
母猪碳水化合物营养研究进展
2022-10-12 01:51:02吴德卓勇花伦
动物营养学报 2022年9期
吴 德 卓 勇 花 伦
(1.四川农业大学动物营养研究所,成都611130;2.动物抗病营养教育部重点实验室,成都611130)
碳水化合物(carbohydrate,CHO)即多羟基醛或多羟基酮以及水解能产生这类化合物的物质的总称,在常规养分分析中属无氮浸出物和粗纤维范畴。CHO对母猪繁殖具有重要作用,不仅是繁殖活动所需能量的主要来源,而且通过物理和化学效应参与肠道健康、采食调控、免疫调节等多种生理过程。CHO来源广泛、成分复杂,本文综述了本实验室和其他国内外母猪营养领域专家的研究成果,深入解析不同类型CHO及其代谢信号对母猪繁殖活动的调控效应及机制,不仅有利于提高母猪的繁殖效率,而且可为精准利用CHO资源提供重要参考依据。
1 CHO对后备母猪繁殖性能的影响
1.1 可消化CHO对母猪情期启动和卵泡发育的影响
作为饲粮能量的主要来源,可消化CHO不仅为母猪提供生长发育所需ATP,而且刺激代谢激素和繁殖激素分泌,促进初情启动和卵泡发育。饲喂母猪含30.5%淀粉的饲粮,餐后血液葡萄糖浓度峰值比饲喂含14.9%淀粉的饲粮高50%,餐后6 h的胰岛素浓度曲线下面积(area under curve,AUC)提高63%[1]。在70~100 kg体重的后备母猪饲粮中添加30%葡萄糖,母猪血液中胰岛素、雌二醇、促卵泡激素(follicle-stimulating hormone,FSH)、促黄体素(luteinizing hormone,LH)分泌显著提高,初情日龄提前26 d[2]。降低饲粮淀粉水平、增加不可消化CHO含量,后备母猪血清胰岛素样生长因子1(insulin like growth factor 1,IGF1)、瘦素、LH的分泌降低[3]。比较淀粉或油脂对后备母猪生长性能、初情启动的影响,淀粉组母猪血液和卵泡液中的IGF1浓度提高,卵母细胞的体外成熟率(75.4%)比油脂组提高7.4个百分点[4]。提高饲粮淀粉与蛋白质比例,母猪初级卵泡和次级卵泡数量分别增加5.4和6.1个百分点,并通过肝脏成纤维细胞生长因子21和卵巢哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)信号通路抑制原始卵泡过度激活,减少卵泡凋亡,延长繁殖寿命[5]。
