导的氧化应激的效果。杨国宇等[4]在肉鸡饲粮中添加0、20、60和100 mg/kg硫辛酸的研究表明,60和100 mg/kg的添加量可以降低肉鸡肝脏中脂肪及血液中甘油三酯和总胆固醇含量。Hamano等[5]研究了饲喂50 mg/kg硫辛酸对肉鸡翅静脉注射异丙肾上腺素的代谢响应,结果表明硫辛酸可降低其血浆中总胆固醇的含量,在维持血浆葡萄糖含量正常的情况下,促进肉鸡脂肪酸动员。有研究报道,硫辛酸能够有效清除生物体内的活性氧自由基,让生物体内肝脏抗氧化酶活性获得提升,同时还能够显著降低肝脏中镉的含量,避免肝脏脂肪出现大量累积,让肝脏拥有更强的解毒能力[6]。在肉鸡饲粮中添加硫辛酸的最新研究表明,400~1 200 mg/kg硫辛酸可以改善肉品质[7],400和800 mg/kg均可改善肉鸡生长性能和胴体性状且二者无显著差异[8]。
不过,硫辛酸对肉鸡生长性能、屠宰性能、脂肪代谢和肠道菌群的系统研究未见报道,尤其是从肠道菌群的角度解析硫辛酸对肉鸡生产的调控作用还属于空白。基于此,本文以白羽肉鸡为试验对象,研究饲粮添加硫辛酸对肉鸡的应用效果,以期从肠道微生物角度探究硫辛酸对肉鸡生产的代谢响应,为硫辛酸的合理利用和以肠道健康为靶点的肉鸡健康养殖提供理论和生产指导。
1 材料与方法
1.1 试验材料
试验所用硫辛酸购自江苏某生物技术有限公司,纯度为99%;所用玉米、豆粕等饲料原料均购自当地农贸市场。
1.2 试验设计和饲养管理
试验选取14日龄体重相近、健康的白羽肉鸡128只,随机分为2个组,即对照组和试验组,每组4个重复,每个重复16只鸡。对照组饲喂基础饲粮,试验组饲喂在基础饲粮的基础上添加400 mg/kg硫辛酸的饲粮。肉鸡基础饲粮参考NRC(1994)配制,其组成及营养水平见表1。试验期28 d,期间肉鸡自由采食和饮水,并按照标准免疫流程完成疫苗接种工作。

1 Items Content Nutrient levels2) ME/(MJ/kg)12.85 CP19.43 Ca1.06 TP0.65 Lys1.02 Met0.38 Thr0.70
1.3 测定指标及方法
1.3.1 生长性能
在试验开始和试验结束时,以重复为单位,在肉鸡空腹的状态下称重,每日记录肉鸡采食量和健康状况。分别计算15~28日龄和29~42日龄肉鸡的平均日增重(ADG)、平均日采食量(ADFI)和料重比(F/G)。
1.3.2 屠宰性能
