
施肥方式对幼龄楸树非结构性碳器官分配和生长季动态的影响
2022-10-02 10:56:58关追追卢奇锋李吉跃
西北植物学报 2022年8期
关追追,卢奇锋,陈 动,邱 权,苏 艳,李吉跃,何 茜
(广东省森林植物种质创新与利用重点实验室,华南农业大学林学与风景园林学院,广州 510642)
碳水化合物是植物光合作用的主要产物,包括结构性碳水化合物(structural carbohydrate, SC)和非结构性碳水化合物(non-structural carbohydrate, NSC)。植物中能量的主要储存形式源自非结构性碳水化合物,它包括可溶性糖(蔗糖和果糖等)和淀粉,是树木生长代谢过程中重要的能源物质,可参与植物呼吸、生长以及渗透调节过程[1-2]。树木依赖并消耗积累的碳库,NSC的季节变化势必会影响源汇之间的平衡[3]。当生理代谢需求超过光合固碳量,树木体内NSC被消耗,反之,碳库得到补充[4]。叶片是植物碳同化的主要器官,叶片形态间的差异势必会影响碳吸收过程,改变NSC的季节分配模式[5]。目前,许多学者已经报道了落叶和常绿树种NSC动态变化间的差异,他们发现,与常绿树种相比,落叶树种NSC季节性波动十分明显[6-8]。在树木生长期,由于光合作用较强,落叶树种体内积累了大量NSC,而在休眠期光合强度减弱,植物呼吸消耗导致体内NSC有所降低,这引起了NSC的季节性波动[9]。只有了解了树木各器官NSC的大小和季节动态变化特征,才能对单株林木以及整个林分碳储进行量化[3,10]。目前针对灌木或草本体内NSC的研究较多[11-12],对乔木NSC的研究也多局限于某一单个器官(例如叶)[13-14],但树木器官NSC间的协同以及时间动态变化的报道较少。
作为一种珍贵落叶树种,楸树(Catalpabungei)生长迅速,树干通直,具有极高的经济和观赏价值。施肥是一项重要的营林措施,目前涉及施肥对楸树苗木生长及养分利用的研究较多[15-17],有关楸树林分碳分配对施肥响应的报道较少。据报道,水肥一体化技术具有提高水分和肥料利用率、有利于农业生产可持续发展等优点,能大幅降低肥料用量,减少养分如氮素的淋失损失[18]。目前将水肥一体化技术应用于经济作物的研究较多[19-21],关于林木的有关研究鲜有报道。基于此,笔者通过不同施肥措施(水肥一体化、穴施以及不施肥)来探究楸树林木生长、各器官NSC分配及时间动态变化对施肥的响应。本研究结果可为深入了解落叶树种器官NSC分配以及碳储变化提供理论依据。
1 材料和方法
1.1 研究地概况