可消化CHO促进母猪情期启动机制与其调节下丘脑-垂体-性腺轴活性有关。下丘脑表达KiSS-1基因的神经元是控制母猪情期启动和卵泡发育的关键,而KiSS-1神经元受CHO代谢信号调控。可消化CHO消化生成葡萄糖,刺激胰腺和肝脏分泌胰岛素和IGF1,多余葡萄糖转化为脂肪并提高瘦素的分泌,上述代谢激素通过KiSS-1神经元上的相关受体上调mTOR信号促进Kisspeptin蛋白释放,后者促进促性腺激素释放激素(gonadotrophin releasing hormone,GnRH)、FSH、LH脉冲分泌[6-7],从而调控母猪情期启动和卵泡发育(图1)。下丘脑存在葡萄糖响应中枢,也可促进mTOR信号调节GnRH分泌[8]。同时,葡萄糖及胰岛素、IGF1、瘦素等代谢信号,可直接作用于卵巢mTOR信号促进蛋白质翻译、细胞器组装,同时上调碳水化合物反应元件结合蛋白(carbohydrate response element binding protein,ChREBP)、甾醇调节元件结合蛋白(sterol regulatory element binding protein,SREBP)调节脂质合成等过程(图1),促进原始卵泡的激活以及生长卵泡发育。卵泡发育过程中,葡萄糖经糖酵解、戊糖磷酸盐途径、氨基己糖途径和多元醇途径,为卵母细胞发育提供透明质酸、嘌呤核苷酸、烟酰胺腺嘌呤二核苷酸(NADPH)等底物,对于维持卵母细胞氧化还原平衡和发育潜能十分重要[9]。
尽管可消化CHO及葡萄糖对猪的卵母细胞发育具有重要作用,但葡萄糖浓度过高诱导猪卵内质网应激,对其发育潜能产生不利影响[10]。葡萄糖可上调mTOR信号通路促进卵泡发育[11],但mTOR信号过高会导致原始卵泡过度激活与闭锁,引起卵泡库耗竭和卵巢早衰[12]。肥胖妇女易发生卵巢早衰,原因之一在于高血糖持续激活mTOR信号导致卵的数量和质量下降[13]。新生仔猪卵母细胞数高达百万,但根据单位体积卵巢组织上卵母细胞数量推算,母猪第三情期时卵母细胞数量仅5万~10万[14-15],猪卵在青春期的过度损失值得关注。
1.2 不可消化CHO对母猪情期启动和卵泡发育的影响
饲粮中主要的不可消化CHO是纤维,纤维对后备母猪的影响体现在3个方面:一是通过物理和化学效应促进肠道发育。后备母猪在90~135 kg体重阶段采食高水平纤维,在不改变体重前提下,增加胃重量20%、小肠重量9%、大肠重量19%[16]。二是降低可消化CHO的消化,降低葡萄糖和胰岛素脉冲。母猪在后备期采食高纤维,采食后6 h内的葡萄糖AUC降低8%、胰岛素AUC降低25%[16]。三是通过肠道微生物的代谢效应,调控卵泡成熟和质量。在玉米-豆粕型饲粮基础上添加膳食纤维包(由菊粉和纤维素按1∶4比例组成),母猪的生长性能和初情启动虽不受影响,但随纤维水平的增加(比对照组提高0.5、0.75、1倍),卵泡闭锁数量最高减少50%,总卵泡数最高增加49%,卵母细胞体外发育成熟率最高增加13个百分点[15,17]。高能饲粮虽然促进母猪初情启动,但卵泡闭锁增加112%,而膳食纤维可保护高能饲粮引起的卵泡闭锁[14],缓解高能饲粮对母猪乳腺发育带来的不良效应[18]。