
糖皮质激素通过nr3c1受体依赖和非依赖途径介导骨质疏松症的机制
2022-08-30 05:58袁涛姜宇
中国骨质疏松杂志 2022年8期
袁涛 姜宇
1.南京医科大学附属无锡第二人民医院骨科,江苏 无锡 214002
2.南京医科大学附属无锡第二人民医院全科医学科,江苏 无锡 214002
糖皮质激素(glucocorticoids,GCs)类药物通常用于治疗多种疾病,包括炎症和自身免疫性疾病[1];然而长期使用GCs会引起骨组织损伤,最终导致糖皮质激素性骨质疏松症(glucocorticoid-induced osteoporosis,GIOP)[2]。骨质疏松症是一种全身性骨病,其特征是骨密度降低,骨组织微结构破坏,导致骨脆性增加、骨折风险增加[3]。GIOP是最常见的由药物引起的骨质疏松症类型,也是第二大常见的骨质疏松症类型[4]。强的松龙(PN)是一种糖皮质激素类药物,具有强大的抗炎和免疫抑制特性[5]。文献报道,长期使用PN会导致骨密度下降和骨丢失,从而导致骨质疏松症[6]。在细胞内,GCs及其类似物通过GC受体(GR,也称为nr3c1)调节下游基因的表达。
斑马鱼是一种优秀的模型生物,并已成功应用于各种生物医学研究[7]。本研究利用CRISPR/Cas9技术成功地建立了斑马鱼nr3c1-突变体,用于研究斑马鱼的骨矿化和骨代谢相关基因的表达,同时评价了nr3c1在骨代谢相关基因表达调控中的作用。
1 材料与方法
1.1 斑马鱼养殖与培养
成年AB品系斑马鱼在28 ℃、14 h/10 h的光/暗周期下,在循环水系统中饲养,每天投喂3次。为了获得胚胎,将雄性和雌性斑马鱼在晚上配对,第2天在开灯后1 h内雌性斑马鱼产卵。将胚胎放入10 cm厚的培养皿中,并用亚甲基蓝(0.3 ppm)加鸡蛋水,在28 ℃的光控(14 h/10 h明/暗)培养箱中培养。
1.2 nr3c1突变斑马鱼的选育