研究地位于山东省济南市章丘区枣园保育库(36°25′~37°09′N, 117°10′~117°35′E)。属于温带季风气候,年均气温12.8 ℃,月均最高气温27.2 ℃(7月),最低气温-3.2 ℃(1月),无霜期192 d,年均降水量600.8 mm,年日照时数2 647.6 h。土壤类型有褐土、潮土和棕壤。
1.2 样地调查与施肥
2017年3月,选择2年生的楸树无性系“9-1”进行造林,所造纯林密度为833株·hm-2(3 m×4 m),宽行窄株距,造林面积为0.8 hm2。造林后一直未进行林分抚育。造林3个月后,测得树木平均树高为4.2 m,平均胸径为4.0 cm。所调查楸树土壤pH为7.67,有机质为19.64 g·kg-1,全氮含量为0.91 g·kg-1,全磷含量为0.53 g·kg-1,全钾含量为16.7 g·kg-1,碱解氮含量为81.88 mg·kg-1,有效磷含量为32.10 mg·kg-1,速效钾含量为176.82 mg·kg-1。
2018年5月初,随机设置9个样地开展施肥试验,每个样地面积约为400 m2。共设置3种施肥方案,分别为水肥一体化(WF)、穴施(HF)以及不施肥对照(CK)。将各施肥措施随机分配于样地中,保证每个施肥处理3次重复。样地之间均设置有隔离带,隔离带的宽度约6~8 m。于2018年5月进行首次施肥,根据前期的施肥经验,每株施肥量控制为纯N约24 g、P2O5约8 g和K2O约16 g,随后每年施肥量递增20%。穴施是在距离树干南北向50 cm的位置挖深约20~30 cm的洞穴,并将肥料一次性施入树木根系附近。水肥一体化是借助智能滴灌设备(HN-BXE, Huinong Automation Company, China)将肥料溶于水中滋润根系的方法。其方法是将全年施肥量等分为12份,从5月初开始,每10 d施肥一次,每个月施肥3次,9月初结束施肥。于2021年5月和11月调查了楸树林分生长情况(表1)。样地设置后,根据树木生长调查结果,每个样地设置5株标准木。
1.3 野外取样与NSC测定
于2021年生长季初期(5月上旬)、中期(7月中旬)和末期(9月下旬)对楸树叶、枝、粗根和细根进行采集。在标准木上,使用高枝剪剪取位于树冠中部南向的枝条,摘取3~5片位于枝末端的功能叶作为枝叶样本。距离标准木树干南向30 cm的位置,使用铁锹挖取0~30 cm深的根系,根据根系直径大小,分为粗根(>2 mm)和细根(≤2 mm)。所有样本收集后,使用纯水冲洗干净,并置于干冰中保存,立即送回实验室进行杀青(105 ℃, 30 min),随后在65 ℃条件下烘干72 h。在进行NSC分析前,将所有烘干样品研磨成细粉,过0.2 mm筛,置于封口袋中保存。