分别于肉鸡28和42日龄,从每个重复中随机抽取体重相近的2只鸡,即每组8只鸡,2组共计16只鸡进行屠宰性能测定。屠宰性能指标的测定根据家禽的生产性能评价统计方法[9]进行,包括肉鸡的屠宰率、全净膛率、半净膛率、胸肌率、腿肌率、腹脂率以及肝脏重等相关指标。皮下脂肪厚度测定是将尾椎结合处横向切开,再从背正中线纵向切一刀,用游标卡尺测量交叉处的脂肪厚度,每只鸡重复测量3次,取平均值,单位用mm表示。
1.3.3 脂肪代谢相关指标
分别于肉鸡28和42日龄,从每个重复中随机抽取体重相近的2只鸡,即每组8只鸡,2组共计16只鸡进行脂肪代谢相关指标测定。颈静脉采血5 mL,在室温中放置30 min后,3 000 r/min离心15 min制备血清,置于-20 ℃保存。屠宰放血后采集胸肌、腿肌样品各5 g,剔去肉眼可见的表面脂肪与筋膜,迅速分装样品并编号置于自封塑料袋中,将胸肌、腿肌剪碎,置于培养皿中,65 ℃烘至恒重,粉碎,即制备成胸肌、腿肌风干样。肉鸡血清、胸肌和腿肌中总胆固醇和甘油三酯的含量采用南京建成生物工程研究所提供的试剂盒进行测定,测定方法参照试剂盒说明书进行。
1.3.4 肠道菌群
分别于肉鸡28和42日龄,进行屠宰性能测定时收集肉鸡盲肠内容物于冻存管中,液氮冷冻后,置于-80 ℃保存待测。提取盲肠内容物微生物总DNA,以总DNA为模板,采用通用引物(338F,5′-ACTCCTACGGGAGGCAGCA-3′;806R,5′-GGACTACHVGGGTWTCTAAT-3′)对16S rRNA V3~V4区进行PCR扩增,然后在HiSeq 2500高通量测序平台进行16S rDNA高通量测序分析,检测工作委托上海中科新生命生物科技有限公司完成。
1.4 数据统计分析
使用Excel 2019对肉鸡生长性能、屠宰性能以及脂肪代谢相关指标等数据进行整理,采用SPSS 20.0软件对数据进行独立样本t检验,结果采用“平均值±标准差”的方式表示,以P<0.05为差异显著性水平。
肠道菌群测序原始数据下机后,首先采用QIIME 1.9.1软件进行数据质控,通过序列拼接、过滤和去嵌合体后得到优化序列,利用USEARCH 7.0软件将优质序列进行操作分类单元(OTU)聚类分析,序列相似性阈值为97%;参考SILVA 128数据库利用RDP Classifier 2.11软件贝叶斯算法进行OTU序列分类学分析,并按照界、门、纲、目、科、属、种的分类水平统计各样本的群落组成;利用R语言完成粪便菌群门主坐标分析(PCoA)的数据分析工作,之后使用STAMP 2.1.3软件实现数据的可视化展示;利用Mothur 1.30.2数据处理软件计算α多样性指数,其中主要包含Shannon指数、ACE指数、Simpson指数和Chao1指数。