本研究利用CRISPR-Cas9技术建立nr3c1-突变体。本研究在第2外显子设计靶位点,并在体外合成nr3c1 gRNA,然后将nr3c1 gRNA(100、50和25 pg)和Cas9-Capped-mRNA(300 pg)在单细胞阶段共注射到斑马鱼胚胎中。然后将注射的胚胎保存在28.5 ℃的E3培养基(5 mmol/L NaCl、0.17 mmol/L KCl、0.33 mmol/L CaCl2和0.33 mmol/L MgSO4)中,并在2 DPF下提取3组(每组10个胚胎)的gDNA,然后扩增出含有nr3c1靶位点的243bp DNA片段,用T7内切酶I(New England Biolabs)将7 μL扩增产物在37 ℃下酶切3 h。用美国国立卫生研究院的ImageJ定量分析裂解带和未裂解带的强度,并估计突变频率。将PCR产物克隆至pMD-19 T(Takara Bio Inc.)用于测序分析。为了鉴定胚系传递的突变,将显微注射的鱼胚胎(F0)培育到成年,然后将F0鱼与WT斑马鱼进行异型杂交,产生F1胚胎。然后从每个杂交组合中收集10个F1胚胎进行gDNA提取和酶消化。携带可遗传突变的F1胚胎的同胞被培育到成年,并通过PCR扩增和剪鳍DNA测序重新鉴定了每一条F1鱼。
1.3 阿尔新蓝染色及全标本包埋骨骼染色
为了观察受精后96 h胚胎前骨骼的发育变化,用立体显微镜获取图像(Leica,Wetzlar,Germany),并用阿尔新蓝染色,重复3次。用Image J测量颅间距离(ICD)、下颌骨长度(LJL)和颈椎软骨长度(CCL)。
斑马鱼幼体的茜素红染色实验如前所述[8]。用立体显微镜(Leica)采集图像。本研究进行了数字图像分析,以量化染色区域的表面和密度。
1.4 斑马鱼样品RNA提取、cDNA合成及qRT-PCR
用Trizol试剂(Invitrogen,Carlsbad,CA,USA)提取30多条幼体的总RNA,用SuperScriptTMⅢ逆转录酶(Invitrogen)逆转录成cDNA。反应条件为95 ℃、10 s和60 ℃、30 s,进行40个循环。以看家基因β-肌动蛋白为对照,所有结果均标准化为该基因的表达水平。
1.5 蛋白质印迹分析
对来自WT和nr3c1-/-样本的约100条幼体进行了蛋白质印迹分析。
1.6 荧光素酶报告测定
将mmp9、alp和acp5a启动子克隆到pGL4.17载体中,构建mmp9、alp和acp5a-Luc载体,并与nr3c1表达载体共转染HEK 293 T细胞进行双荧光报告实验。
