
桑叶发酵工艺优化及营养价值评价
2022-10-12 03:24:06崔艺燕王超普田志梅鲁慧杰刘志昌马现永
动物营养学报 2022年9期
崔艺燕 王超普 彭 苏 邓 盾 田志梅 鲁慧杰 余 苗 刘志昌* 马现永,2*
(1.广东省农业科学院动物科学研究所,畜禽育种国家重点实验室,农业农村部华南动物营养与饲料重点实验室,广东省畜禽育种与营养研究重点实验室,广东畜禽肉品质量安全控制与评定工程技术研究中心,广州510640;2.岭南现代农业科学与技术广东省实验室茂名分中心,茂名525000)
单宁是复杂的水溶性多酚化合物,存在于许多植物中,并可存在于树皮、树叶、树干、果实和根中[1]。单宁能够络合蛋白质、氨基酸、大分子物质,对消化酶产生拮抗作用,阻碍营养物质的吸收,降低动物的消化率[2];并因其苦涩的味道减少动物的采食量,从而降低植物饲料的营养价值[3-4]。桑叶资源丰富,富含粗蛋白质(CP)、氨基酸、维生素、矿物质[5],是一种较好的非常规饲料资源。但桑叶单宁含量较高,限制了其在畜牧业的应用。因此,有效降解单宁对提升桑叶的应用价值具有重要意义。
物理加工技术可以有效地降解单宁,如高温、浸泡、煮沸、高压、挤压蒸煮等[6-8]。尽管上述技术可以降解一些单宁,但也可能降低桑叶营养成分。另外,上述技术耗能成本较高,不利于可持续发展。因此,需要一种更合适的、高效降解桑叶单宁的技术。发酵是利用微生物将植物茎叶成分转化为微生物蛋白、活性小肽、活性物质的技术,能降解抗营养因子,提高饲料原料的营养价值[9]。黑曲霉、地衣芽孢杆菌、酵母、乳酸菌可以高效地将高分子单宁降解为小分子物质[2,10-12]。柔毛镰孢菌和球孢白僵菌等丝状真菌发酵显著减少了橄榄饼单宁含量[13]。黑曲霉、产朊假丝酵母、枯草芽孢杆菌发酵降低了辣木叶41%单宁含量,提高了44% CP含量[14]。罗伊氏乳杆菌、乳酸片球菌、枯草芽孢杆菌发酵降低了朱缨花叶单宁含量[15]。这些文献为发酵降解桑叶单宁提供了思路。但是,关于桑叶发酵工艺的研究很少,通过发酵技术降解桑叶单宁的研究更少,尚不清楚发酵条件如何影响桑叶单宁的降解。因此,本研究通过正交试验优化了提高桑叶CP含量同时降低单宁含量的发酵条件,研究了桑叶发酵基质组成、菌种组合、接种量及发酵时间对单宁降解的影响,并且探讨了发酵对桑叶营养成分的影响,以期为桑叶的应用提供参考。
1 材料与方法
