
银屑病相关非编码RNA 及竞争性内源RNA 调控网络的研究进展
2022-08-30 02:51李睿亚刘芳芳李林烨高凯乐白羽璇
皮肤病与性病 2022年4期
何 月,段 妍,李睿亚,莎 娜 ,刘芳芳,李林烨,高凯乐,白羽璇
(1.内蒙古医科大学,内蒙古 呼和浩特 010059;2.内蒙古自治区人民医院皮肤性病科,内蒙古 呼和浩特 010017)
银屑病是一种临床上常见的慢性炎症性疾病,在西方国家银屑病的患病率约为2%,在中国则小于0.5%[1]。该疾病以难以治愈且极易复发为特点,多发于青壮年,其皮损可局部出现也可泛发至全身,同时可伴有肥胖、高脂血症、高血压、2 型糖尿病及心血管病等疾病[2]。近年来,关于银屑病的病因学与发病机制的研究均取得了一些进展,已知银屑病是由T 细胞介导、多种遗传基因参与、多种外界因素共同作用的一种难治性慢性炎症性疾病[3],具有明显的遗传易感性,尤其是在HLA-C*06:02 风险等位基因存在及具有环境触发因素如链球菌感染、吸烟、肥胖和饮酒等的情况下[4]。由于其发病机制仍不清楚,积极探索发病机制对寻找新的治疗靶点有重要的临床意义。
随着测序技术的发展,非编码RNA(Non-coding RNA,ncRNA)的功能研究逐渐走入人们的视线,包括微小RNA(microRNA,miRNA)、长非编码RNA(Long noncoding RNA,lncRNA)、 环 状RNA(circularRNA,circRNA)、信使RNA(mRNA)等[5]。想要深入了解ncRNA 的作用,就必须引入竞争性内源RNA(Competing endogenous RNA,ceRNA)的概念,ceRNA 的假说是2011 年由哈佛医学院的研究人员[6]根据实验与理论的研究提出的,即ceRNA 假说是一种新的RNA 之间的调控机制,RNA 之间可以通过与miRNA 的竞争与结合来进行交流,不同的RNA 就会形成多种调控关系,这些调控关系相互交叉就会构成ceRNA 网络。目前对ceRNA 相关致病机制的研究在多种疾病研究中逐渐增多,已证明多种ncRNA 及根据差异表达的ncRNA 构建的ceRNA 网络和多种疾病发病机制相关,进一步说明了不同RNA 分子之间互相联系及调控表达的改变可能与银屑病发病有关。本文针对ncRNA 及ceRNA 调控网络在银屑病发病机制中作用研究的进展进行概述,为推进银屑病相关ncRNA 的发病机制的研究提供新的思路与方向。
1 非编码RNA 与银屑病
对银屑病非编码RNA 调控的研究主要集中于miRNA,近年来对lncRNA 及circRNA 的研究也不断兴起。
1.1 银屑病相关异常表达的miRNA
研究发现一些miRNA 在银屑病的发病中通过调节皮肤炎症、增殖及形态发生过程发挥重要作用,随着对miRNA 的深入研究,发现越来越多的非编码miRNA 在银屑病的病理条件下发挥作用[7-8]。在银屑病中异常表达的miRNA 存在于银屑病患者外周血单核细胞及血浆中,或皮损中,或两者中都存在,主要有 miR-125b、31、203、miR-2l、miR-146a/b 等,其 他miRNA 包 括miR-99a、miR-210、miR-193b、miR-223、miR-122-5p、miR-548a-3p、miR-20a-3p、miR-136、miR-138、miR-155 等也有报道[9-11]。目前研究发现250 多个miRNA 在银屑病患者的皮损或血液中异常表达且作用的靶点及信号通路不尽相同[12],现将miRNA 影响角质形成细胞、免疫调节等不同作用通路导致银屑病的发病机制汇总如下。
1.1.1 miRNA 对角质形成细胞的作用 多年来,角质形成细胞被发现在免疫皮肤反应的启动、维持和调节中起着至关重要的作用。它们作为反应炎症介质(包括白介素-17、36)的执行者,发展成全面的银屑病表型。角质形成细胞可以产生多种趋化因子、细胞因子、抗菌肽等,这些细胞因子和抗菌肽通过参与表皮增生来扩大局部炎症反应和维持炎症级联反应,而miRNA 可对角质形成细胞进行调控,例如引起细胞异常增殖及分化等[13]。