1.7 统计学分析
数据以平均值±SD表示。两组或更多组之间的统计差异分别使用t检验和单因素方差分析,然后进行Student-Newman-Keuls检验。所有实验至少重复3次。P<0.05表示差异有统计学意义。
2 结果
2.1 nr3c1缺失突变斑马鱼的选育
在斑马鱼中,糖皮质激素受体由nr3c1基因编码,该基因包含9个外显子。起始密码子ATG位于第二外显子,编码蛋白由746个氨基酸组成。在与ATG位点相邻的第二个外显子上预测并选择了该基因上的gRNA靶位点(图1A)。诱变效率实验结果表明,注射100、50和25 pg的gRNA组的效率分别为90%、87%和88%(图1B)。将PCR片段克隆到测序载体PMD-19 T后,单克隆测序显示F0中nr3c1 gRNA靶点有4种不同类型的插入突变(图1C)。因此,这些显微注射的F0胚胎的同胞被培育到成年,并与WT鱼杂交产生F1胚胎。获得了两个可遗传突变,即5-bp缺失和7-bp缺失,并与含有相同突变的F1鱼杂交,获得纯合子nr3c1突变系(图1 D、1E)。Western blotting结果表明,在nr3c1-突变体中未检测到NR3C1蛋白(图1F)。这些结果表明该nr3c1-突变体是一个无义突变体。
2.2 nr3c1-突变体的软骨发育异常
对4 DPF、5 DPF和6 DPF斑马鱼幼体的软骨进行阿尔新蓝染色,结果表明,4 DPF时,PN组、nr3c1突变组和nr3c1突变PN处理组与WT组相比,头面部软骨发育明显减慢(图2A~2 D),尤其是nr3c1突变PN处理组,难以观察到Meckels软骨(图2 D)。5 DPF时,将主要软骨结构染色,PN组和nr3c1突变PN组与WT组比较,前侧筛板、外侧缘McGills软骨、舌内侧软骨和后角形态无明显差异(图2E~2 H)。然而,nr3c1突变组体现出角质支气管部和角质膜形成的延迟(图2 G)。6 DPF时,各组颅面软骨清晰可见,软骨形态无明显变化(图2I~2 L)。为了进一步评估软骨的发育变化,测量了6 DPF斑马鱼幼体的软骨距离,即颅内距离(ICD)、下颌长度(LJL)和角软骨长度(CCL)(图3A)。