2 结果与分析
2.1 饲粮添加硫辛酸对肉鸡生长性能的影响
由表2可知,在整个试验期不同阶段,与对照组相比,饲粮添加硫辛酸对肉鸡ADG、ADFI和F/G均未产生显著影响(P>0.05)。但与对照组相比,试验组肉鸡15~28日龄的ADFI降低了6.61%,F/G降低了3.78%;29~42日龄的ADFI提高了1.20%,F/G提高了4.23%。

表2 饲粮添加硫辛酸对肉鸡生长性能的影响
2.2 饲粮添加硫辛酸对肉鸡屠宰性能的影响
由表3可知,28日龄,试验组肉鸡屠宰率较对照组显著提高了0.81%(P<0.05);42日龄,试验组全净膛率较对照组显著降低了0.83%(P<0.05),胸肌率较对照组显著提高了9.34%(P<0.05);28和42日龄,与对照组相比,饲粮添加硫辛酸对肉鸡半净膛率、腿肌率、腹脂率、皮下脂肪厚度和肝脏重均无显著影响(P>0.05)。

表3 饲粮添加硫辛酸对肉鸡屠宰性能的影响
2.3 饲粮添加硫辛酸对肉鸡脂肪代谢相关指标的影响
由表4可知,28日龄,试验组肉鸡血清总胆固醇含量较对照组显著提高了19.78%(P<0.05);试验组和对照组腿肌、胸肌总胆固醇含量之间均无显著性差异(P>0.05)。42日龄,试验组腿肌总胆固醇含量较对照组显著降低了47.62%(P<0.05),而胸肌总胆固醇含量较对照组显著提高了33.33%(P<0.05);与对照组相比,试验组血清总胆固醇含量降低了5.25%,但差异不显著(P>0.05)。
28日龄,与对照组相比,试验组肉鸡血清、腿肌和胸肌甘油三酯含量均无显著性差异(P>0.05)。42日龄,试验组腿肌甘油三酯含量较对照组显著降低了14.94%(P<0.05),而胸肌甘油三酯含量较对照组显著提高了24.05%(P<0.05);与对照组相比,试验组血清甘油三酯含量降低了3.57%,但差异不显著(P>0.05)。

表4 饲粮添加硫辛酸对肉鸡脂肪代谢相关指标的影响
2.4 饲粮添加硫辛酸对肉鸡肠道菌群的影响
2.4.1 肉鸡肠道菌群OTU分析