在实验室使用蒽酮-硫酸法[22]对样品NSC进行测定。NSC主要包括可溶性糖(主要是葡萄糖、果糖和蔗糖)和淀粉两部分。NSC含量用干重的百分比表示。总NSC为可溶性糖与淀粉的总和。
1.4 数据统计分析
在方差分析前,检验数据是否符合正态分布,使用Levene’s test进行方差齐性检验。当处理间方差齐性或不齐性时,分别使用Duncan和Dunnett’s T3对均值进行多重比较。使用双因素方差分析(two-way ANOVA)来探究施肥和器官(或生长时期)对NSC各组分含量的影响。使用Pearson对器官NSC进行相关性分析。使用Origin(2021 Edition, OriginLab Company, US)进行制图。
2 结果与分析
2.1 施肥方式对幼龄楸树生长的影响
各施肥处理均不同程度提高了楸树树高和胸径生长(表1)。其中,在11月份,与不施肥对照相比,穴施和水肥一体化施肥处理的树高分别提高了4.7%和7.1%,胸径分别提高了7.1%和20.5%。水肥一体化施肥的胸径显著高于穴施和不施肥对照(P<0.05)。经历一个生长季,楸树的树高、胸径在各施肥条件下均有所增加,但是增加幅度略有不同,树高增幅表现为水肥一体化(9.6%)>穴施(8.5%)>不施肥(3.7%),胸径则表现相反,为不施肥(24.5%)>穴施(23.6%)>水肥一体化(22.4%)。总体而言,水肥一体化的施肥效果要优于穴施。

表1 楸树林分生长情况调查
2.2 施肥方式对幼龄楸树NSC器官分配的影响
楸树各器官中NSC各组分含量在施肥方式下的变化不尽相同(表2)。其中,楸树叶、枝、粗根、细根的可溶性糖含量在不同施肥措施间差异均未达到显著水平(P>0.05);楸树叶、枝中淀粉含量在各施肥措施间仍无显著差异,粗根和细根中的淀粉含量在水肥一体化处理(WF)下均显著高于对照(CK),增幅分别为70.2%和18.6%,但在穴施处理(HF)下均与CK无显著差异;与淀粉含量表现类似,楸树叶、枝总NSC含量并未受到施肥方式的显著影响,但粗根中总NSC含量在WF处理下显著高于CK(22.2%),细根中总NSC含量在WF处理下显著高于HF处理,而与CK无显著差异,HF处理粗根和细根中总NSC含量均与CK无显著差异;施肥方式对楸树各器官糖淀比影响较小,叶、粗根以及细根糖淀比在不同施肥措施间差异均不显著(P>0.05),仅HF处理枝中的糖淀比显著高于CK。同时,在相同施肥措施下,各器官间总NSC及其组分含量均存在显著差异,但各器官NSC的分配特征在施肥方式间相似。