PN处理没有影响软骨与对照组之间的距离(图3B~3 D);相反,在nr3c1突变体中,软骨发育距离显著缩短,表明nr3c1突变导致了头面部骨骼变小(图3B~3 D)。此外,用PN处理nr3c1-突变体后,3个参数没有显著差异(图3B~3 D)。为了探讨颅面骨缺损背后的原因,在6 DPF中计算了颈椎软骨中的软骨细胞数(图4A~4 D)。结果表明,nr3c1突变体的软骨细胞数量显著减少(图4E)。这些结果表明,nr3c1的缺失影响软骨的发育,而PN影响软骨发育的具体机制尚需进一步研究。
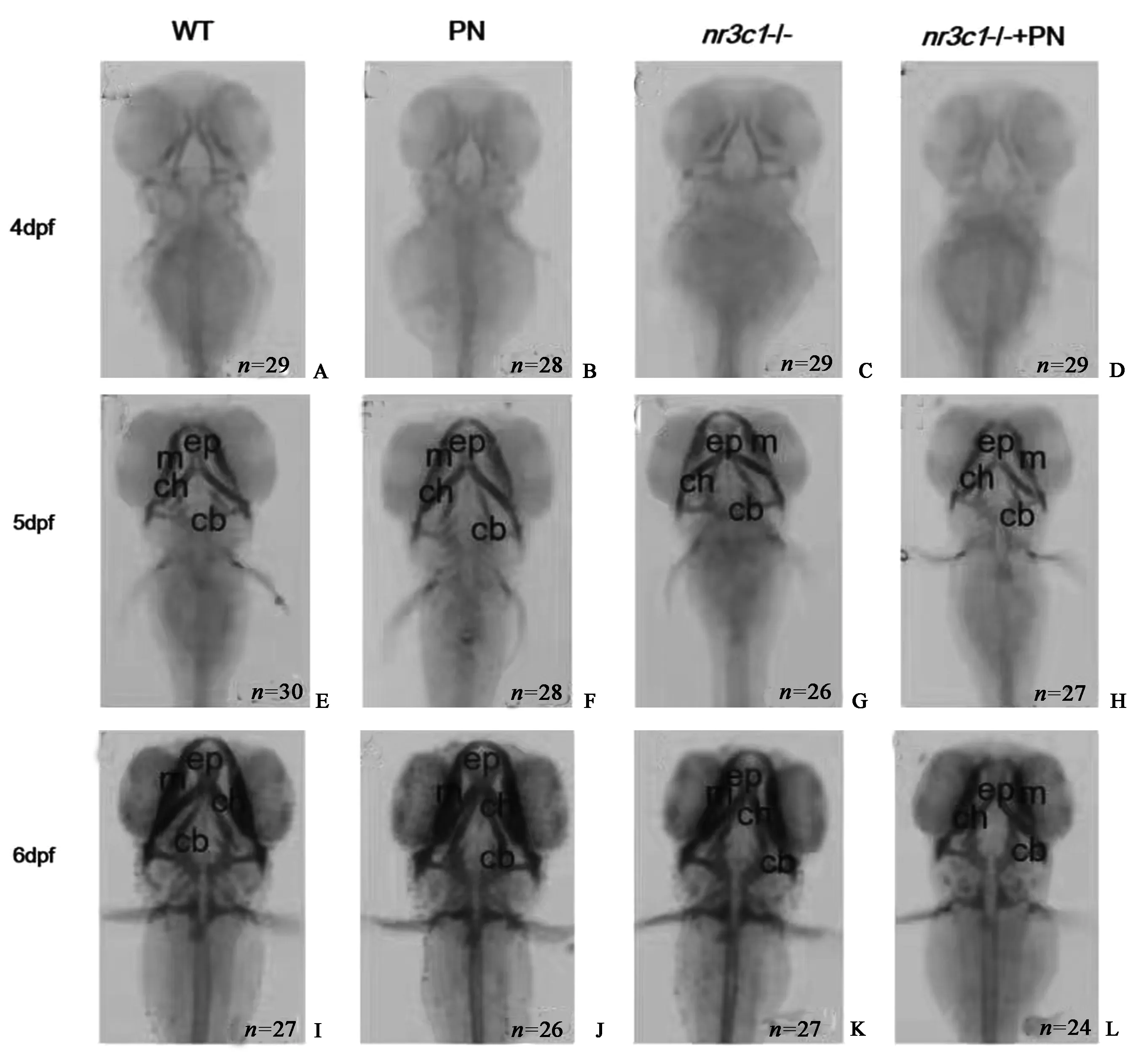
图2 nr3c1-突变体的软骨异常 WT组(A、E、I组)、25 μmol/L PN处理组(B、F、J组)、nr3c1-/-突变组(C、G、K组)和25 μmol/L PN处理的nr3c1-/-突变组(D、H、L组)受精后4~6 d进行软骨阿尔辛蓝染色。
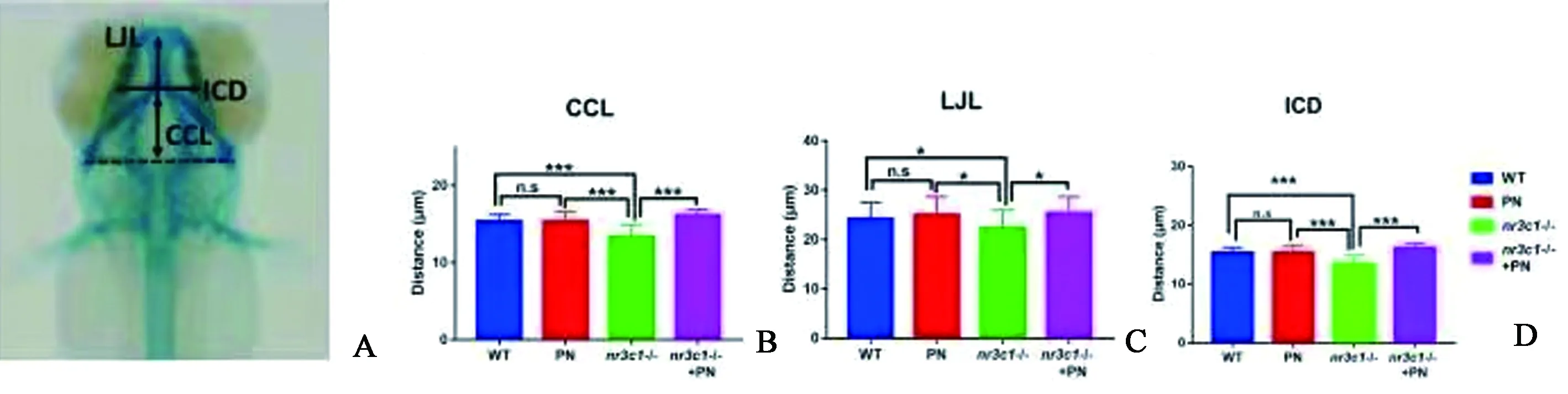
图3 nr3c1-突变体软骨长度异常

图4 nr3c1的突变减少了软骨细胞的数量 WT组(A)、PN处理组(B)、nr3c1-/-组(C)和PN处理的nr3c1-/-组(D)显示角软骨发育情况;E为计算软骨细胞的数量。
2.3 nr3c1-突变体骨矿化减少
取7 DPF、8 DPF、9 DPF的幼鱼进行茜素红染色(图5A),结果显示,在7 DPF,与WT相比,PN处理导致骨矿化显著减少(图5B),nr3c1突变组显示骨矿化显著减少(图5C),nr3c1突变与PN处理导致更严重的骨矿化减少(图5 D)。在8 DPF和9 DPF,PN处理和nr3c1突变后的骨矿化表型更为明显(图5E~5 L)。用Image J测量矿化面积,结果表明,在PN处理后和nr3c1-突变体中,矿化面积在7 DPF(图6A)、8 DPF(图6B)和9 DPF(图6C)处与对照组相比显著减少。这些结果表明,nr3c1的缺失和PN处理均会影响斑马鱼幼体的骨矿化。