通过维恩(Venn)图可以清晰地展示出组间样本存在的独有以及特有的OTU数,这样能够客观地评价生物体内微生物群落相似度。由图1-A可知,28日龄,肉鸡盲肠共鉴定出1 529个OTU,其中对照组和试验组共有OTU 812个,占53.11%;由图1-B可知,42日龄,肉鸡盲肠共鉴定出1 726个OTU,其中对照组和试验组共有OTU 860个,占49.83%。
2.4.2 肉鸡肠道菌群α多样性分析
微生物群落的丰富度和多样性可以通过α多样性分析来表示。由表5可知,各组的Coverage指数均在1左右,说明测序数据约覆盖100%的微生物,覆盖度较好。28日龄,与对照组相比,试验组Shannon指数和Simpson指数分别显著提高了27.15%和20.55%(P<0.05);42日龄,与对照组相比,试验组α多样性指数均无显著性差异(P>0.05)。
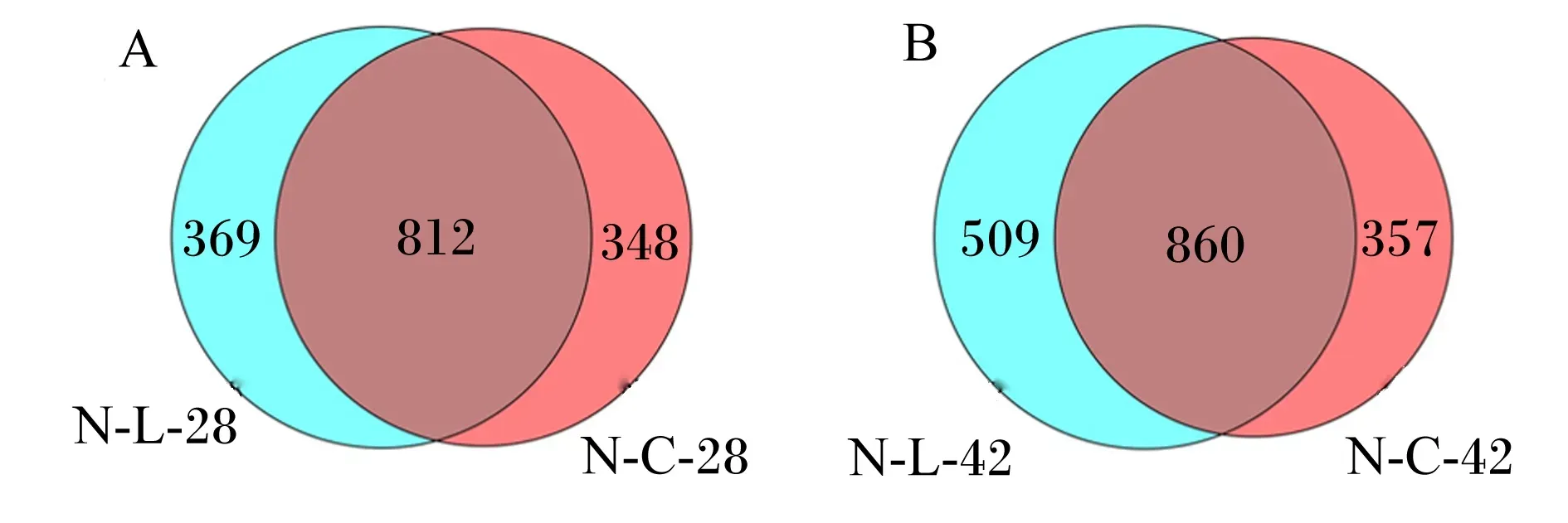
N-C-28和N-C-42分别表示对照组28和42日龄,N-L-28和N-L-42分别表示试验组28和42日龄。下图同。
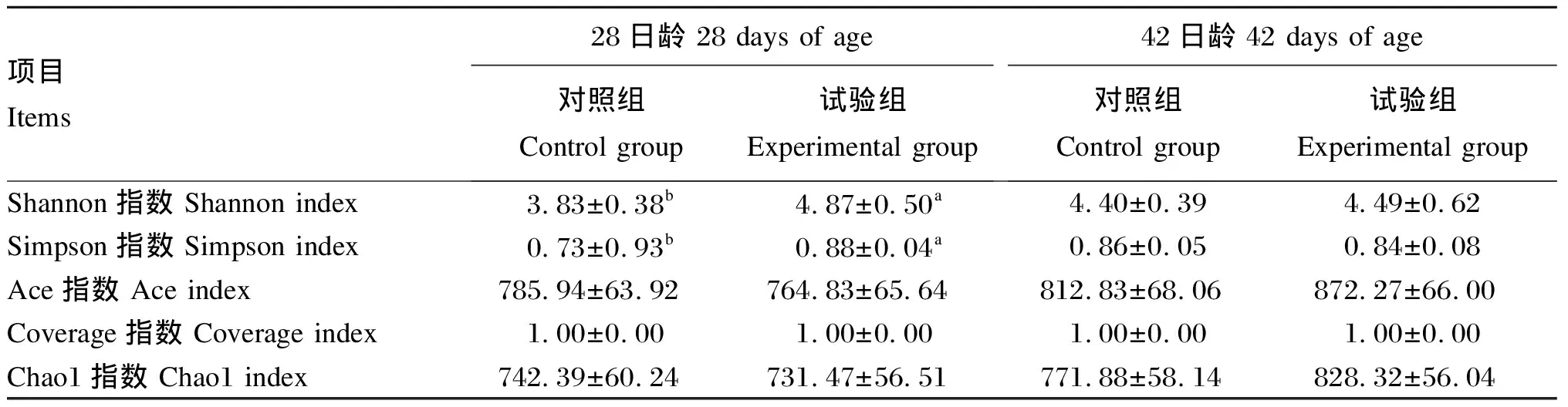
表5 饲粮添加硫辛酸对肉鸡肠道菌群α多样性的影响
2.4.3 肉鸡肠道菌群差异分析
为了便于观察各组之间的肠道菌群差异,应用非度量多维尺度(NMDS)图对各组粪便微生物群落进行测试。图2中每个点代表1个样本,利用样品点之间的距离来评判个体以及群体两者存在的不同和差异性,其距离越小,则表明微生物群落具有更强的相似性。由图2可知,28和42日龄,对照组和试验组各样品之间均可区分开来,表明组间微生物群落组成差异明显。
2.4.4 肉鸡肠道菌群在门水平上的组成
由图3可知,28日龄,对照组和试验组在门水平上的优势菌群为拟杆菌门(Bacteroidetes)、厚壁菌门(Firmicutes)、变形菌门(Proteobacteria)和疣微菌门(Verrucomicrobia);42日龄,对照组和试验组在门水平上的优势菌群为拟杆菌门、厚壁菌门、互养菌门(Synergistetes)和变形菌门。
2.4.5 肉鸡肠道菌群在属水平上的组成
由图4可知,28日龄,对照组和试验组在属水平上的优势菌群均为理研菌科RC9肠道群(Rikenellaceae_RC9_gut_group)、拟杆菌属(Bacteroides)、瘤胃球菌科UCG_014(Ruminococcaceae_UCG_014)和考拉杆菌属(Phascolarctobacterium);42日龄,对照组和试验组在属水平上的优势菌群均为拟杆菌属、理研菌科RC9肠道群、考拉杆菌属和瘤胃球菌科UCG_014。