据报道,miRNA 可以靶向调控1/3 以上的人类基因,即在许多细胞活动(例如细胞生长、分化、发育和凋亡)中发挥调节作用,越来越多的研究支持miRNA 参与皮肤及其附属器官的形态发生和动态平衡,miRNA 在包括角质形成细胞在内的皮肤细胞中起着分化、增殖和激活等作用,这可表明miRNA 在表皮屏障完整性中起着重要作用。例如,细胞外钙、p63 和Notch 信号可上调或下调角质形成细胞分化和增殖过程中的miRNA,miRNA 还可以直接靶向作用于p63 和Notch。大量研究表明,miR-31 通过调节NF-κB、Ras/MAPK、Notch 信号通路等多种细胞因子,对角质形成细胞的增殖、分化和细胞活性具有正向调节作用。目前,角质形成细胞中miR-31 与NF-κB信号通路的相互作用是研究的热点,角质形成细胞的异常状态可能是多种疾病的突破口,而miR-31 和NF-κB 信号形成的正反馈环可能为银屑病的预防带来新的思路,miR-31 通过正向调节角质形成细胞促进创面愈合的能力在慢性溃疡和创伤的治疗中也值得进一步研究[14]。在角质形成细胞异常增殖的皮肤疾病中,角质形成细胞在完整的皮肤屏障中的增殖和分化之间的平衡是负的,类似的miRNA 是否参与角质形成细胞的分化和增殖可能取决于相应miRNA 的上调或下调。miR-203 是第一个被发现,也是在表皮分化中表达最高的miRNA,在miR-125b、miR-146a和miR-181a 中,类似的miRNA 上调和下调对角质形成细胞分化和增殖的影响也得到了证实,但是关于miRNA 与细胞间黏附或皮肤脂质之间联系的证据较缺乏[15]。
2021 年Soonthornchai W 等[16]在活动性银屑病皮损活检中发现miR-378 a-3p 会显著上调,而在甲氨蝶呤或窄带紫外线光疗治疗后会下调。该研究还使用角质形成细胞体外模型进一步证明miR-378 a会干扰细胞周期进程,导致G1 期细胞周期停滞。对miR-378 a 过表达和缺失的角质形成细胞进行相关的转录组分析揭示了与炎症紧密连接相关的重要生物学机制,通过荧光素酶分析评估的靶基因转录物还进一步确定了骨形成蛋白2(Bone morphogenetic protein 2,BMP-2)是miR-378 a 的新靶基因。2021年Huang C 等[17]证实了miR-193B-3p 在银屑病患者、银屑病样炎性细胞模型和咪喹莫特诱导的小鼠模型中的下调。
1.1.2 miRNA 在银屑病中的免疫调节作用 目前,银屑病与免疫系统异常调节是研究的热点,银屑病的发生和发展与T 细胞为主的免疫细胞密切相关,即免疫介导的炎症介质如肿瘤坏死因子-α、干扰素-γ、白细胞介素-1、17、22 的分泌会进一步导致银屑病的炎症表现[3,18]。
miR-203 被证实是可以调节银屑病细胞因子如肿瘤坏死因子-α、白细胞介素-24 和白细胞介素-8的作用,该研究表明miR-31 通过STK40 靶向促进NF-κB,并 导 致CXCL1、CXCL8、CXCL5 和 白 介素-1β 的分泌,这些物质促进血管内皮细胞活化并通过趋化性将白细胞吸引到皮肤中,还有一些研究表明银屑病患者皮肤和外周血单个核细胞中高水平的miR-146a 分别与皮肤和血清中的白介素-17 水平呈正相关,miR-155-5p 已显示在血液和银屑病皮损中上调[18]。2020 年的一些研究中[19]发现miR-146a在免疫系统动态平衡中起着重要作用,它的异常表达或功能导致了各种免疫性和非免疫性炎症性疾病的发生和发展。