图5 nr3c1-突变体组和PN处理的nr3c1-突变体组的骨矿化 受精后7~9 d的斑马鱼幼体用茜素红在(A、E和I) WT、(B、F和J) PN处理组、(C、G和K) nr3c1-突变体组和(D、H和L) PN处理的nr3c1-突变体组中染色。
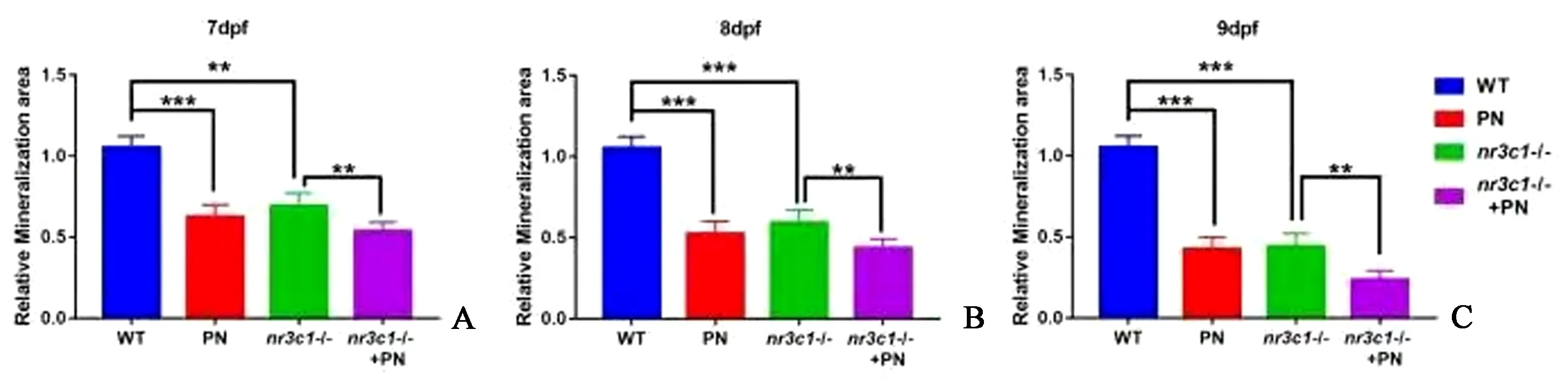
图6 nr3c1-突变体组和PN处理的nr3c1-突变体组中骨矿化的定量分析
2.4 nr3c1-突变体骨代谢相关基因表达模式的改变
为了探究nr3c1对骨发育影响的机制,采用qRT-PCR方法检测了nr3c1-突变组和PN处理组的骨代谢相关基因的表达水平。结果表明,nr3c1-突变体的nr3c1基因表达较WT组下调。在PN处理组,nr3c1表达下调,但不显著(图7A)。
研究表明,mmp9、mmp13和sparc参与细胞外基质(ECM)功能,并通过影响ECM代谢参与骨代谢[9-10]。mmp9和mmp13在以前的研究中已经被确定为PN的靶基因,这两个基因的表达在PN处理后显著增加(图7B)。mmp9和mmp13的表达与WT中没有显著差异,而在nr3c1突变体、5 DPF和8 DPF幼体中,这两个基因的表达降低(图7B、7C)。在WT和nr3c1-突变组中,PN在5 DPF后上调sparc的表达。与野生型相比,nr3c1-突变体的sparc表达无显著差异。在8 DPF时,sparc的表达水平在4个样本组之间没有显著差异(图7D)。
runx2b、entpd5a、spp1和alp是涉及骨代谢的成骨细胞相关基因[11-12]。qRT-PCR结果显示,与对照组相比,nr3c1-突变体中的成骨细胞分化基因runx2b的表达不受PN处理的影响(图7E)。在5 DPF和8 DPF,PN治疗后entpd5a的表达与对照组相比没有显著变化(图7F)。在nr3c1-突变体中,entpd5a的表达水平显著下调,PN没有改变其在nr3c1-突变体中的表达(图7F)。在nr3c1-突变体中,spp1的表达在5 DPF时表达降低,但差异不显著(图7G)。在8 DPF时,与WT相比,nr3c1-突变体中spp1的表达水平显著下调,而PN并不改变nr3c1-突变体中的这种下调(图7G)。