NMDS:非度量多维尺度 non-metric multidimensional scaling。

图3 肉鸡肠道菌群在门水平上的组成
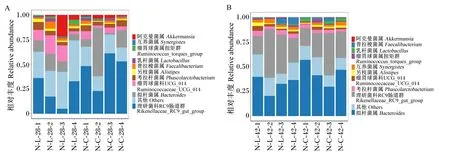
图4 肉鸡肠道菌群在属水平上的组成
2.4.6 肉鸡肠道菌群在属水平上的差异
由图5可知,28日龄,对照组和试验组在属水平上的差异菌群为不可培养菌(uncultured)、Shuttleworthia、肠球菌属(Enterococcus)、理研菌属(Rikenella)、萨特氏菌属(Sutterella)、Tyzzerella和Harryflintia;其中,与对照组相比,试验组中不可培养菌、Shuttleworthia、肠球菌属和理研菌属的相对丰度显著升高(P<0.05),萨特氏菌属、Tyzzerella、Harryflintia的相对丰度显著降低(P<0.05)。42日龄,对照组和试验组在属水平上的差异菌群为脱硫弧菌属(Desulfovibrio)、丁酸球菌属(Butyricicoccus)和Dielma;其中,与对照组相比,试验组中脱硫弧菌属和Dielma的相对丰度显著升高(P<0.05),丁酸球菌属的相对丰度显著降低(P<0.05)。

图5 肉鸡肠道菌群在属水平上的差异
2.4.7 肉鸡脂肪代谢相关指标与肠道菌群的相关性分析
由图6可知,肉鸡胸肌甘油三酯含量与萨特氏菌属、脱硫弧菌属和Dielma呈极显著正相关(P<0.01);胸肌总胆固醇含量与不可培养菌呈极显著正相关(P<0.01),而与丁酸球菌属呈显著负相关(P<0.05)。肉鸡腿肌甘油三酯含量与Shuttleworthia呈显著正相关(P<0.05),与理研菌属和Harryflintia呈显著负相关(P<0.05)。
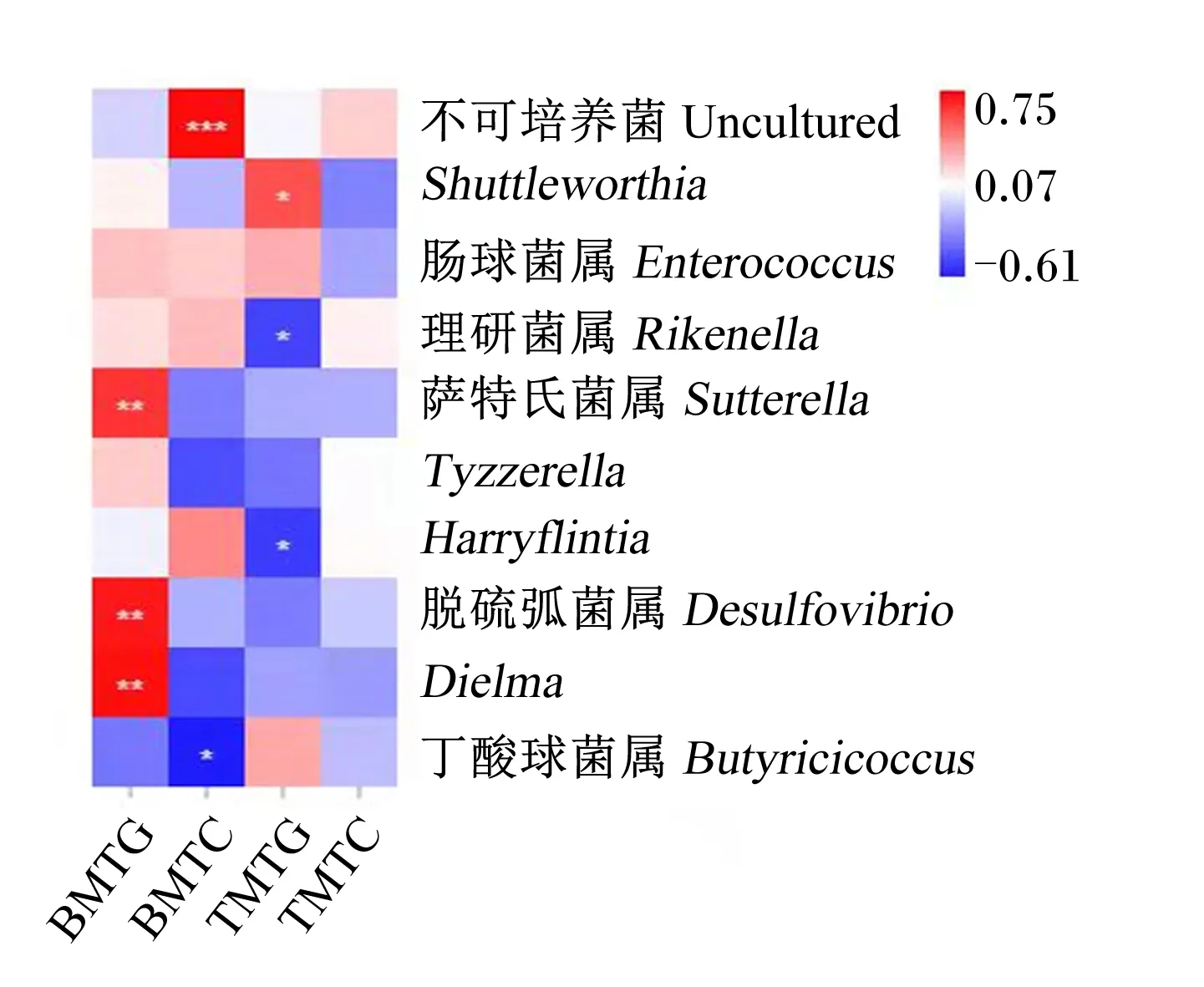
BMTG和BMTC分别表示肉鸡胸肌甘油三酯和总胆固醇含量;TMTG和TMTC分别表示肉鸡腿肌甘油三酯和总胆固醇含量。*表示P<0.05,**表示P<0.01,***表示P<0.001。
3 讨 论
3.1 饲粮添加硫辛酸对肉鸡生长性能的影响