后备母猪在配种前19 d采食富含甜菜粕的高纤维饲粮,其妊娠28 d的胚胎存活率比对照组提高10.2%,而增加饲喂水平或蛋白质摄入等营养手段均不能改善母猪早期胚胎存活率[19],高纤维提高母猪早期胚胎存活的机理在于卵母细胞质量的改善[20]。在后备母猪饲粮中添加功能性膳食纤维(含17.4%鼠李糖、4.1%岩藻糖、11.1%阿拉伯糖、30.6%木糖、16.4%半乳糖等),母猪第1胎有效活仔数提高趋势,宫内发育迟缓发生率显著下降[21];同时,母猪第3~5胎产活仔猪数提高1.2~1.5头/窝[22],证实后备期纤维营养可能通过提高卵泡数量和质量改善母猪终身繁殖性能。
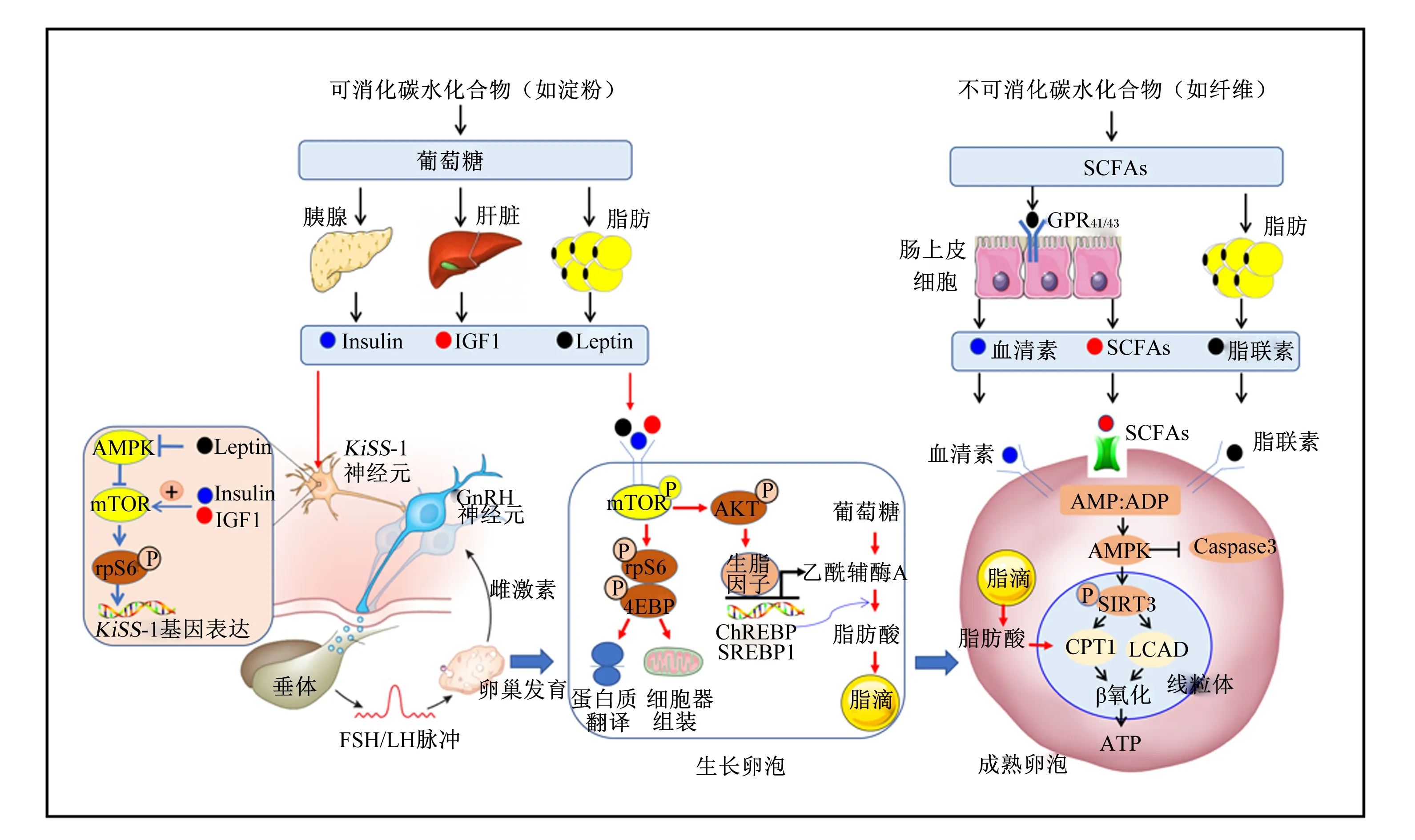
可消化CHO产物代谢信号葡萄糖、胰岛素、IGF1等作用于中枢KiSS-1神经元增强GnRH/FSH/LH脉冲分泌,或作用于生长卵泡mTOR信号促进蛋白质翻译、细胞器组装和脂质合成,调控情期启动和卵泡发育;不可消化CHO产物SCFAs及其代谢信号血清素等,作用于卵内AMPK信号通路抑制凋亡蛋白Caspase-3表达,或启动脂肪酸β-氧化供能为卵母细胞成熟和胚胎发育供能。SCFAs:短链脂肪酸;GPR41/43:G-蛋白偶联受体41/43;mTOR:哺乳动物雷帕霉素靶蛋白;GnRH:促性腺激素释放激素;FSH:促卵泡激素;LH:促黄体素;AMPK:激活腺苷酸活化蛋白激酶;Caspase-3:天冬氨酸特异性半胱氨酸蛋白酶-3;SIRT3:Sirtuin蛋白3;IGF1:胰岛素样生长因子1;Insulin:胰岛素;Leptin:瘦素;rpS6:核糖体蛋白S6;AKT:丝氨酸-苏氨酸蛋白激酶;CPT1:肉碱棕榈酰基转移酶1;LCAD:长链酰基辅酶A脱氢酶;ChREBP:碳水化合物反应元件结合蛋白;SREBP1:甾醇调节元件结合蛋白。
不可消化CHO产生的乙酸、丙酸、丁酸等短链脂肪酸(short-chain fatty acids,SCFAs),可显著上调细胞内AMP∶ADP比例,激活腺苷酸活化蛋白激酶(AMP-activated protein kinase,AMPK)信号通路,启动细胞内的分解代谢过程,抑制促凋亡因子天冬氨酸特异性半胱氨酸蛋白酶-3(cysteine aspartic acid-specific protease 3,Caspase-3)表达,并通过增强细胞抗逆特性改善卵泡发育质量及繁殖组织的健康水平(图1)[23]。随纤维水平增加,母猪卵巢磷酸化AMPK蛋白表达水平线性增加,促凋亡因子B细胞淋巴瘤2相关X蛋白(B-cell lymphoma-2 associated X protein,BAX)和Caspase-3的蛋白表达线性下降[15,17]。