1.1 试验材料
桑叶由广东省农业科学院蚕业与农产品加工研究所提供,测得桑叶干物质(DM)、CP、粗脂肪(EE)、单宁含量分别为30.50%、23.67%、2.52%、1.32%。
酒窖片球菌(Pediococcuscellicola)CGMCC1.3787购自中国普通微生物菌种保藏管理中心,地衣芽孢杆菌(Bacilluslicheniformis)和葡萄牙棒孢酵母(Clavisporalusitaniae)均来自广东省农业科学院动物科学研究所。
1.2 主要试剂及仪器
培养微生物使用的MRS肉汤培养基、营养肉汤培养基、马铃薯葡萄糖培养基购自广东环凯微生物科技有限公司。测定指标所用的单宁酸(货号16201)购自西格玛奥德里奇(上海)贸易有限公司,茚三酮(分析纯)购自上海伯奥生物科技有限公司,氨基酸标准品购自日本和光纯药工业株式会社,其余试剂均为分析纯。
测定指标使用的主要仪器有KjeltecTM8400全自动凯氏定氮仪(FOSS公司)、Spectra Max M5多功能酶标仪(Molecular Devices公司)、2055 SOXTEC半自动脂肪测定仪(FOSS公司)、Ankom 220纤维分析仪(Ankom公司)、SX2-4-10马弗炉(上海一恒科技有限公司)、L8900氨基酸分析仪(Hitachi公司)等。
1.3 试验设计和方法
1.3.1 种子液的制备
取0.1 mL酒窖片球菌保存液于100 mL灭菌MRS肉汤培养基中,37 ℃静置培养24 h。取0.1 mL地衣芽孢杆菌保存液于100 mL灭菌营养肉汤培养基中,37 ℃振荡培养24 h。取0.1 mL葡萄牙棒孢酵母保存液于100 mL灭菌马铃薯葡萄糖培养基中,28 ℃振荡培养24 h。用灭菌水调整上述活化后菌液浓度为1.5×106CFU/mL,备用。
1.3.2 发酵基质的制备
将鲜桑叶切短(1~2 cm),按照一定比例添加麸皮混合均匀,按照所需菌种组合和接种量添加菌种并搅拌均匀,发酵基质未灭菌,基质pH自然状态(不调整pH),每个呼吸袋装入200 g发酵基质,密封发酵。在空调房内进行发酵,发酵温度为(25±1) ℃。
1.3.3 正交试验