其中,各施肥方式下楸树的可溶性糖含量均表现为粗根和叶片显著高于枝和细根,并以粗根最高;淀粉含量均表现为粗根和细根明显高于枝和叶,并以粗根最高,均与枝和叶差异显著;总NSC含量也表现为粗根和叶片明显高于枝和细根,并以粗根最高,均与枝和细根差异显著;糖淀比仍以叶和粗根较高。可见,水肥一体化施肥显著提高了淀粉在楸树粗根中的积累量,从而显著提高了总NSC在粗根中的积累量;各施肥方式下楸树可溶性糖、淀粉和总NSC均在粗根中分配最多,而在枝中均分配较少,施肥方式对NSC在器官中的分配没有明显影响。
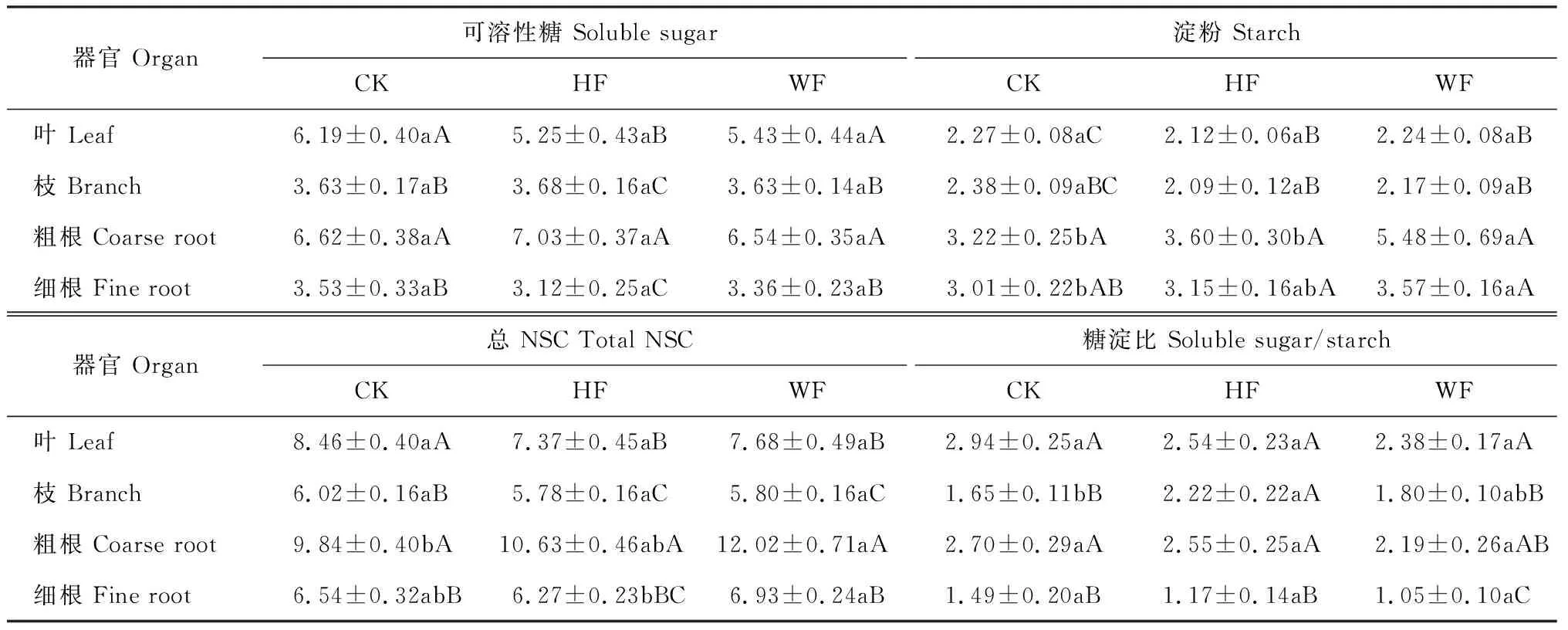
表2 楸树器官NSC分配随施肥方式的变化
2.3 施肥方式对楸树体内NSC生长季变化的影响
表3显示,施肥方式对不同生长时期楸树体内可溶性糖含量均没有显著影响(P>0.05);施肥方式对生长中期楸树体内淀粉含量也无显著影响,而对生长初期和末期淀粉含量均有显著影响,均表现为WF处理显著高于HF处理和CK,而HF处理与CK之间无显著差异,WF处理初期和末期淀粉含量分别比CK显著提高了24.6%和46.0%;施肥方式对生长初期、中期的总NSC含量影响也不显著(P>0.05),末期总NSC含量在水肥一体化施肥后比不施肥对照显著提高21.5%,而在穴施后无显著变化;穴施方式对楸树不同生长时期糖淀比均无显著影响,水肥一体化施肥楸树生长中期和末期糖淀比也均与对照无显著差异,仅导致生长初期糖淀比比对照显著降低。同时,从楸树各生长时期NSC组分含量分配情况来看,各施肥方式下不同时期的NSC组分含量分配规律基本一致。其中,可溶性糖和总NSC含量大致呈“V”型变化,即初期含量显著较高,中期含量迅速大幅降低,末期含量有所回升;淀粉含量随时间的变化呈现逐渐上升的趋势,至末期达到峰值,且各生长期之间大多存在显著差异。
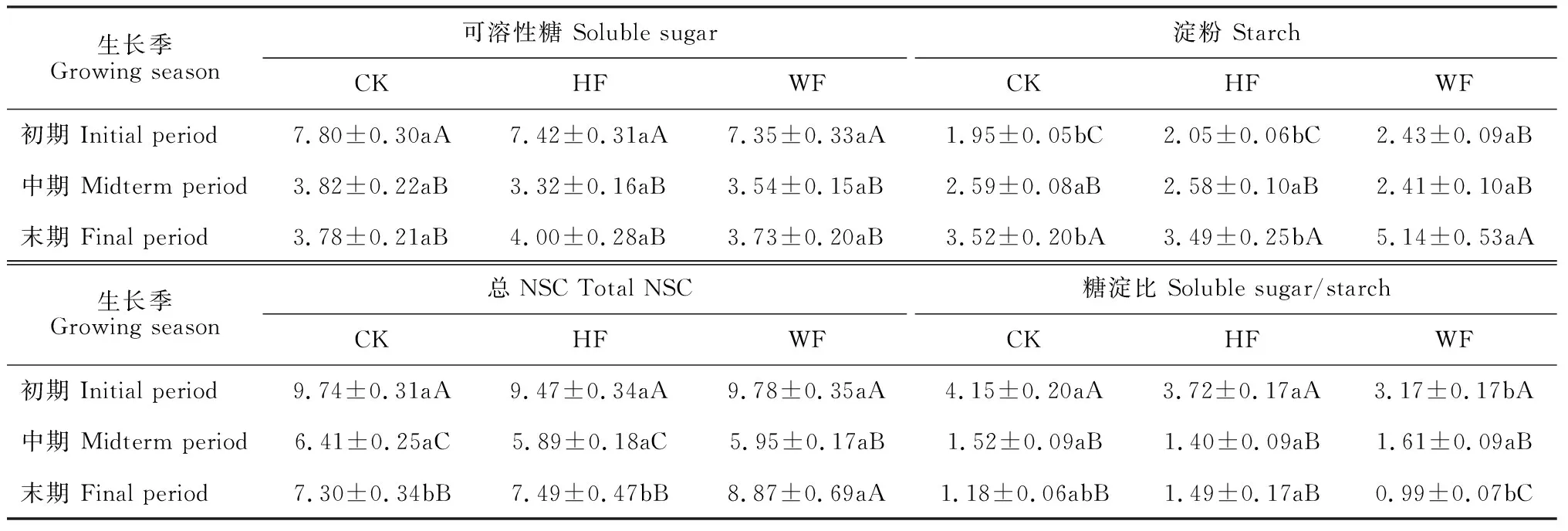
表3 楸树生长季NSC变化对施肥方式的响应
2.4 楸树器官NSC及其组分含量间的相关性
由图1可知,楸树叶可溶性糖、总NSC含量与枝、粗根和细根淀粉含量呈负相关关系;枝可溶性糖含量与粗根、细根可溶性糖含量呈正相关关系;粗根可溶性糖含量与细根可溶性糖含量呈正相关关系,但与细根淀粉含量呈负相关关系。纵观整个生长季,随着楸树各器官中可溶性糖以及叶、枝和细根中总NSC含量的消耗,提高了细根中淀粉,以及粗根中淀粉和总NSC的积累(表4)。

表4 生长季楸树器官NSC含量变化
3 讨 论