猪是卵内脂质含量最高的种属,也是对脂肪酸β-氧化最敏感的种属,研究已证实,卵内AMPK信号激活并启动脂肪酸β-氧化过程是卵子成熟和早期胚胎发育的必需过程[24]。多组学分析发现,血清素代谢是饲粮纤维影响后备母猪最显著的代谢途径之一[14,17]。血清素作为一种“愉快”激素,95%来自肠道分泌,是褪黑素合成前体,不仅影响动物情绪,而且影响卵母细胞成熟和早期胚胎存活[25]。脂联素是受到纤维调控的重要脂肪分泌因子[26],也可能参与纤维调控卵母细胞成熟。
因此,可消化CHO产生葡萄糖及胰岛素等代谢信号,通过mTOR信号调节促进情期启动和卵泡发育;不可消化CHO代谢产生SCFAs,通过AMPK信号降低卵泡闭锁,提高卵母细胞质量。生产中后备母猪通常按维持+生长设定营养需要,忽视卵巢等繁殖组织的需求。结合上述研究进展,后备母猪饲粮应当充分考虑两类CHO的适宜配比。
2 CHO对妊娠母猪繁殖性能的影响
2.1 可消化CHO对妊娠母猪胚胎附植与胎儿发育的影响
可消化CHO与孕酮互作调节胚胎附植与发育,同时通过调节印迹基因表达影响胎盘养分转运与胎儿宫内生长。妊娠饲粮添加32%的玉米淀粉,母猪死胎数显著降低,初生重为0.8~1.0 kg仔猪比例显著减少[27]。比较妊娠85 d至分娩阶段饲喂添加32%玉米淀粉、3.0%大豆油和3.0%鱼油的3种饲粮对母猪繁殖性能的影响,结果显示各组母猪产仔性能差异不显著,但与大豆油组或鱼油组相比,玉米淀粉组母猪产程缩短100~170 min、死胎率降低5.7~10.6百分点、仔猪初生重提高100 g[28]。葡萄糖代谢对母猪胚胎附植与胎儿发育具有重要作用。猪胚胎在附植窗口期,输卵管和子宫液中葡萄糖浓度显著增加[29]。葡萄糖不仅为胚胎附植与发育提供ATP,且葡萄糖在孕酮作用下,通过上调磷酸果糖激酶1(fructose-1,6-bisphosphatase 1,PFK1)促进中间代谢产物1,6-二磷酸果糖在子宫内膜组织富集,1,6-二磷酸果糖调节子宫内膜基质细胞分泌白细胞介素-27(interleukin-27,IL-27),促进M2型巨噬细胞分化及基质细胞蜕膜化,增加子宫容受性,促进胚胎附植(图2)[30]。
葡萄糖代谢产生乳酸盐、丙酮酸盐、嘌呤核苷酸前体、多元醇等,是早期胚胎发育过程中细胞增殖与分化、氧化还原平衡的必需营养底物[29,31]。此外,葡萄糖可在转运载体溶质载体家族2成员1(solute carrier family 2 member 1,SLC2A1)和溶质载体家族2成员3(solute carrier family 2 member 3,SLC2A3)协助下被子宫内膜或孕体组织摄取,经磷脂酰肌醇3激酶(phosphatidylinositol 3-kinase,PI3K)/丝氨酸-苏氨酸蛋白激酶(serine-threonine kinase,AKT)信号通路调节合成糖原[32-33],可提高猪胚胎囊胚率及多功能干细胞的增殖分化潜能[34]。葡萄糖在孕体组织转化为果糖,附植期子宫液、羊水中的果糖浓度是葡萄糖的10~30倍,果糖通过氨基己糖途径、一碳单位代谢途径、NADH途径改善猪的早期胚胎发育[35]。