为了获得高CP含量和低单宁含量的发酵桑叶的优化组合,以桑叶与麸皮质量比(A)、菌种组合(B)、接种量(C)和发酵时间(D)为影响因素,进行L9(34)正交试验,设计依据为前期单因素试验结果。正交试验设计如表1所示,每个因素设3个水平,共9组(T1~T9),每组3个重复。按照正交试验设计,根据1.3.2发酵基质的制备方法进行桑叶发酵,共制备发酵桑叶27袋,每袋200 g发酵基质。
1.3.4 验证试验
根据正交试验得出的组合,进行桑叶发酵,比较最优条件下发酵桑叶和正交试验中最优CP和单宁含量。
1.3.5 对比试验
根据最优组合进行桑叶发酵(10个重复),比较未发酵桑叶和发酵桑叶的营养价值。发酵桑叶制备过程同1.3.2发酵基质的制备,即按照桑叶与麸皮质量比(9∶1)制备20袋发酵基质(200 g/袋),其中10袋发酵基质不发酵,于-20 ℃存放。余下10袋加入2%的酒窖片球菌和地衣芽孢杆菌菌液,(25±1) ℃下发酵4 d。发酵结束后,将样品烘干后进行营养成分的测定。
1.3.6 相关指标测定
CP含量的测定参照GB/T 6432—1994,单宁含量的测定参照文献[16],DM含量的测定参照GB/T 6435—2014,EE含量的测定参照GB/T 6433—2006,粗纤维(CF)、中性洗涤纤维(NDF)、酸性洗涤纤维(ADF)含量的测定参照文献[17],粗灰分含量的测定采用550 ℃灼烧法,钙含量的测定参照GB/T 6436—2018,磷含量的测定参照GB/T 6437—2002,氨基酸含量的测定参照GB/T18246—2000,根据CP、EE、粗灰分含量计算碳水化合物含量[15]:
碳水化合物(%)=100-CP-EE-粗灰分。

表1 正交试验设计
1.4 数据统计分析
采用SPSS 25.0进行一般线性模型(general linear model)或独立样本t检验(independent samplest-test)分析,采用Duncan氏法进行多重比较。结果用平均值和均值标准误(SEM)表示,P<0.05为差异显著,0.05≤P<0.10为有显著趋势。
2 结果与分析
2.1 正交试验极差分析
由表2可知,通过直观分析方法比较表中极差大小,对CP含量影响因素大小排序为:桑叶与麸皮质量比(A)、菌种组合(B)、接种量(C)、发酵时间(D)。根据Ki值的大小判断最优水平,得到各因素对CP含量的最优组合为A1B2C2D2。同理,对单宁含量影响因素大小排序为:菌种组合(B)、发酵时间(D)、桑叶与麸皮质量比(A)、接种量(C),得到各因素对单宁含量的最优组合为A1B2C2D1。综合上述分析,桑叶发酵最优方案为A1B2C2D1或A1B2C2D2,即桑叶与麸皮质量比为9∶1,菌种组合为酒窖片球菌+地衣芽孢杆菌,接种量为2%,发酵时间为4或6 d。

