
纳米硒对草莓叶片光合特性及叶绿素荧光参数的影响
2022-08-26 07:02:16张雪洁赵世宇仝璐遥杨天佑单长卷
江苏农业科学 2022年16期
张雪洁, 周 梦, 赵世宇, 仝璐遥, 杨天佑, 张 浩, 单长卷
(1.河南科技学院,河南新乡 453003; 2.长江大学,湖北荆州 434023)
草莓是广受欢迎的小浆果之一,其果实营养丰富,口味甜美,具有较高的经济价值和广阔的商业前景。提升草莓果实品质及产量以达到经济效益最大化,已成为近年来国内外研究的重点。光合作用是植物体内最重要的生理活动,植物体内绝大多数碳源均来源于光合作用。同时,大量研究表明,植物叶片光合作用的强弱程度,对植物果实的产量及品质起到至关重要的作用。因此,提升植物叶片光合能力,对提高果实产量及改善品质具有重要意义。
硒元素对植物生长发育,特别是在叶片光合作用方面,同样可以起到促进作用。冯涛等研究表明,通过根施硒肥,梨树叶片净光合速率、胞间CO浓度、蒸腾速率以及气孔导度均得到显著提升,同时,果实中可溶性糖、维生素C等含量也随之提升。钟松臻等研究表明,适宜浓度的亚硒酸钠(NaSeO) 显著提升了水稻叶片中光合色素含量及净光合速率。郭美俊等通过对谷子叶片喷施NaSeO发现,其叶片叶绿素含量、净光合速率、气孔导度、PSⅡ实际光化学效率以及表观电子传递速率等指标均得到显著提升,最终提升了谷子产量。笔者前期研究也表明,对草莓植株进行氨基酸硒处理,草莓叶片光合色素含量、净光合速率等指标均显著上升,进而提高草莓果实产量和改善品质。近年来,红色零价态纳米硒(SeNPs)的出现,在食品及医疗领域备受关注,由于纳米硒活性高、无毒害、易吸收等优势,现已成为生物强化富硒的最佳选择。目前,有关于硒元素调控植物叶片光合特性的研究已有相关报道,但有关于新型纳米硒与草莓植株叶片光合生理特性的研究尚属空白。因此,探究SeNPs对草莓叶片光合特性的影响,对其应用于农业生产具有指导意义。
本研究2021年3—5月采用盆栽试验,以不同浓度纳米硒(SeNPs)溶液对草莓植株进行叶面喷施处理,测定果实生长发育3个时期(大果期、转色期、成熟期)叶片光合生理参数、叶绿素荧光参数及叶绿素含量相对值(SPAD值),同时测定果实糖度、单果质量和植株总生物量,最终筛选合适的浓度,以期为SeNPs提升草莓光合能力和在农业生产中的应用提供理论依据。
1 材料与方法
1.1 试验材料
本试验所用材料为我国主栽品种章姬草莓幼苗,将幼苗定植于塑料盆内,塑料盆规格为 14 cm×15 cm,容积约为2 L,每盆装培养土2 kg。试验所用纳米硒购买自上海四通纳米技术港有限公司,纳米硒粒子颗粒直径在20~60 nm之间。
1.2 试验设计