生长性能是反映肉鸡生长最直接的指标,提高生长性能是增加经济效益的关键。硫辛酸作为一种功能性的饲料添加剂,可有效地减缓氧化应激给肉鸡生长性能带来的不利影响[3]。李杏[2]在饲粮中添加300 mg/kg硫辛酸后,未引起21~52日龄黄羽肉鸡生长性能的改变。El-Senousey等[7]在22~42日龄爱拔益加(AA)肉鸡饲粮中添加硫辛酸表明,400和800 mg/kg的添加量显著降低了肉鸡ADG和ADFI,但不影响饲料转化效率,1 200 mg/kg的添加量与800 mg/kg的添加量也无显著差异。然而,杨潇等[8]研究发现,1~42日龄肉鸡饲粮中添加400和800 mg/kg 硫辛酸时,肉鸡的ADFI和F/G均显著降低且二者之间无显著差异,但200 mg/kg的添加量未产生显著影响。郭志有等[10]研究发现,硫辛酸添加量达到1 000 mg/kg时,肉鸡的ADFI和体重都显著下降。由此可知,饲粮中添加适量硫辛酸能够促进动物生长,而过低剂量和过高剂量硫辛酸对动物的生长无促进作用。本试验结果表明,在肉鸡饲粮中添加400 mg/kg的硫辛酸对肉鸡生长性能无显著影响,这说明硫辛酸对肉鸡生长性能的影响与使用剂量和使用时间密切相关。
3.2 饲粮添加硫辛酸对肉鸡屠宰性能的影响
屠宰性能是衡量家禽产肉性能的重要指标。有研究表明,硫辛酸添加剂量在200~800 mg/kg时,对肉鸡屠宰率、胸肌率和腿肌率等屠宰性能的影响不显著[8]。也有研究表明,1~42日龄肉鸡饲粮中添加250~1 000 mg/kg硫辛酸并没有影响其屠宰率、全净膛率、胸肌率和腿肌率[10]。而本研究结果表明,饲粮添加400 mg/kg的硫辛酸可显著提高肉鸡28日龄屠宰率、42日龄胸肌率,显著降低42日龄全净膛率。这说明,硫辛酸对屠宰性能的影响在肉鸡生长后期效果更明显,且作用效果与肉鸡添加日龄和添加时间有关。本试验的结果显示,硫辛酸的添加不影响肉鸡腿肌率和腹脂率,这与文献报道结果[7]一致。
3.3 饲粮添加硫辛酸对肉鸡脂肪代谢相关指标的影响
体内脂肪代谢情况可以通过血清和组织总胆固醇和甘油三酯含量反映。本试验结果显示,与对照组相比,肉鸡42日龄腿肌中总胆固醇和甘油三酯含量分别显著降低了47.62%和14.94%,而胸肌中总胆固醇和甘油三酯含量分别显著提高了33.33%和24.05%。对于胸肌和腿肌脂肪代谢的差异变化说明硫辛酸的调控作用可能存在组织特异性,硫辛酸可促进胸肌脂质沉积而抑制腿肌脂质沉积。有研究发现,硫辛酸能够对脂肪代谢产生影响的主要原因是对肠道内糖的转运过程造成影响[11],也可能与肠道菌群的变化存在密切关系,为此本研究也进行了肠道菌群相关的研究。