施肥可明显改变树木的生产力。本研究发现,与不施肥对照相比,穴施条件下楸树胸径提高了7.1%,尤其是水肥一体化施肥的增产效果十分明显,胸径提高了20.5%。这与秘洪雷等[23]的研究结果部分一致。他们发现,与不施肥对照相比,滴灌施肥(水肥一体化)条件下欧美杨107(Populuseuramericanacv.‘74/76’)的胸径年增量提高了19.9%~44.7%,蓄积年增量提高了18.2%~54.6%。贺曰林[24]研究发现,与空白对照相比,最佳的水肥配比下,‘毛白杨S86’(PoplartomentosaS86)人工林的胸径、树高和林地生产力分别提高了15.6%、4.9%和51.0%。究其原因可能是水肥一体化施肥增强了磷、钾等元素在土壤中的移动性,提高其利用效率,改善了土壤理化性质,提高了土壤孔隙度,进而有利于植物根系的生长发育;同时增强了叶片光合作用能力,促进了光合产物的积累,特别是增加了根系中的碳分配比例,进而提高了树木生物量和生产力[25-27]。
养分添加改变了植物的光合速率,进而可能影响其体内NSC数量及分配模式[28]。魏丽娜等[13]发现水曲柳(Fraxinusmandshurica)叶片可溶性糖的质量分数随着施氮量的增加而加大,细根淀粉和总NSC质量分数在施磷水平相同的情况下,表现出随施氮量的增加呈逐渐降低的趋势。Kong 等[29]发现施氮、磷、钾肥降低了尾巨桉(Eucalyptusurophylla×E.grandis)地下器官(主要是粗根)的可溶性糖含量,可溶性糖含量的消耗用以帮助树木抵抗干旱胁迫。Peng 等[30]发现与对照相比,施氮、磷肥显著降低了辣木(Moringaoleifera)叶片、茎和根中的NSC含量,这可能是由于施氮、磷肥后生物量增加引起的稀释效应。本研究发现,施肥虽然没有显著改变叶、枝中可溶性糖和淀粉含量,但是与不施肥对照和穴施相比,水肥一体化施肥明显提高了淀粉在根中的积累。产生不同结果的原因可能是由于树木的生活策略、施肥方式的不同引起的器官碳分配间的差异。从各器官总NSC分配情况来看,楸树叶和粗根中总NSC分配较高,枝中总NSC分配较低。一方面由于初期叶芽生长需要消耗枝中NSC引起枝NSC降低,另一方面叶通过光合作用固定了一部分碳,上一年光合产物的积累导致粗根中NSC含量升高。
不同施肥方式没有明显改变各生长时期楸树可溶性糖含量,但水肥一体化施肥明显提高了生长初期和末期淀粉和总NSC含量。楸树体内NSC含量的动态变化可能是由于树木在生长季响应环境变化(光照、温度和降水等)以及施肥而启动的碳分配策略。此外,从各生长时期NSC组分含量分配情况来看,可溶性糖和总NSC初期含量较高,中期含量迅速降低,末期含量有所回升,而淀粉含量一直增加。