表2 正交试验极差分析
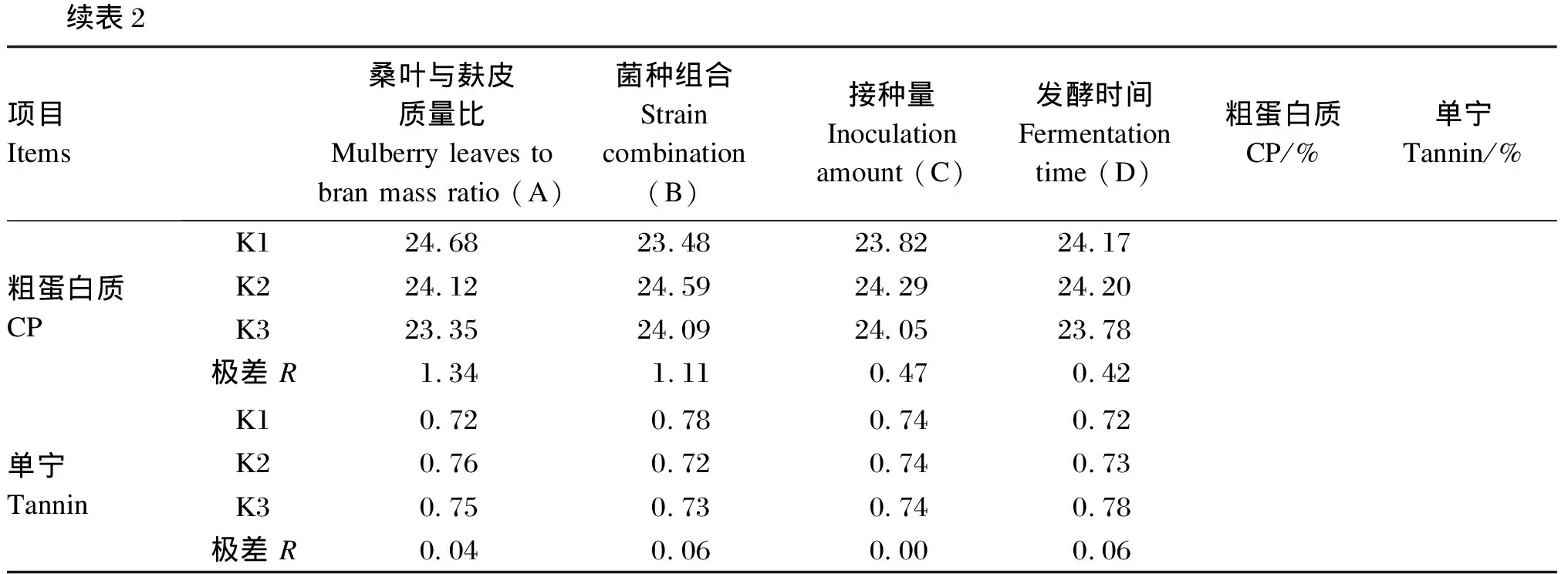
2ItemsMulberry leaves to bran mass ratio (A)Strain combination (B)Inoculation amount (C)Fermentation time (D)CP/%Tannin/%CPK124.68 23.48 23.82 24.17 K224.12 24.59 24.29 24.20 K323.35 24.09 24.05 23.78 R1.34 1.11 0.47 0.42 TanninK10.72 0.780.740.72 K20.76 0.72 0.74 0.73 K30.75 0.73 0.74 0.78 R0.040.06 0.00 0.06
2.2 正交试验方差分析
由表3和表4可知,CP的校正模型P值为0.030<0.05,说明校正模型对试验结果有显著影响,正交试验结果具有可信度。桑叶与麸皮质量比和菌种组合显著影响发酵桑叶CP含量,说明二者是影响发酵桑叶CP含量的主要因素。根据F值得到,对CP含量影响因素大小排序为:桑叶与麸皮质量比、菌种组合、接种量、发酵时间。对桑叶与麸皮质量比(A)分析,A1的CP含量显著高于A3(P<0.05),A2的CP含量与A1、A3差异均不显著(P>0.05),因此选择A1。同理,菌种组合(B)选择B2。桑叶与麸皮质量比(A)和菌种组合(B)对CP含量的最优组合为A1B2。
同理,菌种组合和发酵时间是影响单宁含量的主要因素。菌种组合(B)和发酵时间(D)对单宁含量的最优组合为B2D1。综合CP和单宁含量的最优组合,得到桑叶发酵最优组合为A1B2CiD1。
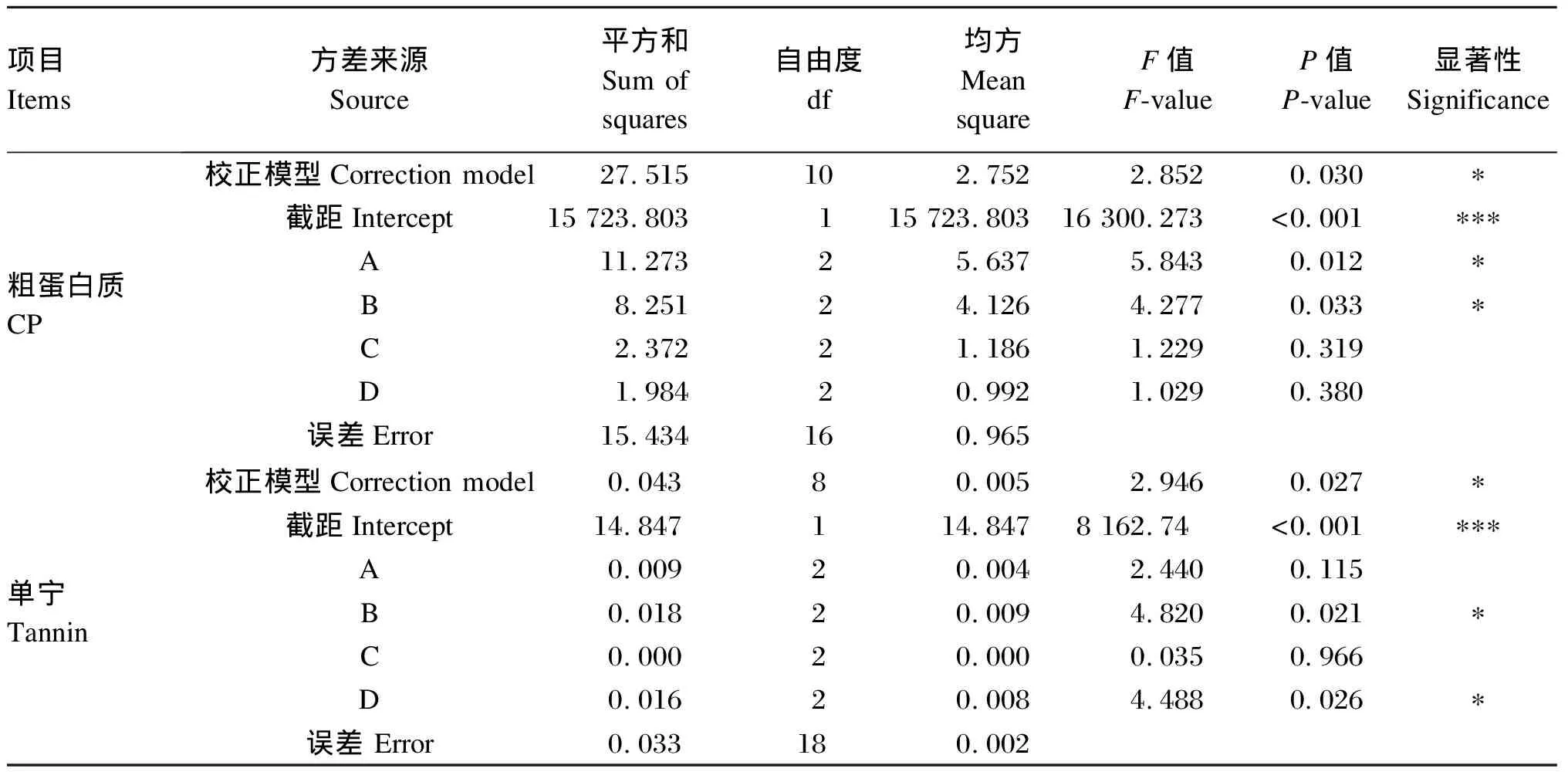
表3 正交试验方差分析
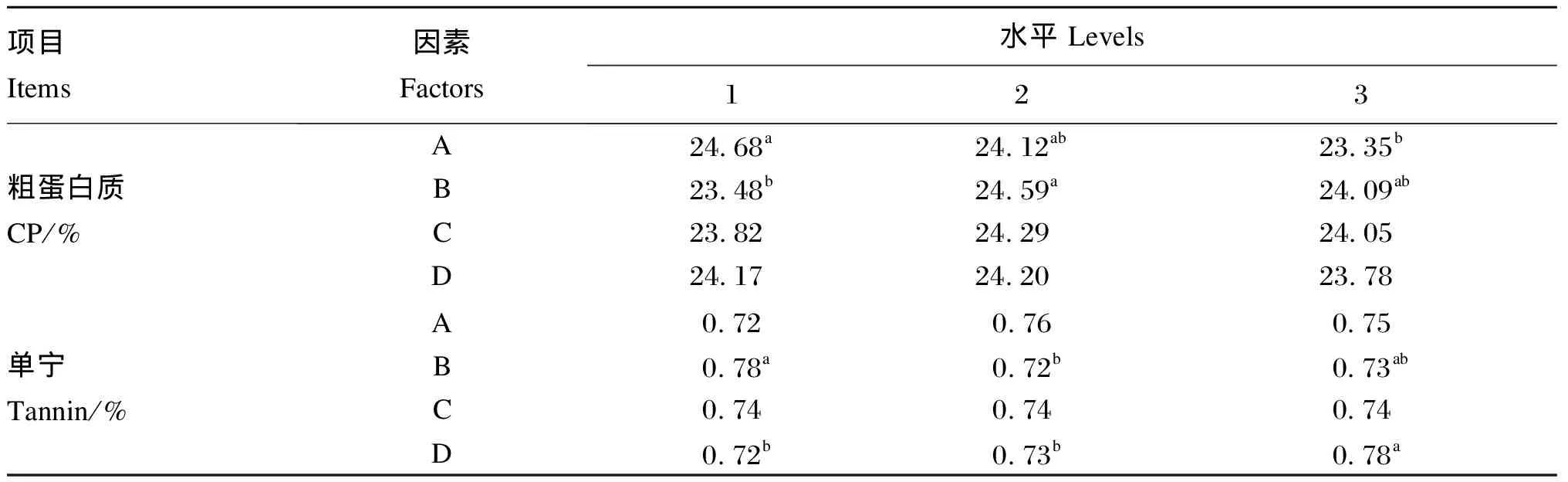
表4 正交试验方差分析多重比较
2.3 验证试验