这种miRNA 是典型的炎症负性调节因子,并与NF-κB 途径相互作用,可参与几个过程(如基因多态性、miRNA 编辑等),进而造成不同疾病的性质、表达和调节功能都会发生不同程度的变化。在银屑病患者的皮损、全血和血浆中miR-146a的表达显著升高,并与白介素-17 的表达、银屑病严重程度指数、面积呈正相关。2020 年Liu T 等[20]发现miR-617 在银屑病患者皮肤组织和白介素-22刺激的角质形成细胞中均升高,miR-617 可通过靶向FOXO4 促进白介素-22 刺激的角质形成细胞生长。2021 年Srivastava A 等[21]通过对银屑病中干扰素-γ的作用进行研究,发现干扰素-γ 激活角质形成细胞通过抑制miR-149 诱导炎症反应,可能具有治疗银屑病的意义。Meng J 等[22]通过探讨miR-15a-5p 对角质形成细胞增殖和凋亡的影响以及潜在的分子机制发现银屑病患者miR-15a-5p 表达显著下调,miR-15a-5p 在体外银屑病模型中使用白介素-22 诱导的细胞对角质形成细胞具有抗增殖和促凋亡作用,并通过体外银屑病细胞培养模型揭示其可以通过靶向FOSL1 调节MAPK/ERK 信号通路。有研究发现miR-21-3p 在银屑病样小鼠模型中的上调与白介素-22在体外和体内的表达和功能有关,并通过STAT3 和NF-κB 信号发生,这项研究强调了miR-21-3p 在皮肤系统中的多效性功能,从调节细胞增殖和存活到潜在地调节细胞先天性免疫和获得性免疫[23]。在一些研究中,银屑病患者血清中miR-221-3P 水平明显升高,血清miR-221-3p 与肿瘤坏死因子-α、白细胞介素-17A、22 的表达水平呈正相关,miR-2213p 的下调也抑制了角质形成细胞中某些炎性因子的释放,进而抑制角质形成细胞的增殖和炎症反应,为银屑病的治疗干预提供了可能的靶点[24]。
1.1.3 miRNA 在银屑病中的其他作用 miRNA 还可以作为潜在的生物标志物,用来诊断银屑病、反映银屑病的活动性、监测治疗效果、作为相关的合并症的生物标志物、预测银屑病性关节炎的发生等[25],例如2020 年Domingo S[18]等发现银屑病患者外周血单个核细胞(Peripheral blood mononuclear cell,PBMC)中miR-223 和miR-143 表达明显上调,提示miR-223 和miR-143 可能成为银屑病新的诊断生物标志物。与正常对照组相比,只有银屑病患者发现miR-19a 水平显著上调,结果支持将其作为银屑病的非侵入性诊断标记物;目前的数据表明PBMC 中miR-99a 与疾病严重程度呈负相关。最后检测血清和皮肤miR-369-3p 水平,并证实其与疾病严重程度的相关性,其中皮肤miR-369-3p 水平与PASI 评分呈线性正相关。相反,皮肤中miR-369-3p 和miR-135B 水平与疾病改善和严重程度正相关。
2019 年Duan Y 等[26]研究发现血浆miR-126 的表达与银屑病的危险性和严重程度呈负相关,其高基线水平可作为预测雷公藤红素联合阿维A 治疗银屑病临床疗效较差的生物标志物。银屑病中发现的失调miRNA 已被证明会影响角质形成细胞和T 细胞的功能,一些研究发现上调的miR-31/miR-203/miR-155/ miR-21 和下调的miR-99a/miR-125b 可以促进银屑病角质形成细胞的过度增殖和异常分化;上调的miR-210 和下调的miR-138 协同作用,使银屑病患者的CD4+T 细胞亚群失去平衡。毫无疑问,对于参与银屑病的miRNA 相关调节机制的进一步研究有助于发现治疗这种难治性疾病的新策略[27]。
1.2 银屑病相关异常表达的lncRNA长链非编码RNA(Long non-coding RNA,lncRNA)为非蛋白质编码RNA 转录本,长度超过200 个核苷酸,是多种细胞过程中的关键调控因子,还参与表观遗传沉默、剪接过程、翻译、凋亡和细胞周期调控等。此外,多种lncRNA 的表达水平与表皮分化和免疫调节密切相关,有很多例子表明lncRNA 也参与调节多种皮肤病理状况,包括皮肤癌、伤口愈合、银屑病等[28]。