生物体内的脂质代谢的平衡与增强氧化供能有着紧密的关系。脂质沉积主要和甘油三酯储存与合成以及脂质氧化有关[12]。研究表明,饲粮添加100 mg/kg硫辛酸能够显著降低大鼠血脂和肝脏总胆固醇和游离脂肪酸含量,调节血脂紊乱,改善脂质沉积[13]。Koh等[14]研究发现,硫辛酸可以抑制脂肪细胞的分化,并在肥胖受试者中诱导体重减轻。
3.4 饲粮添加硫辛酸对肉鸡肠道菌群的影响
生物体内的肠道微生态系统对生物的消化吸收能力有着显著影响,确保肠道菌群处于平衡状态能够明显增强生物体的消化和吸收能力,有利于保持动物机体的健康[15]。本试验中,28日龄,与对照组相比,试验组Shannon指数和Simpson指数分别显著提高了27.15%和20.55%,表明该阶段的菌群物种多样性显著增加。42日龄,试验组各α多样性指数较对照组均无显著性差异,但试验组的Chao1指数、Ace指数和Shannon指数均高于对照组,这表明硫辛酸可增加肉鸡肠道菌群多样性。
鸡肠道中的菌群主要以厚壁菌门为主,其次为拟杆菌门、变形菌门和放线菌门(Actinobacteria)[16],肠道菌群改变可导致肠道的消化吸收功能发生改变[17]。当肠道中的厚壁菌门相对丰度提高时,能够帮助机体获取食物里的能量和热量[18],微生物代谢产生的短链脂肪酸也能确保生物体的肠道处于健康状态[19]。拟杆菌门是第2大主要门类,拟杆菌菌群发酵的终产物丙酸盐和乙酸盐,为宿主和肠道微生物所需[20];并影响胃肠道的正常发育和蛋白质、碳水化合物代谢[21]。研究表明,厚壁菌门和拟杆菌门与脂肪沉积存在一定的相关性,这2类微生物都能促进机体脂肪的沉积,且厚壁菌门比例升高时更有利于脂肪沉积[22]。在本试验中,肉鸡盲肠中拟杆菌属、理研菌科RC9肠道群、考拉杆菌属和瘤胃球菌科UCG_014为优势菌群;且与对照组相比,试验组理研菌属的相对丰度显著升高,试验组萨特氏菌属、Tyzzerella、Harryflintia等细菌的相对丰度显著降低。这表明,硫辛酸可通过影响肠道菌群来调控肉鸡脂肪沉积。
本研究对肠道差异性菌属与脂肪代谢相关指标进行相关性分析结果显示,在属水平,与胸肌或腿肌甘油三酯和总胆固醇含量呈正相关的菌群有Shuttleworthia、萨特氏菌属、脱硫弧菌属和Dielma,呈负相关的有丁酸球菌属、理研菌属和Harryflintia。丁酸球菌属是可以产生丁酸盐的一种肠道益生菌,而丁酸盐可以促进机体的生长发育。