具体表现为,楸树生长初期叶、枝、粗根和细根中可溶性糖含量较高,至生长末期各器官可溶性糖含量一直下降,而枝、粗根和细根中淀粉含量一直增加,这与Schädel等[31]和Hoch等[8]的研究结果基本一致。一方面,消耗的可溶性糖用来帮助维持树木呼吸与生长;另一方面,可溶性糖转化为淀粉主要储存在枝和根中[31]。以上结果表明楸树把更多的碳从叶转移、供给到枝和根中,以促进根吸收水分以及更多地参与到植物的形态建成中,引起楸树叶片可溶性糖和NSC含量的降低。
本研究虽然详细探讨了楸树不同器官NSC含量随时间的动态变化情况,但也存在以下几点不足之处:其一,由于取样原因,本研究未涉及树干NSC方面的内容,树干NSC的径向和纵向变异对树木NSC分配产生较大影响[32],是今后研究的重点;其二,冠层高度对枝叶碳分配也会产生一定的影响[33-34],值得进一步研究;其三,今后应结合各器官生物量数据对林木甚至整个林分碳库进行精准评估。
4 结 论
施肥提高了楸树树高和胸径生长,尤其应用水肥一体化施肥技术增产效果十分明显。不同施肥方式提高了楸树体内总NSC含量,且水肥一体化施肥明显提高了楸树的固碳能力。楸树器官NSC含量存在明显的时间动态变化,生长初期各器官可溶性糖和总NSC含量较高;随着树木进入快速生长阶段(中期),各器官总NSC含量迅速降低;树木生长减缓或者停止时(末期),枝和根的总NSC含量逐渐升高,粗根中积累的淀粉用于维持楸树冬季呼吸以及春季枝芽生长。总之,从提高林木产量和碳储量的角度来看,将水肥一体化施肥技术应用于营林上值得优先考虑。
致谢:感谢卢奇锋、陈动对试验的帮助,感谢邱权、苏艳、李吉跃、何茜老师对本文方案设计提出的宝贵意见以及对手稿的修改,感谢鲁仪增、韩庆军和栗宁宁老师对材料收集过程中提供的帮助,感谢王军辉、麻文俊老师提供了试验材器以及试验林分,为本研究的顺利开展提供了物质保障。
猜你喜欢
热带作物学报(2022年7期)2022-08-06 02:49:32
——致秋天的花楸树
北方人(2021年19期)2021-10-29 00:46:00
农业机械学报(2020年11期)2020-11-24 13:16:54
水土保持研究(2020年2期)2020-03-19 09:03:26
农家致富顾问·下半月(2019年6期)2019-10-21 13:34:44
山东林业科技(2019年2期)2019-06-03 10:11:08
乡村科技(2019年21期)2019-02-21 09:19:29
生态学报(2018年17期)2018-10-18 02:26:48
四川农业大学学报(2018年3期)2018-09-04 08:38:36
湖北农业科学(2018年3期)2018-03-27 09:27:38