幼苗定植后,每盆浇水400 mL,置于室内进行缓苗处理。3 d后,选取长势良好的幼苗置于人工物候箱中进行培养。培养期间,利用称质量法控制持水量在70%~75%之间。本研究共设置4个处理,即T1(10 mg/L)、T2(20 mg/L)、T3(40 mg/L) 3个浓度梯度SeNPs溶液处理以及对照组(CK),其中对照组仅采用等量清水进行处理。待草莓现蕾期开始,每8 d喷施1次,以叶面喷施的形式对草莓植株叶片进行SeNPs处理,共喷施3次,每次每盆喷施溶液10 mL。每个处理15个重复,共计60盆。
1.3 测定项目与方法
分别在大果期(花后18 d)、转色期(花后 23 d)、成熟期(花后28 d),选取叶片大小、叶龄相近,无损伤、病虫害且完全展开的叶片进行测定。
1.3.1 SPAD值 SPAD值以SPAD-502叶绿素含量测定仪进行测定。
1.3.2 光合生理参数 采用Li-6800便携式光合仪进行测定,时间选在天气晴朗的08:00—12:00。测定指标包括净光合速率()、蒸腾速率()、胞间CO浓度()、CO总导度(Gtc)、气孔限制值()以及水分利用效率(WUE)。
1.3.3 叶绿素荧光参数 采用PAM-2500便携式调制荧光仪进行测定,所测定指标包括PSⅡ实际光化学效率[Y(Ⅱ)]、PSⅡ最大光化学量子产额(/)、PSⅡ有效光化学量子产额(′/′)、表观电子传递速率(ETR)、非光化学淬灭系数(NPQ)、光化学淬灭系数()、快速光曲线初始斜率(alpha)以及最大电子传递速率(ETR)。
1.3.4 果实糖度、单果质量、总生物量 果实糖度采用手持折光仪进行测定,单果质量采用电子天平进行测定,植株总生物量以烘干法进行测定。
2 结果与分析
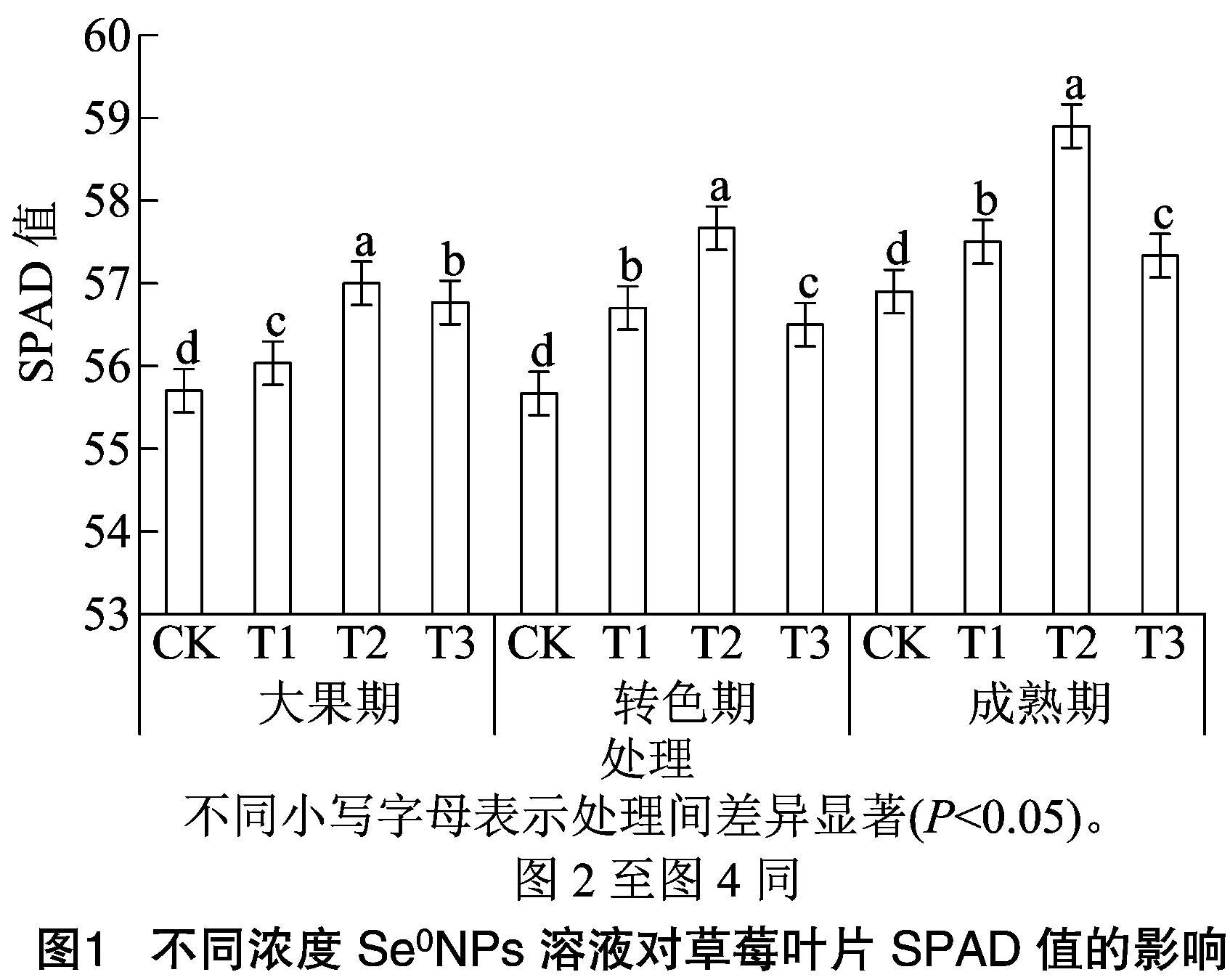
2.1 不同浓度Se0NPs溶液对不同生育时期草莓叶片相对叶绿素含量的影响
如图1所示,不同浓度SeNPs对不同时期草莓叶片SPAD值的影响呈现相似的变化趋势。在大果期,T1、T2、T3处理使SPAD值分别较对照组提升0.6%、2.3%、1.9%;在转色期,T1、T2、T3处理使SPAD值分别较对照组提升1.8%、3.6%、1.4%;在成熟期,T1、T2、T3处理使SPAD值分别较对照组提升1.1%、3.5%、0.8%。各时期SPAD值均随SeNPs浓度的升高呈现先升后降的变化趋势,均以T2处理提升最为显著。表明SeNPs显著提升叶片的SPAD值。
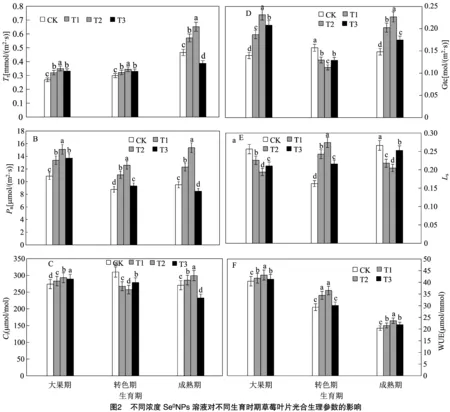
2.2 不同浓度Se0NPs溶液对不同生育时期草莓叶片光合生理参数的影响
如图2-A所示,在大果期,不同浓度SeNPs处理下叶片均较对照组显著提升,T1、T2、T3处理分别提升19.2%、30.0%、23.5%,其中以T2处理提升最为显著;在转色期,不同浓度处理下叶片亦显著高于对照,T1、T2、T3处理分别较对照组提升7.7%、15.4%、3.6%,其中以T2处理提升最为显著,T1和T3处理间差异并不明显;在成熟期,T1、T2处理均显著高于对照组22.5%、39.9%,而T3处理则显著低于对照组16.9%。表明SeNPs显著提升大果期、转色期草莓叶片,但在转色期这种提升趋势则会相对放缓,而在成熟期T1、T2处理下则显著提升,而T3处理却显著下降。