有研究表明,产丁酸的丁酸梭菌可以通过调节腿肌肌激素敏感脂肪酶和脂蛋白脂酶等脂肪代谢相关酶活性以及甘油三酯脂肪酶mRNA表达量起到调控脂肪沉积的作用[23]。机体的生长发育与各种代谢变化密切相关,其中尤以脂肪代谢较为旺盛[24]。本试验结果表明,胸肌总胆固醇含量与丁酸球菌属呈显著负相关,表明丁酸球菌属能够促进胸肌胆固醇的代谢。Dielma是一种目前报道较少的菌属,在人的粪便中首次被发现[25]。脱硫弧菌属在鸡的盲肠中起着氢库的作用,而氢有抑制短链脂肪酸生成的作用[26]。张培培[27]发现脱硫弧菌属与体重、腰围呈显著正相关,与体脂分布存在潜在联系,和本试验中脱硫弧菌属与胸肌甘油三酯含量呈显著正相关的结果一致。本试验中,理研菌属与腿肌甘油三酯含量呈显著负相关,表明理研菌属对腿肌甘油三酯的代谢有促进作用。研究表明,理研菌属与脂肪代谢、肥胖等疾病密切相关[28]。此外,肠道微生物代谢产物短链脂肪酸是畜肉脂肪合成过程中最重要的前体物质,其含量不仅显著影响肠道微生物的组成,还直接通过影响胸肌、腿肌中脂肪的含量[29]。硫辛酸通过肠道菌群对胸肌和腿肌脂肪的沉积也可能存在组织特异性,因而表现出胸肌和腿肌总胆固醇和甘油三酯含量变化的差异,且这种差异性变化可能与脂肪代谢相关酶活和基因表达有关[23],其详细机理有待进一步研究。
4 结 论
① 饲粮添加400 mg/kg硫辛酸对肉鸡生长性能无显著影响,但可提高肉鸡28日龄屠宰率以及42日龄胸肌率和胸肌脂肪沉积,降低42日龄全净膛率以及腿肌脂肪沉积。
② 饲粮添加400 mg/kg硫辛酸可以提高肉鸡28日龄盲肠不可培养菌、Shuttleworthia、肠球菌属和理研菌属的相对丰度,提高42日龄盲肠脱硫弧菌属和Dielma的相对丰度。
③ 硫辛酸调控肉鸡胸肌和腿肌的脂肪沉积作用与肠道菌群存在显著相关性。
猜你喜欢
今日农业(2022年4期)2022-11-16
基层中医药(2022年5期)2022-10-24
中国畜牧杂志(2022年6期)2022-06-13
农业工程学报(2022年1期)2022-03-25
现代农村科技(2022年1期)2022-01-21
中国药学药品知识仓库(2021年18期)2021-02-28
现代畜牧科技(2018年7期)2018-10-21
科学种养(2018年7期)2018-09-13
医学信息(2017年1期)2017-02-28
少儿科学周刊·少年版(2015年11期)2015-12-17