2.2 不可消化CHO对妊娠母猪胚胎附植与胎儿发育的影响
纤维等不可消化的CHO对妊娠母猪具有重要作用,一是通过物理效应增加母猪胞感,减少便秘等胃肠道疾病;二是调节免疫平衡,促进胚胎附植;三是调节胎盘养分转运及胎儿生长,降低窝内变异系数。纤维来源广、成分杂,对母猪繁殖性能的影响存在差异[36-38]。研究发现,以大豆皮纤维为纤维来源时,早期胚胎存活率(65.1%)显著低于燕麦纤维组(80.3%)[39]。以麦麸为纤维源将中性洗涤纤维水平由10.8%分别提升至15.8%和20.8%,发现初产母猪饲喂低纤维时总产仔数高于中、高水平纤维组,同时增加窝内变异系数;但上述日粮处理持续至二胎时,中纤维水平组母猪总产仔数比低、高纤维组分别提高0.9和1.1头[40],暗示纤维对妊娠母猪产仔性能的影响与胎次有关。妊娠饲粮以甜菜粕为纤维源时,母猪产仔性能优于麦麸和大豆皮[39,41]。Renteria-Flores等[39]提出,不可溶/可溶纤维(insoluble/soluble fiber,ISF/SF)比例可能是导致不同纤维源对妊娠母猪营养效应产生差异的原因。常规玉米-豆粕型饲粮中ISF/SF比例超过10∶1,通过在妊娠饲粮中添加SF降低ISF/SF比例至4∶1~8∶1时,母猪的产仔性能、泌乳性能、糖代谢、脂代谢等显著改善[42-46]。以瓜尔豆胶为纤维源提高饲粮中可溶性纤维比例,发现母猪总纤维水平达到18.8%(且ISF/SF=6.3)时,产活仔猪数最高[47]。
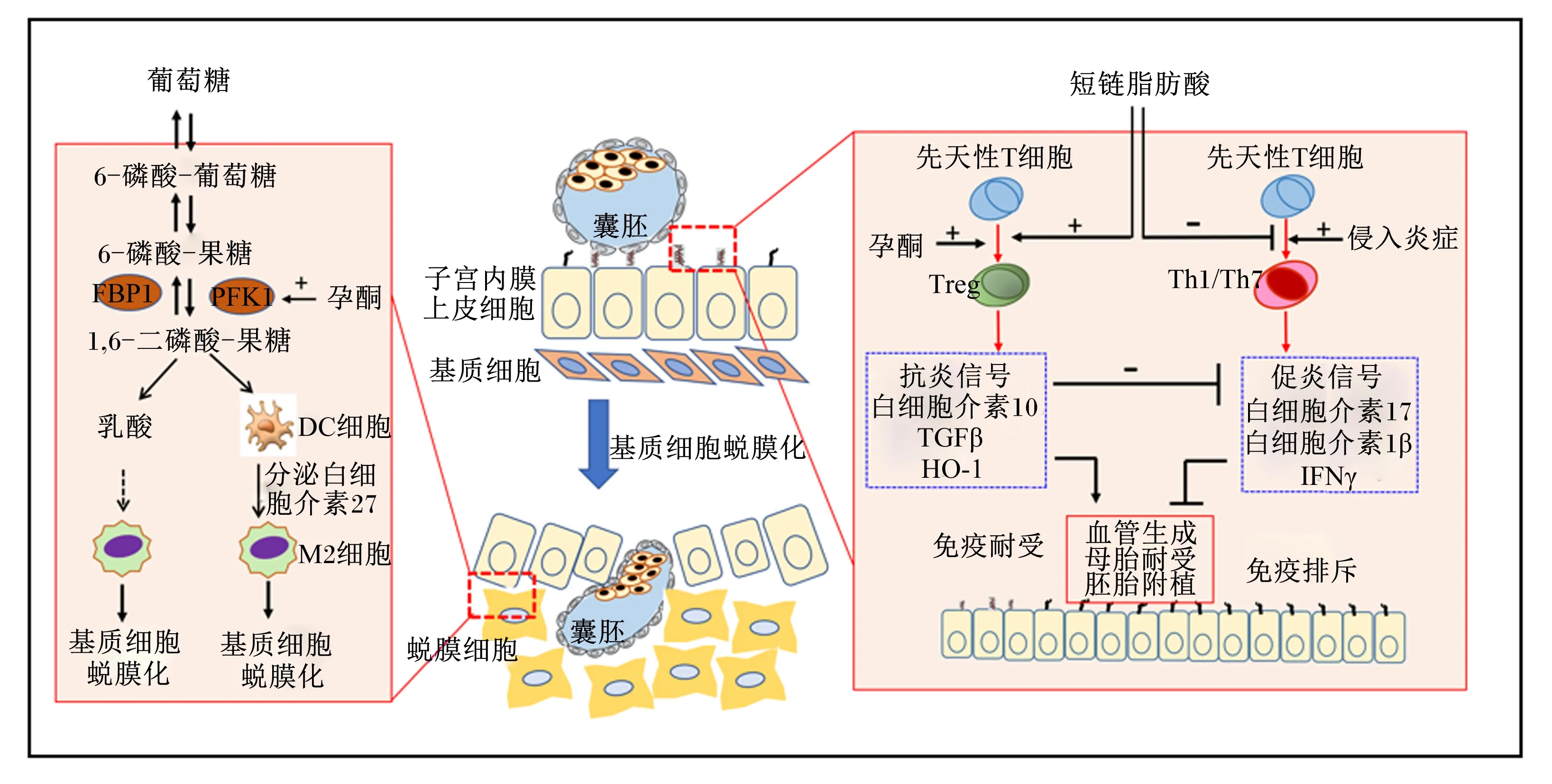
葡萄糖代谢参与子宫内膜基质细胞蜕膜化,增加子宫容受性;SCFAs促进Treg细胞分化,降低促炎反应,增加免疫耐受。PFK1:磷酸果糖激酶1;FBP1:果糖-1,6-二磷酸酶1;Treg:调节性T细胞;TGFβ:转化生长因子β;HO-1:血红素氧合酶-1;IFNγ:干扰素γ。
SCFAs是纤维首要代谢产物,纤维提高胚胎存活及产仔性能的机制与免疫有关。胚胎作为外源移植体,易被母体免疫系统攻击导致胚胎死亡,母-胎界面调节性T细胞(regulatory T,Treg)细胞是母体免疫系统对胎儿形成免疫耐受的关键,是胚胎附植的生理基础[48]。而纤维及其代谢产物SCFAs显著促进Treg细胞分化,是机体对食物过敏原、移植器官等异源物质形成免疫耐受的关键[49]。在小鼠上的研究发现,乙酸通过影响母-胎界面的T细胞分化促进母胎耐受,改善胚胎附植与发育[50]。