碱性磷酸酶(ALP)是成骨细胞的标志物。在5 DPF时,PN没有诱导alp的表达,而在8 DPF时,PN显著增加了alp的表达(图7H)。与WT组相比,在5 DPF和8 DPF nr3c1-突变体中,alp的表达下调,而在nr3c1-突变体中,PN上调了alp的表达,但其水平没有达到WT组的水平(图7H)。
ctsk、acp5a、rankl和opg与骨代谢过程中的破骨细胞有关[13-15]。qRT-PCR结果显示,与WT组相比,PN处理组ctsk在5 DPF时表达上调,在nr3c1-突变体中表达上调(图7I)。在8 DPF时,PN处理不改变ctsk的表达。在nr3c1-突变体中,ctsk的表达水平显著下调,并不受PN的影响(图7I)。与WT组相比,nr3c1突变体中acp5a的表达下调,PN在5 DPF时促进了nr3c1-突变体中acp5a的下调(图7J)。rankl和opg是两个功能相反的配体,它们分别编码激活和抑制破骨细胞分化的蛋白。在5 DPF时,PN处理组和nr3c1-突变体的两个基因表达下调。PN处理组5 DPF时rankl和opg的表达水平与WT组相比无显著差异,而在8 DPF时,nr3c1-突变体的rankl和opg表达水平较WT组明显下调。在nr3c1-突变组,PN上调了opg的表达,但不影响ranklL的表达(图7K~7L)。以上数据表明,nr3c1影响斑马鱼破骨细胞及成骨细胞相关基因的表达。
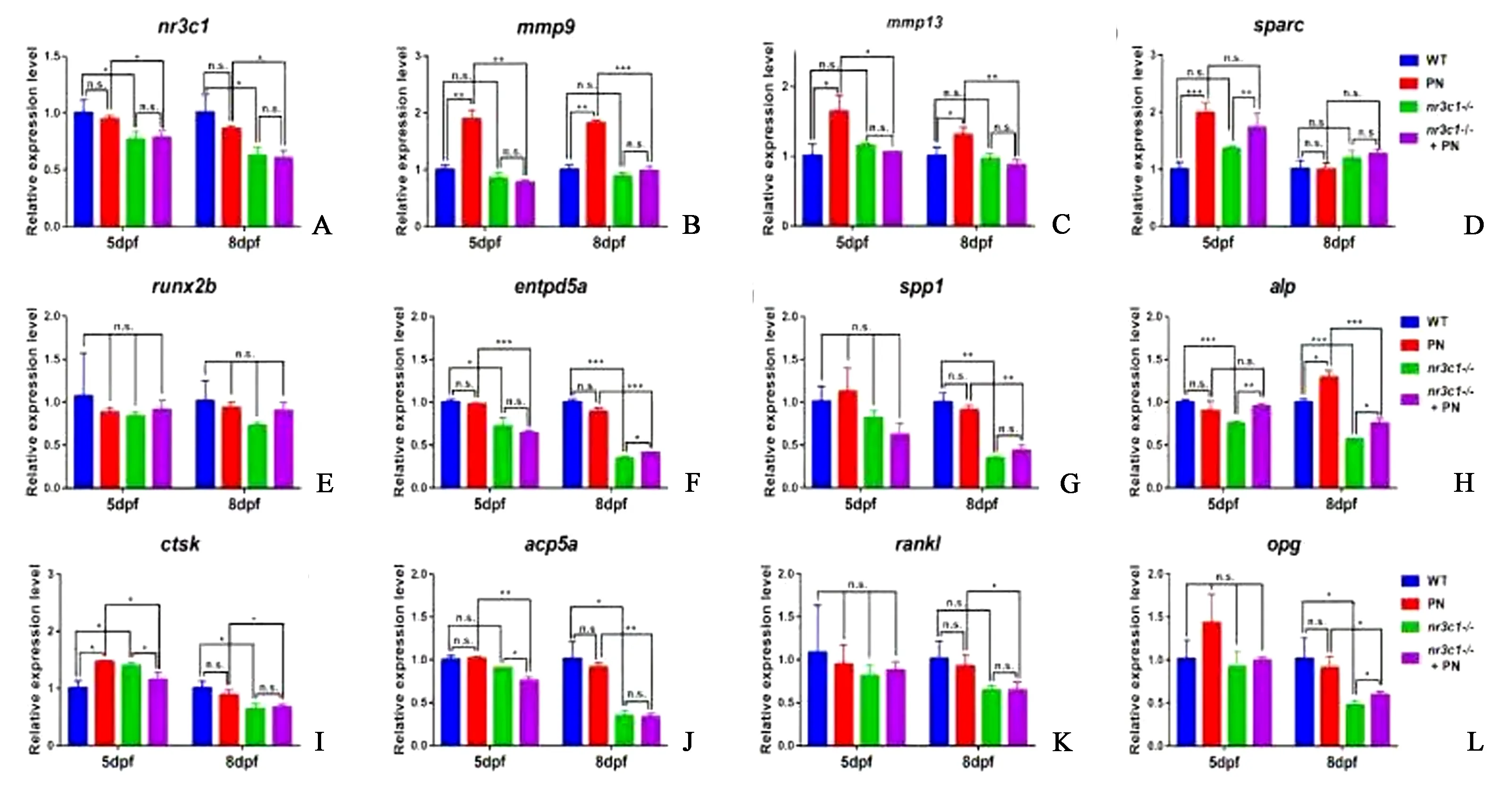
图7 nr3c1突变改变骨代谢相关基因的表达(用25 μmol/L PN处理WT和nr3c1突变胚胎,并在受精后第5天和第8天收集)
2.5 GC-NR3C1介导的骨相关基因表达
双荧光报告实验结果表明,PN或nr3c1单独上调mmp9的表达,PN可进一步增强mmp9的表达(图8A)。对碱性磷酸酶(alp)表达的分析显示,与mmp9的结果相似(图8B)。单独使用PN并不能显著上调acp5a,而单独添加nr3c1则显著上调acp5a。PN和nr3c1的加入增强了acp5a的这种上调表达(图8C)。这些结果表明PN和NR3C1协同调节骨代谢相关基因的表达。