综合极差分析(A1B2C2D1或A1B2C2D2)和方差分析(A1B2CiD1)结果,桑叶发酵的最优组合为A1B2C2D1,即桑叶与麸皮质量比为9∶1,菌种组合为酒窖片球菌+地衣芽孢杆菌,接种量为2%,发酵时间为4 d。由于分析出来的最优组合A1B2C2D1不在正交试验设计之内,因此对该组合进行了验证试验。测得CP含量为26.18%,高于正交试验表最高CP含量(25.60%);测得单宁含量为0.47%,低于正交试验表最低单宁含量(0.67%),从而验证了桑叶发酵优化组合为A1B2C2D1。
2.4 对比试验
2.4.1 营养成分
发酵桑叶颜色黄绿,有桑叶清香,味道酸甜。由表5可知,发酵桑叶CP和EE含量分别比未发酵桑叶提高了16.82%和30.57%(P<0.05)。发酵桑叶粗灰分和磷含量显著高于未发酵桑叶(P<0.05)。发酵桑叶单宁、NDF、碳水化合物含量分别比未发酵桑叶降低了56.40%、16.78%、7.29%(P<0.05)。发酵桑叶pH显著低于未发酵桑叶(P<0.05)。发酵桑叶与未发酵桑叶之间DM、CF、ADF、钙含量差异不显著(P>0.05)。
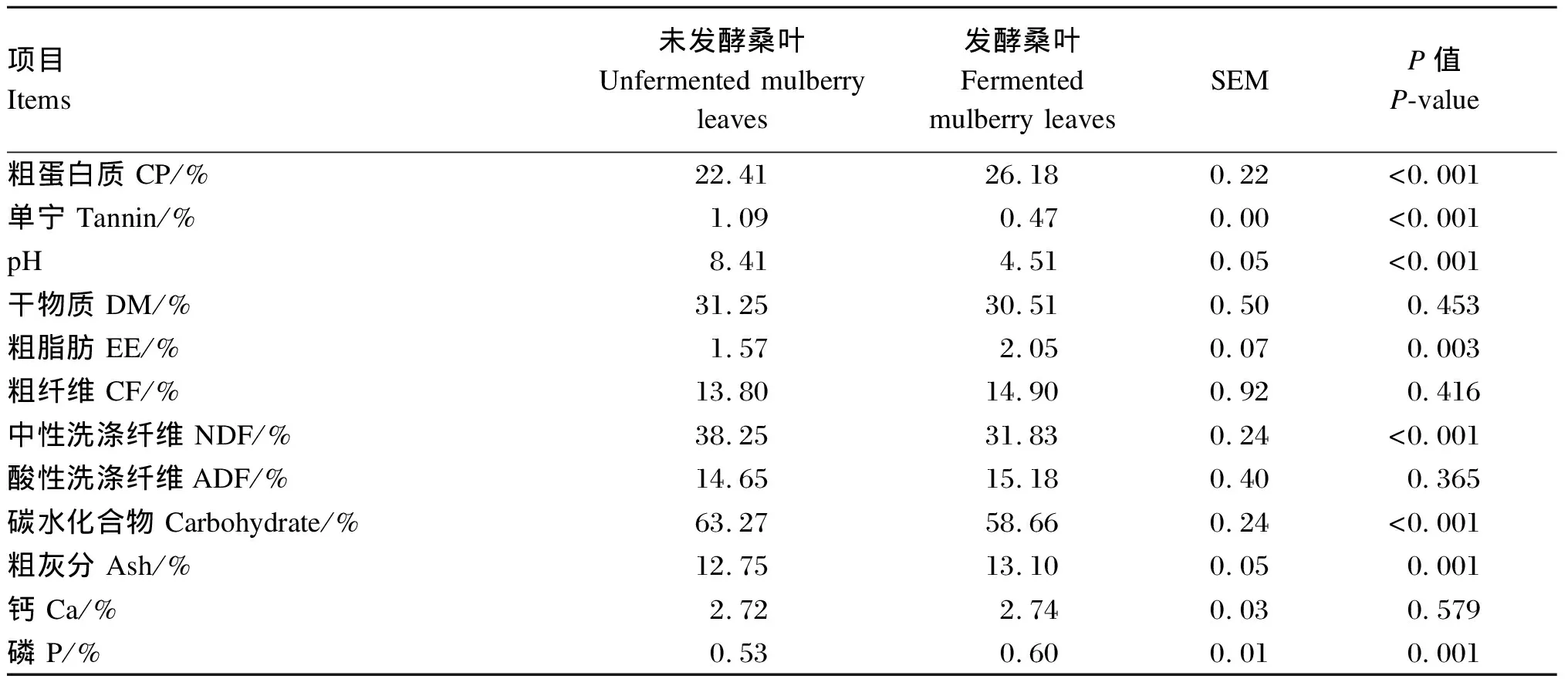
表5 未发酵桑叶和发酵桑叶营养成分对比
2.4.2 氨基酸含量
由表6可知,与未发酵桑叶相比,发酵桑叶精氨酸、丝氨酸、谷氨酸、天冬氨酸、总非必需氨基酸、总氨基酸含量显著降低(P<0.05),而丙氨酸含量、总必需氨基酸/总氨基酸(TEAA/TAA)、总必需氨基酸/总非必需氨基酸(TEAA/TNEAA)显著提高(P<0.05)。
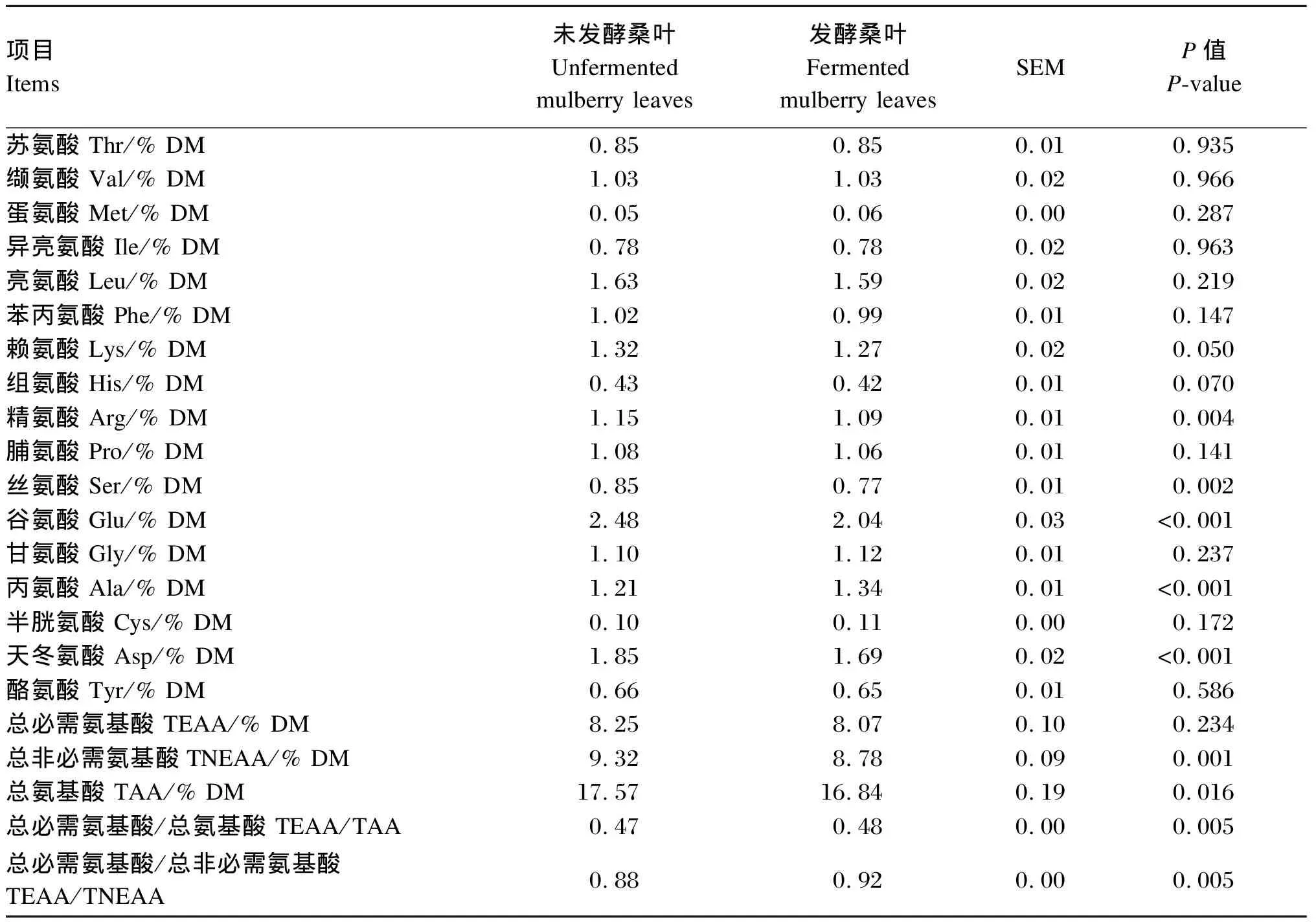
表6 未发酵桑叶和发酵桑叶氨基酸含量对比
2.4.3 必需氨基酸组成
由表7可知,发酵桑叶的半胱氨酸+蛋氨酸、苏氨酸、亮氨酸、酪氨酸+苯丙氨酸组成均显著高于未发酵桑叶(P<0.05)。与未发酵桑叶相比,发酵桑叶缬氨酸组成有提高的趋势(0.05≤P<0.10)。未发酵桑叶和发酵桑叶的赖氨酸和亮氨酸组成高于鸡蛋蛋白和联合国粮农组织(FAO)模式,苏氨酸、缬氨酸、异亮氨酸、酪氨酸+苯丙氨酸和总必需氨基酸组成高于FAO模式,稍低于鸡蛋蛋白模式,半胱氨酸+蛋氨酸组成低于鸡蛋蛋白和FAO模式。由此可见,未发酵桑叶和发酵桑叶的必需氨基酸组成较为均衡,发酵桑叶必需氨基酸组成优于未发酵桑叶。
3 讨 论
3.1 影响桑叶发酵的因素