研究表明,随着纤维水平的升高,母猪妊娠30 d时血液Treg细胞分泌因子白细胞介素-10(interleukin-10,IL-10)的浓度线性增加,且产仔数与血液中IL-10浓度呈正相关关系,妊娠30 d时血液中IL-10浓度由152 ng/L增加至248 ng/L时,母猪产活仔数由12.1头提高至13.9头[47],暗示纤维通过T细胞介导的免疫耐受机制调节胚胎附植与产仔性能(图2)。
在NRC(2012)营养水平基础上降低妊娠期25%的淀粉摄入,仔猪初生重降低100 g,骨骼肌降低5 g,28日龄断奶重降低520 g,189日龄体重降低3.3 kg,而提高淀粉摄入水平并不会改善肌纤维发育及出生后的生长[51]。妊娠中后期,胎盘印迹基因是调控葡萄糖转运的关键。基于印迹基因Igf2-Igf2r胎盘特异性敲除的小鼠模型,发现印迹基因是控制母体微血管系统、葡萄糖转运的关键信号通路[52]。在猪上的研究发现,印迹基因Neuronatin表达量的高低直接影响葡萄糖转运载体1(glucose transporter 1,GLUT1)表达,最终影响猪的初生重[53]。高淀粉饲粮通过葡萄糖和胰岛素上调父源印迹基因Igf2表达,促进胎儿生长,增加窝内变异系数;进一步研究证实,葡萄糖上调胚胎成纤维细胞环状RNA circ-Amotl1的表达,通过“海绵吸附”效应抑制miR-17-5p表达,降低后者对Igf2基因的转录抑制效应,从而促进Igf2基因表达[54]。而高纤维饲粮通过SCFAs及甲基化修饰上调母源印迹基因H19表达,降低窝内变异系数(图3)[40,55-56]。
因此,可消化和不可消化CHO分别调节子宫蜕膜化和母-胎免疫平衡调节胚胎附植,并通过影响胎盘印迹基因表达调控胎盘养分转运及胎儿生长。目前,大量研究对妊娠母猪饲粮中纤维水平关注较多,但对淀粉等可消化CHO的水平关注偏少。解析现有文献[27-28,36-38]及本课题组研究[40-42,47],在母猪妊娠饲粮中含18%~22%纤维(ISF/SF比例在4∶1~8∶1)时,推荐饲粮淀粉含量在30%~35%。

M:母源 maternal;P:父源 paternal;DNMTs:甲基转移酶 methyltransferase;ICR:印迹控制区域 imprinted control region;CTCF:CTCF基因转录因子 CTCF transcription factor。
3 CHO对围产期和泌乳期母猪繁殖性能的影响
3.1 可消化CHO对围产期和泌乳期母猪繁殖性能的影响