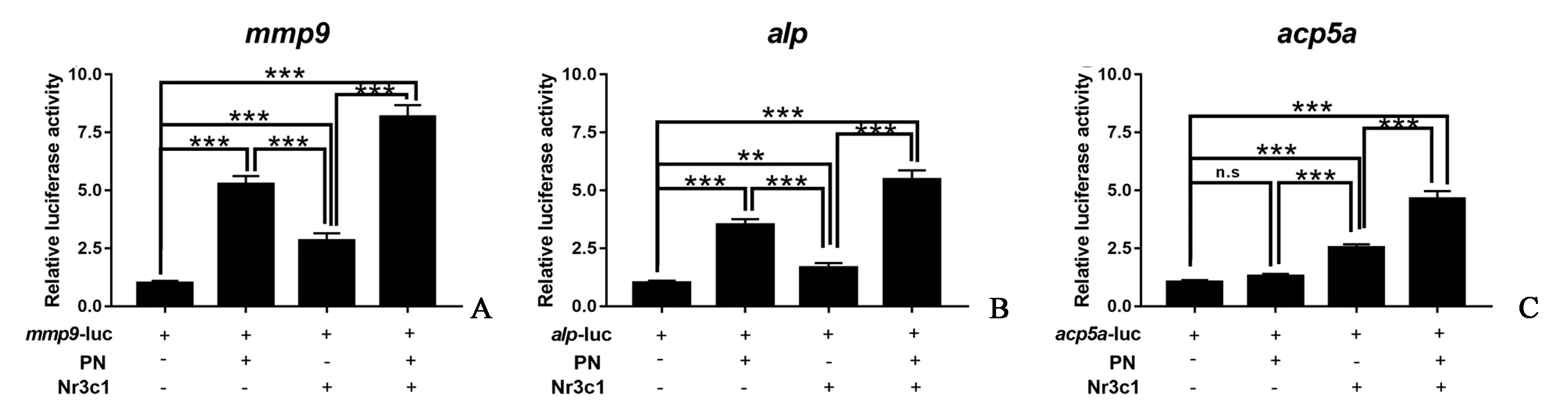
图8 PN通过nr3c1调节骨代谢相关基因的表达(双荧光素酶报告分析PN和nr3c1对骨代谢相关基因表达的影响)
3 讨论
Paul等[16]发现PN显著促进颅面骨的发育。本研究结果表明,PN对颅面骨发育没有影响,这可能是由于PN浓度不同所致。然而,在nr3c1-突变体中,每个软骨参数都显著降低,这表明nr3c1的缺失抑制了颅面骨的发育。此外,25 μmol/L PN改变了由nr3c1缺失介导的软骨表型。这一结果表明,过量的GC可以部分补偿nr3c1信号通路的缺失(糖皮质激素受体非依赖性通路)。
骨矿化是成骨细胞成熟和骨形成的重要过程[17]。在骨矿化涉及的蛋白质中,entpd5a可以调节磷酸盐的稳定性[18]。根据定量结果,在nr3c1-突变体中entpd5a的表达显著下调,但PN单独不能抑制其表达,表明NR3C1是通过entpd5a途径参与骨矿化的介质之一。在斑马鱼nr3c1-突变体中,spp1的表达显著下调,表明nr3c1也可以通过spp1影响骨矿化。先前的研究发现,高浓度的PN会抑制斑马鱼成骨细胞的分化和数量[19]。在小鼠中,nr3c1被发现降低成骨细胞分化和相关基因的表达[20]。因此,成骨细胞是nr3c1发挥作用的靶细胞,并对骨代谢产生影响,但nr3c1如何调控entpd5a和spp1从而影响骨矿化尚需进一步分析。
破骨细胞能够吸收骨组织,在骨重建中发挥重要作用[15]。先前的动物和细胞实验表明,GCs可以促进破骨细胞分化,并会导致GIOP[21]。然而,在斑马鱼中,PN被发现可以减少破骨细胞数量并增加凋亡[19];在小鼠中,破骨细胞可以通过GR介导的途径走向凋亡,随后的研究表明PN可以抑制破骨细胞相关基因的表达。本研究发现在nr3c1-突变体中ctsk、acp5a、rankl和opg表达下调,表明nr3c1可以调节许多破骨细胞基因的表达。RANKL/OPG比例对破骨细胞分化至关重要[22]。而GR-二聚体是RANKL/OPG轴的重要调节因子[23]。定量结果表明,OPG和RANKL都是nr3c1的靶基因。本研究未对nr3c1-突变体中RANKL/OPG的比例进行统计分析,该比例是否影响成骨细胞的分化和增殖还有待进一步分析。