如图2-B所示,在大果期,各浓度SeNPs处理下叶片均较对照组显著提升,T1、T2、T3处理分别较对照组提升23.0%、38.8%、26.1%,其中以T2处理提升最为显著;在转色期,各浓度处理叶片亦均显著高于对照,T1、T2、T3处理分别较对照组提升26.3%、43.5%、6.2%,其中以T2处理提升最为显著;而在成熟期,仅T1、T2处理显著高于对照组29.4%、61.7%,而T3处理则显著低于对照10.8%。表明SeNPs在大果期、转色期显著提升,而在成熟期仅提升T1、T2处理的,并显著降低T3处理的。
如图2-C所示,在大果期,各浓度SeNPs处理下叶片均较对照组显著提升,T1、T2、T3处理分别较对照组提升3.3%、7.0%、5.5%,其中以T2处理提升最为显著;而在转色期,各浓度处理则较对照组显著下降,T1、T2、T3处理分别较对照组下降13.7%、16.9%、10.1%,其中以T2处理下降最为显著;在成熟期,各浓度处理下变化不一,T1、T2处理分别显著高于对照组5.6%、10.6%,T3处理则显著低于对照14.1%。表明SeNPs显著提升大果期草莓叶片,以T2处理最为显著,同时显著降低转色期的,以T2处理下降最为显著,而在成熟期,T1、T2处理下显著提升,而T3处理下该值则显著降低。
如图2-D所示,在大果期,各浓度SeNPs处理下叶片Gtc均得到显著上升,T1、T2、T3处理分别较对照组提升33.9%、65.6%、48.9%,其中以T2处理上升最为显著;在转色期,不同浓度处理下该值则会显著低于对照组,T1、T2、T3处理分别较对照组降低17.6%、28.1%、17.9%,其中亦以T2处理下降最为显著;而在成熟期,各浓度处理下该值亦均显著高于对照组,T1、T2、T3处理Gtc较对照组分别显著提升36.7%、53.0%、17.9%。表明SeNPs能够显著提升草莓大果期和成熟期Gtc值,且均以T2处理提升最为显著;SeNPs显著降低转色期的Gtc值。
如图2-E所示,在大果期,各浓度SeNPs处理下叶片均出现显著降低,T1、T2、T3处理分别较对照组降低11.9%、24.4%、17.8%,其中以T2处理下降最为显著;在转色期,各浓度处理下叶片值则出现显著上升的趋势,T1、T2、T3处理分别较对照组提升49.3%、68.8%、32.8%,其中以T2处理上升最为显著;在成熟期,各浓度处理下的亦呈显著下降趋势,T1、T2、T3处理分别较对照组显著降低18.1%、23.1%、5.1%,其中以T2处理下降最为显著。表明SeNPs显著降低大果期和成熟期草莓叶片值,均以T2处理最为显著;显著提升转色期的值,且亦以T2处理最为显著。