桑叶与麸皮质量比是影响发酵桑叶CP含量的主要因素。桑叶与麸皮质量比为9∶1时发酵所得的CP含量最高。这与桑叶的本底CP含量和微生物蛋白的合成有关。另外,试验中使用麸皮与新鲜桑叶混合发酵,调节发酵基质的水分含量。微生物生长需要最佳的水分含量,较低的水分含量会降低基质营养物质的溶解度[18]。本试验中A1的发酵基质水分含量高于A2和A3,有利于微生物的生长和营养物质的积累。
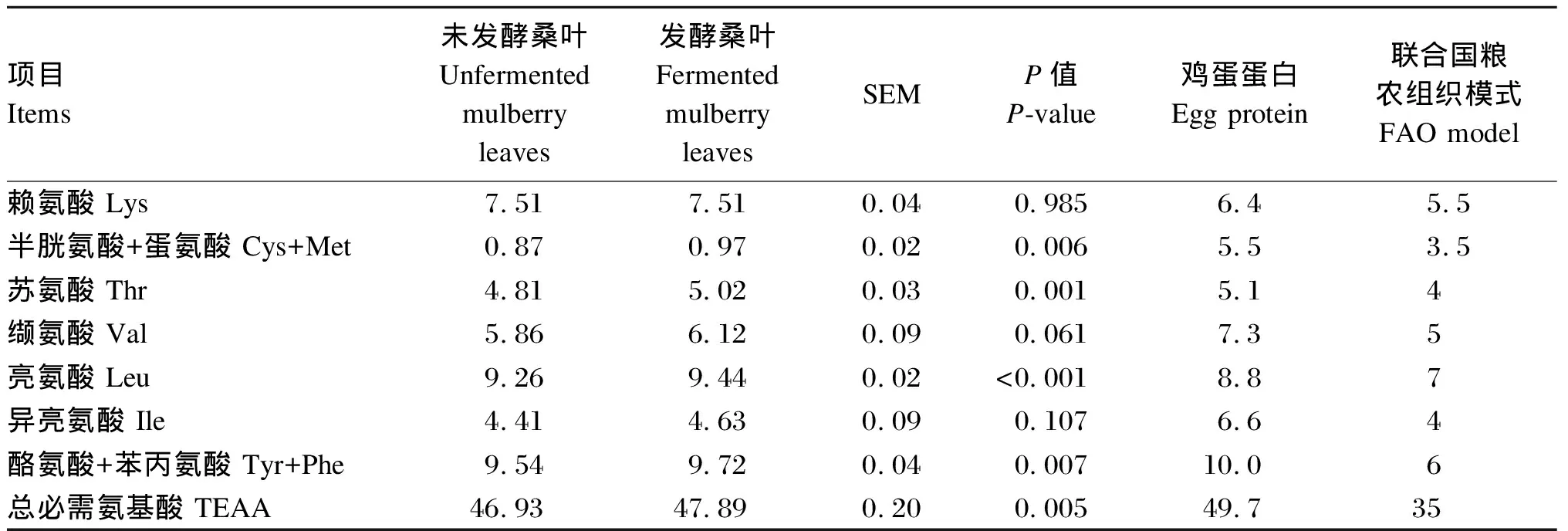
表7 未发酵桑叶和发酵桑叶必需氨基酸组成对比
菌种组合是影响发酵桑叶CP含量的主要因素。酒窖片球菌和地衣芽孢杆菌组合发酵所得的CP含量最高。酒窖片球菌、地衣芽孢杆菌与发酵基质原有的微生物产生了协同作用。混菌发酵能利用菌种之间的协同作用达到更好的发酵效果,但试验中与葡萄牙棒孢酵母有关的组合未能提高CP含量。这与微生物共生与拮抗关系有关,某种微生物生长迅速会抑制其他微生物的生长[19]。葡萄牙棒孢酵母在发酵初期的有氧状态下生长迅速,抑制酒窖片球菌或地衣芽孢杆菌的生长,在发酵后期厌氧状态下,菌体部分降解,造成CP含量未能提高。这需要测定不同时间发酵桑叶的微生物多样性才能进一步验证上述说法。
菌种组合也是影响单宁含量的主要因素。发酵技术降解单宁的实质是微生物酶(单宁酶、没食子酸脱羧酶)催化单宁降解的酶促反应[1-2,20]。酒窖片球菌+地衣芽孢杆菌、酒窖片球菌+地衣芽孢杆菌+葡萄牙棒孢酵母发酵桑叶的单宁含量显著低于酒窖片球菌+葡萄牙棒孢酵母组合。研究报道芽孢杆菌和乳酸菌降解单宁[2, 21-22],酒窖片球菌和地衣芽孢杆菌组合的微生物群落共同作用产生的单宁降解酶也许比其他菌种组合产生的降解酶效果更优、产量更大[23]。
发酵时间是影响单宁含量的主要因素,发酵4、6 d的单宁含量显著低于发酵8 d。发酵时间影响了微生物的数量,改变微生物代谢产物的含量,继而影响pH的变化。单宁酶活性通常在pH 5.5时最大[24]。地衣芽孢杆菌来源的单宁酶适宜pH为5.75[11]。不同微生物群落产生不同的单宁降解酶,这些酶在适宜pH下发挥较好的降解作用[1,20]。发酵初期微生物开始生长,pH逐步降低,发酵4、6 d期间的pH适合单宁降解酶发挥作用。另外潜在的其他变化需要更多的研究才能确定。