在细胞核内,激活的nr3c1以二聚体的方式与GRE元件或其他转录因子结合,以调节基因表达。在本研究评估的基因中,GC/Nr3c1可以通过4种方式调节与骨代谢有关的基因的表达。第一种途径是通过GC-Nr3c1途径,涉及mmp13、mmp9和alp。其次,在nr3c1突变的情况下,GC可以通过多种途径调节骨代谢相关基因的表达,例如sparc。第三,nr3c1调控基因的表达与GCs无关,如entpd5a、spp1、acp5a、opg和rankl。第四,GC调节基因的表达既可以通过nr3c1途径,也可以通过非nr3c1途径,如alp。GC-Nr3c1途径是GN发挥调节骨代谢相关基因作用的经典途径,主要通过调节细胞外基质从而影响骨结构和骨相关细胞的微环境。在nr3c1突变的情况下GC也可通过非nr3c1途径发挥调节作用,如cAMP反应元件结合蛋白、激活蛋白(AP)-1、核因子-κB等。
4 结论
本研究利用CRISPR/Cas9技术成功地获得了斑马鱼nr3c1-突变体。利用该模型发现Nr3c1影响软骨发育和骨矿化,并且Nr3c1突变改变了细胞外基质、成骨细胞和破骨细胞相关基因的表达。进一步的实验表明,GC/Nr3c1通过Nr3c1依赖和非Nr3c1途径调控细胞外基质、成骨细胞和破骨细胞相关基因的表达。本研究为确定Nr3c1在骨代谢和发育中的作用提供了依据,同时为临床治疗糖皮质激素性骨质疏松症确定了新的效应靶点。
猜你喜欢
中国老年学杂志(2022年19期)2022-11-21
作物学报(2022年2期)2022-11-06
安徽医科大学学报(2022年6期)2022-07-13
中国兽医学报(2022年4期)2022-06-17
作物学报(2022年8期)2022-05-29
蔬菜(2021年7期)2021-11-27
小天使·二年级语数英综合(2021年8期)2021-08-16
家庭医学(2020年12期)2020-01-20
作文评点报·作文素材小学版(2016年8期)2016-03-16
第二课堂(小学版)(2015年2期)2015-05-22