母猪在围产期提高基础能量代谢,以促进子宫收缩和初乳合成,可消化CHO产物葡萄糖对母猪围产期的繁殖活动十分重要。基于子宫动静脉插管技术,发现子宫在分娩前会大量摄取血液葡萄糖用于产前缩宫[57],乳腺组织也会摄取大量葡萄糖用于乳腺增殖和初乳合成[58]。在分娩开始后的前6 h,乳腺组织摄取的葡萄糖和乳酸比分娩前增加1倍;母猪宫缩分娩时,葡萄糖代谢产生大量乳酸,后者被乳腺摄取,占分娩初期食入总碳的15%[59]。围产期母猪首先利用血液中葡萄糖,当血糖不足时,机体分解脂肪供能。对分娩母猪进行外源葡萄糖灌注处理,发现母猪死胎率由16.1%降至7.4%,助产比例由21%降至9.0%[60]。妊娠85 d~分娩阶段,将饲粮淀粉/脂肪比例由10∶1提高至15∶1,有助于维持母-仔猪脂质代谢稳态,提升母-仔猪健康水平[61]。泌乳饲粮添加5%葡萄糖,母猪泌乳期体损失显著减少,哺乳仔猪增重提高,母猪断奶-发情间隔缩短[62]。同时,饲粮结构影响母猪乳腺对葡萄糖的摄取,母猪饲喂粗蛋白质含量为13%的饲粮时,乳腺对葡萄糖的摄取高于饲喂粗蛋白质含量为18.2%和23.5%的饲粮[63]。
3.2 不可消化CHO对围产期和泌乳期母猪繁殖性能的影响
不可消化CHO延长围产期母猪的供能时间,同时通过物理和化学效应保障胃肠道功能,有利于母猪泌乳期采食量的提升。母猪分娩前食欲下降,可消化CHO产物葡萄糖虽快速为母猪供能,但持续时间短(图4)。围产期饲粮添加高水平纤维不仅减缓葡萄糖的消化吸收,而且在后肠持续发酵供能(图4),可缩短产程,减少死胎率25%[64]。与快发酵的甜菜粕纤维相比,妊娠后期至泌乳期饲粮添加高水平大豆皮,母猪产程缩短,死胎减少,抗氧化能力增强,其机制在于大豆皮纤维提高丙酸产生菌的丰度,显著改善母猪能量代谢[65-67]。
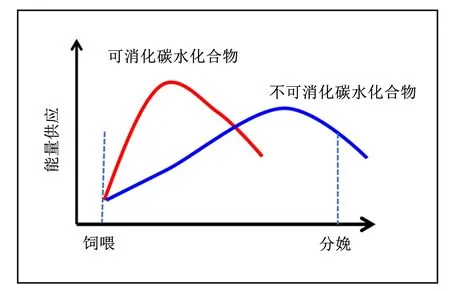
图4 不同类型碳水化合物对母猪供能的时间顺序
总结相关研究结果发现,母猪妊娠期采食高水平可消化CHO会增加体脂沉积,抑制泌乳期采食量,即出现“高妊娠低泌乳”现象[36-38]。而妊娠期采食高纤维饲粮可提高泌乳期采食量,可能机理如下:一是高纤维促进胃肠道发育,使母猪具有更大的胃肠道容积,为采食量的提高奠定生理基础[16];二是妊娠期高纤维降低脂肪细胞分泌的瘦素水平,减轻瘦素对母猪的抑食效应[68];三是纤维抑制母猪肠道中大肠杆菌、产气荚膜梭菌等有害微生物增殖[69],可能降低大肠杆菌分泌肽如酪蛋白水解蛋白酶B蛋白质同族体(caseinolytic peptidase B protein homolog,Clpb)等因子对采食中枢的抑制效应[70](图5)。此外,华中农业大学彭健课题组在纤维发酵特性与母猪食欲调节方面开展大量工作[69,71-72],该课题组研究发现纤维发酵产生的SCFAs可通过G-蛋白偶联受体(G protein-coupled receptor,GPR)信号途径提高抑食欲因子[如肽YY(PYY,peptide YY)、胰高血糖素样肽-1(glucagon-like peptide-1,GLP-1)等]的浓度,上述途径不仅有利于提高妊娠母猪胞感,而且对泌乳期采食量的提升具有重要作用。