正交试验得出的最优条件(桑叶与麸皮质量比为9∶1,菌种组合为酒窖片球菌+地衣芽孢杆菌,接种量为2%,发酵时间为4 d)是一个整体,单独采用其中一个条件,不能达到最好的效果。发酵基质决定了能供给微生物生长的能量和营养物质,与接种量和发酵时间影响着酒窖片球菌和地衣芽孢杆菌的生长代谢。酒窖片球菌和地衣芽孢杆菌决定了产生的单宁降解酶种类[18],降解酶的产量和活性受到水分和其他代谢物含量的影响[25]。基质水分含量高,则溶氧量较低,地衣芽孢杆菌是兼性厌氧菌,在桑叶与麸皮质量比为9∶1条件下迅速生长创造厌氧环境,有利于厌氧的乳酸菌生长。2%的接种量和发酵4 d使得酒窖片球菌和地衣芽孢杆菌改变了基质的微生物群落和营养成分,使得增加的微生物和营养成分之间达到平衡,这使酶的合成和活性达到最佳值[25]。这些条件综合作用改善了桑叶的CP和单宁含量。
本发酵试验采用的是成熟绿叶,不是嫩叶和老叶。桑叶的CP含量随着叶片的发育而变化,从而导致不同的水分含量、光合作用和次生代谢产物含量[26]。嫩桑叶的EE、CP、还原性糖、总黄酮、总多酚等含量均高于老桑叶,可溶性糖含量低于老桑叶[27-28]。这些营养物质和活性物质的变化会对微生物代谢产生影响,成熟度对桑叶发酵的影响需要进行更多的研究。
3.2 发酵对桑叶pH和营养成分的影响
pH是重要的发酵饲料质量指标之一,大多数发酵饲料的最终pH均低于4.6[29]。新鲜桑叶鲜嫩多汁,口味甘甜[30],发酵后气味酸香,有助于改善适口性。发酵桑叶的pH显著低于未发酵桑叶,表明桑叶发酵良好。酒窖片球菌产生较多的有机酸,使pH快速降低,可抑制不良微生物生长。
发酵显著提高了桑叶的CP、EE、粗灰分、磷含量,降低了NDF和碳水化合物含量,与樊路杰[31]研究结果相似。樊路杰[31]研究表明,芽孢杆菌、乳酸杆菌、酵母混合发酵3 d提高了桑叶CP和EE含量,降低了桑叶DM和CF含量。上述研究的差异是桑叶来源、发酵菌种、基质配比的不同造成的。樊路杰[31]研究使用商品化菌剂发酵桑叶,本文采用酒窖片球菌和地衣芽孢杆菌发酵桑叶和麸皮。发酵后CP含量较高与乳酸菌和地衣芽孢杆菌合成菌体蛋白有关。研究报道,乳酸菌和地衣芽孢杆菌发酵能提高树叶CP含量[32-33]。另外,发酵低pH可抑制蛋白酶或形成蛋白质-多酚复合物而直接降低蛋白质水解作用[34]。此外,桑叶的功能成分可抑制微生物活性或酶促活性来促进蛋白质保存[35-36]。EE含量提高可能由于微生物利用碳水化合物合成脂肪,高灰分被认为是矿物质食物的良好来源[37]。NDF与碳水化合物含量减少,说明微生物在生长中利用碳水化合物合成了蛋白质、核酸、脂肪、有机酸等[38-39]。另一个原因是酒窖片球菌和地衣芽孢杆菌产生的高活性纤维素酶和淀粉酶能够降解纤维和碳水化合物[34]。