1.2.1 lncRNA 在角质形成细胞中的作用 抗分化非 编 码RNA(Antidifferentiation non-coding RNA,ANCR)在表皮分化中起负调控作用,祖细胞中ANCR 的丢失快速诱导分化程序,因此可以抑制表皮基底层的过早分化,降低角质形成细胞对自然凋亡的敏感性,改变正常角质细胞的功能[18]。组织分化诱导非蛋白编码RNA (Tissue differentiation inducing non-protein coding RNA ,TINCR)在表皮组织中的主要功能是诱导分化,在皮肤病,尤其是以表皮屏障破坏为特征的病变中,它异常表达[29]。一项研究表明长链非编码RNA-PRINS 在银屑病患者的非皮损区升高,而在银屑病斑块中降低,PRINS 可以通过下调G1P3 参与银屑病的发生[18]。H19 直接与miR-130b-3p 相互作用,增强下游靶基因Dsg1 的表达促进角质形成细胞的分化[30]。银屑病中,lncRNA H19 的下调通过上调miR-766-3p 的表达水平,通过AKT/mTOR 途径抑制S1PR3 的激活,即下调lncRNA H19 可以促进角质形成细胞增殖和皮肤炎症[31]。lncRNA MEG3 可以通过调节miR-21 表达可以调控caspase-8 的表达,进而影响银屑病角质形成细胞、Act-HaCaT 和Act-HHEK 细胞的增殖和凋亡[32]。lncRNA MEG3 还可以在TNF-α 处理的角质形成细胞和银屑病小鼠中促进自噬、抑制炎症反应且依赖于PI3K/AKT/mTOR 信号通路[33]。
1.2.2 lncRNA 在银屑病中免疫调节作用 2018 年,Meng Q 等[34]的实验结果表明,lncRNA-MSX2P1 可通过抑制miR-6731-5p、激活S100A7 进一步促进白介素-22 的释放,刺激角质形成细胞的生长。2021年研究[35]检测到银屑病患者病变中 MALAT-1表达的显著增加,与对照组相比,患者的血清 miRNA-9 显著增加,银屑病患者皮损皮肤中MALAT-1 的表达升高可能有助于银屑病斑块的发展,MALAT-1 和miRNA-9 的高血清表达表明这些循环标志物可以作为银屑病诊断的潜在标志物。一些研究为了发掘lncRNA MALAT-1 对角质形成细胞的影响,通过检测银屑病皮损、正常皮肤组织及白介素-22 刺激的角质形成细胞中的MALAT-1和S100A7 水平,监测细胞增殖、炎症和凋亡,发现MALAT-1 和S100A7 水平均上调,MALAT-1 还可以与S100A7 竞争以阻止mir -330-5p 诱导的S100A7 表达抑制,miR-330-5p 与MALAT-1(或S100A7)在银屑病病变组织中的表达呈负相关。在白介素-22治疗的反应中,miR-330-5p 沉默消除了角质形成细胞中MALAT-1 敲低的影响,表明MALAT-1 可以通过miR-330-5p/S100A7 轴调节白介素 -22 诱导的角质形成细胞的变化[36]。
lncRNA NEAT1 表达量和银屑病疾病活动度呈正相关,还可以直接调控炎症反应相关通路,TLR4 介导的炎症反应过程是通过直接调节炎症反应相关通路来促进的,例如诱导MAPK 通路激活,从而诱导促炎因子的表达[37]。一些研究还发现芍药苷可以通过NEAT1/miR-3194-5p/Galectin-7 轴控制银屑病角质形成细胞的增殖和迁移[38]。有研究发现银屑病患者皮肤组织中AGAP2-AS1 水平较健康对照组高,且AGAP2-AS1 可促进角质形成细胞增殖并抑制凋亡,METTL3 的下调还会导致银屑病中 AGAP2-AS1 的上调,AGAP2-AS1 还可以通过miR-424-5p 上调 AKT3、激活 AKT/mTOR 通路以及促进角质形成细胞的增殖[39]。lncRNA AGAP2-AS1 还可以抑制miR-6731-5p,间接促进S100A7的表达,进而促进炎症因子的分泌,从而促进角质形成细胞增殖并抑制凋亡[34]。