因此,可消化CHO对围产期和泌乳期母猪能量平衡十分重要,而不可消化CHO不仅与可消化CHO协同维持围产母猪的血糖稳态,且通过物理和化学效应提高泌乳期采食量。在遗传育种持续提高产仔性能背景下,优化母猪在围产期的能量供给模式对于缩短产程、降低死胎具有重要作用。
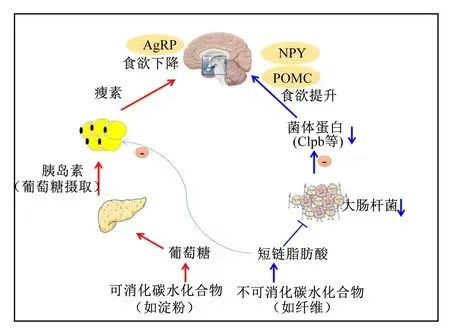
AgRP:刺鼠相关蛋白 agouti-related protein;NPY:神经肽Y neuropeptide Y;POMC:前阿黑皮素原 prepromelanocortin;Clpb:酪蛋白水解蛋白酶B蛋白质同族体 caseinolytic peptidase B protein homolog。
4 小结与展望
综上可知,可消化CHO的产物葡萄糖促进胰岛素、IGF1、瘦素等代谢激素的分泌,是机体合成代谢的信号,是保障母猪卵母细胞成熟、胚胎与胎儿发育、泌乳等繁殖活动顺利进行的营养基础;不可消化CHO不仅具有物理效应,而且代谢产生SCFAs,对于提升母猪的健康水平具有重要作用。目前,针对母猪碳水化合物营养体系的构建尚处于起步阶段,以下理论或技术难点亟待突破:1)精准解析饲料原料中的淀粉、寡糖、纤维等CHO组分含量,完善针对母猪的CHO营养价值数据库;2)确定母猪不同繁殖生理阶段单糖、二糖、寡糖、淀粉、非淀粉多糖等CHO的最适添加水平,以及不同CHO组分之间的信号交联作用(如纤维调控机体葡萄糖稳态);3)揭示不同CHO与蛋白质和脂质等其他营养素代谢的互作关系,及其对母猪繁殖效率的调控机制;4)阐明肠道微生物对不同CHO组分响应规律,及不同CHO组分的微生物靶向特性,为母猪饲粮精准利用CHO提供依据
猜你喜欢
动物营养学报(2022年8期)2022-09-01 03:20:52
保健与生活(2020年18期)2020-09-26 12:19:37
解放军医学杂志(2020年7期)2020-09-02 06:03:18
河南畜牧兽医(2017年8期)2017-11-24 03:21:44
动物营养学报(2017年8期)2017-08-16 09:40:00
上海农业学报(2017年3期)2017-04-10 12:39:26
中国卫生标准管理(2015年25期)2016-01-14 09:29:32
动物营养学报(2015年9期)2016-01-07 11:29:44
科技视界(2014年29期)2014-08-15 00:54:11
生殖医学杂志(2013年5期)2013-03-11 18:43:47