3.3 发酵对桑叶氨基酸含量和组成的影响
必需氨基酸的种类、含量及组成是影响饲料蛋白质品质的主要因素。除色氨酸未测定外,发酵桑叶测得17种氨基酸。发酵减少了桑叶的非必需氨基酸含量。这与以往结果一致,即发酵桑叶的氨基酸含量较未发酵桑叶减少[5]。微生物从发酵基质中摄取氨基酸进行生命活动,微生物代谢和酶促转化过程中消耗了氨基酸[9]。从CP和氨基酸含量可知,发酵提高了游离氨基酸含量。这也是发酵后部分氨基酸含量减少的原因。此外,植物蛋白酶与酒窖片球菌和地衣芽孢杆菌分泌的蛋白酶可水解桑叶蛋白质产生游离氨基酸、小肽等非蛋白氮[40]。
根据FAO/世界卫生组织(WHO)提出的理想蛋白模式[41],发酵桑叶和未发酵桑叶的TEAA/TAA(0.47~0.48)和TEAA/TNEAA(0.88~0.92)均高于理想蛋白(TEAA/TAA>0.4,TEAA/TNEAA>0.6),是优质的蛋白质来源。但发酵桑叶中含硫氨基酸组成较低,使用中需要注意氨基酸平衡。
4 结 论
发酵桑叶的最佳条件是:桑叶与麸皮质量比为9∶1,菌种组合为酒窖片球菌+地衣芽孢杆菌,接种量为2%,发酵时间为4 d。发酵提高了桑叶CP、EE、粗灰分、磷含量,降低单宁、NDF、碳水化合物、非必需氨基酸含量和pH。
猜你喜欢
英语世界(2023年6期)2023-06-30 06:29:16
今日农业(2021年21期)2021-11-26 05:07:00
今日农业(2021年13期)2021-08-14 01:38:04
——“单宁”
山东国资(2020年6期)2020-07-09 09:28:34
快乐语文(2018年36期)2018-03-12 00:55:50
阅读(高年级)(2016年4期)2016-05-30 10:48:04
应用化工(2014年11期)2014-08-16 15:59:13
环球时报(2014-07-01)2014-07-01 11:06:22
食品科学(2013年13期)2013-03-11 18:24:11
食品科学(2013年8期)2013-03-11 18:21:21