2019 年Yan J 等[40]研究发现1549 个异常调控的lncRNA 并预测了靶基因。发现lncRNA 在调节免疫系统的各种功能中发挥着重要作用,越来越多的证据表明lncRNA 失调可能在银屑病的发病机制中起着重要作用。银屑病病变皮肤组织中lncRNA-ANRIL水平与TNF-α、白介素-6、17 水平正相关,表明lncRNA-ANRIL 可能是通过促进患者的炎症来加重患者的疾病严重程度,它还可以通过调控多种通路,如诱导NF-κB 途径激活来促进NF-κB 下游的促炎因子TNF-α、白介素-6、17 的表达[41]。
1.3 银屑病中异常表达的circRNA circRNA 也是一种非编码RNA,几乎可以存在于所有生物细胞中,具有更高的相对稳定性及保守性[42]。随着对ncRNA 逐步了解,部分银屑病相关circRNA 的研究也逐渐增多。2018 年Qiao M 等[43]采用银屑病患者与健康者的皮肤组织提取RNA 并筛选验证出差异表达的circRNA,发现HSA_CIRC_0061012 在银屑病皮损中显著上调,提示其可能是银屑病的候选生物标志物。2021 年He Q等[44]发现CIRC_0061012 可以通过靶向miR-194-5p/GAB1 轴促进白介素-22 诱导的HaCaT 细胞的增殖和运动,为银屑病患者开发新的治疗靶点提供了新的思路。2020 年Moldovan LI 等[45]发现银屑病和特应性皮炎的环状RNA 转录组具有共同的表达特征,包括相对于健康皮肤的整体下调,但这在银屑病中最为明显,而且只有银屑病的几个环状RNA 不依赖于它们的同源线性转录本而处于失调状态,所以特异性环状RNA 可能被用于区分特应性皮炎与银屑病。2019年Liu R 等[46]首次报道银屑病患者血浆中hSA_CIRC_0003689、CHR4:121675708|121732604 和hSA_CIRC_0003718 在银屑病患者血浆中大量表达,这与在骨髓间充质干细胞中观察到的结果一致,提示银屑病患者中广泛存在着异常的circRNA,还发现银屑病皮损来源中的circRNA-ChR2:206992521|206994966通过影响T 淋巴细胞细胞因子的分泌,改变其活性,进而影响银屑病的发病机制[47]。
2 ceRNA 网络与银屑病
竞争性内源RNA(Competing endogenous RNA,ceRNA) 是一类能与miRNA 结合位点结合,从而能进一步增加相应mRNA 的表达来达到RNA 间相互沟通及转录后调控基因表达的一类非编码RNA。例如当细胞内lncRNA 表达量下降时,更多的miRNA 与mRNA 结合导致mRNA 的表达量下降,因此不同的lncRNA、不同的circRNA 与miRNA 就可以相互影响、相互竞争(图1)。ceRNA 机制还被发现广泛存在于各类肿瘤疾病中,并在肿瘤基因调控等生物过程中发挥作用[5],作为生物标志物、潜在的治疗靶点及强有力的预后指标,构建ceRNA 调控网络对于进一步了解ncRNA 的功能具有重大意义。
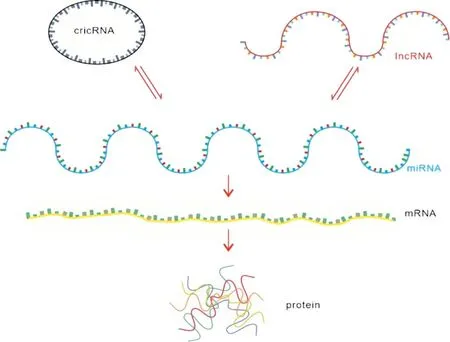
图1:ceRNA 网络作用机制
本文将结合ceRNA 网络作用的机制和分子基础,阐述银屑病相关ceRNA 即lncRNA 和circRNA 的最新研究进展,进一步探讨和总结该机制在银屑病发病机制中的作用,进一步拓宽ncRNA 在银屑病发生发展机制中的视野,银屑病ceRNA 网络的最新研究结果总结如下:
2.1 构建lncRNA-miRNA-mRNA 网络以确定lncRNA在银屑病中的调节作用 2019 年Zhou Q 等[48]筛选出差异表达的lncRNA、miRNA 和mRNA,并根据ceRNA 理论构建了银屑病相关的lncRNA-miRNAmRNA 网络,结果发现其中共有253 个lncRNA、106mRNA 和1156mRNA 在银屑病患者皮肤和健康对照皮肤中差异表达。最后该研究还发现了两个在银屑病的发病机制中可能起重要作用的关键lncRNA:AL035425.3 和PW AR6,而且PW AR6 可能通过调控miR-155-5p 和miR-369-3p 的表达参与银屑病的发生。
2020 年Li H 等[49]采用加权基因共表达网络分析的方法,对银屑病患者和正常皮肤组织的RNA序列进行分析,得到与银屑病相关关键mRNA 和lncRNA,确定银屑病相关基因模块。共鉴定出11 个重叠的关键mRNA,包括2 个已知基因(KRT15 和CCL27)和9 个新基因(ARSF、CLDN1、DACH1、LONRF1、PAMR1、RORC、SLC26A2、STS、UNC93A),共筛选出11 个关键mRNA 构建共表达网络,以研究潜在的候选lncRNA,还发现76 对lncRNA-mRNA 共表达关系,结果发现银屑病皮损中CCL27 和lncRNA-AL162231.4 表达降低。
2.2 构建circRNA-miRNA-mRNA 网络以确定circRNA 在银屑病中的调节作用 2019 年Liu R 等[46]采用RNA 测序技术检测银屑病皮损和正常皮肤的间充质干细胞中circRNA 的表达,并预测了circRNAmiRNA 相互作用网络。结果共检测到6323 个circRNA,其中3227 个是之前未报道过的circRNA。通过鉴定出129 个在银屑病和对照组之间表现出显著不同的circRNA 表达模式,并通过生物信息学分析建立了一个circRNA-miRNA 相互作用网络,进一步表明miR-17-5p、miR-30e-5p、miR-142-3p/5p、miR-3693p、miR-184、miR-654-3p 和miR-423-5p与circRNA 相互作用。而且,miR-17-5p、miR-30e-5p 和miR-142-3p/5p 在银屑病患者的皮损中均有不同程度的表达,其中miR-142-3p 特别涉及到表皮炎症。根据circRNA-miRNA-mRNA 相互作用网络,与这些mRNA 相关的circRNA 的表达在银屑病皮肤病变中也被下调,根据ceRNA 调控网络机制,circRNA可以通过竞争miRNA 位点进而减轻miRNA 对靶基因的抑制,由此可推测银屑病中circRNA 的表达下调可能与其靶mRNA 的表达下调有关[47]。
3 展望
随着研究技术的不断更新,对ncRNA 种类及其功能的研究逐渐受到越来越多的重视,而银屑病的发病机制研究一直是皮肤科医生和研究人员的难题,本篇文章通过对3 种主要非编码RNA 对银屑病发病机制的作用及影响进行综述,进一步总结了ncRNA在银屑病中的研究进展,为后续银屑病相关的病因、治疗及预后等研究提供新的思路。
猜你喜欢
世界中医药(2022年22期)2022-12-14
中国典型病例大全(2022年13期)2022-05-10
疑难病杂志(2021年10期)2021-10-21
皮肤病与性病(2021年3期)2021-07-30
健康之家(2021年19期)2021-05-23
儿童故事画报·智力大王(2020年1期)2020-04-28
凤凰生活(2018年6期)2018-06-25
健康之家(2016年10期)2016-10-28
中国实用医药(2016年1期)2016-01-11
母子健康(2015年8期)2015-12-13