如图2-F所示,不同浓度SeNPs对草莓叶片WUE产生较为相似的影响,各时期均呈单峰变化趋势,即在T2处理达到峰值,随之开始下降。在大果期,T1、T2、T3处理分别较对照组提升3.2%、6.3%、1.9%;在转色期,T1、T2、T3处理分别较对照组提升17.3%、20.8%、1.9%;在成熟期,T1、T2、T3处理分别较对照组提升5.7%、14.7%、6.3%。表明SeNPs能够显著提升植株水分利用效率。
2.3 不同浓度Se0NPs溶液对不同生育时期草莓叶片叶绿素荧光参数的影响
如图3-A所示,在大果期,各浓度SeNPs处理下Y(Ⅱ)均得到显著提升,T1、T2、T3处理分别较对照组提升23.4%、30.3%、26.1%,其中以T2处理提升最为显著;在转色期,不同浓度处理下Y(Ⅱ)变化不一,T1、T2处理分别高于对照3.0%、7.1%,其中T2处理显著高于对照组,T1处理则仅略高于对照,而T3处理却显著低于对照组8.6%;在成熟期,不同浓度处理下Y(Ⅱ)值均较对照组有所提升,T1、T2、T3处理分别较对照组提升4.6%、17.1%、2.3%,其中T2处理显著高于对照组,T1、T3处理则仅略高于对照组。表明SeNPs提升了大果期、成熟期的Y(Ⅱ)值,且均以T2处理提升最为显著,而在转色期,T1、T2处理下该值会提升,以T2处理上升最为显著,T3处理该值则会显著降低。
如图3-B所示,在大果期,各浓度SeNPs均显著提升了/,T1、T2、T3处理下该值分别较对照组提升0.1%、1.0%、0.7%,其中以T2处理提升最为显著;在转色期,不同浓度处理下/变化不一,T1、T2处理分别较对照组提升0.3%、0.8%,而T3处理则显著低于对照组0.7%;在成熟期,各处理变化趋势与转色期相似,T1、T2处理显著高于对照组0.9%、0.5%,而T3处理则略低于对照组。表明SeNPs能够提升大果期的/,而在转色期、成熟期,T1、T2处理下该值显著上升,T3处理则下降。
如图3-C所示,在大果期,各浓度SeNPs处理下叶片′/′均得到显著提升, T1、T2、T3处理分别较对照组显著提升2.1%、2.7%、1.5%,其中以T2处理提升最为显著;在转色期,仅T1、T2处理下该值高于对照组6.7%、11.8%,其中以T2处理提升最为显著,而T3处理则略低于对照组0.2%;在成熟期,各浓度处理下该值则呈现随硒浓度上升而逐渐上升的趋势,T1、T2、T3处理分别显著高于对照组3.3%、4.2%、6.1%,其中以T3处理提升最为显著。表明SeNPs提升了大果期、成熟期的′/′值,且这2个时期分别以T2、T3处理提升最为显著,而在转色期,仅T1、T2处理下该值得到提升,以T2处理最为显著,T3处理下该值则会出现小幅度下降。
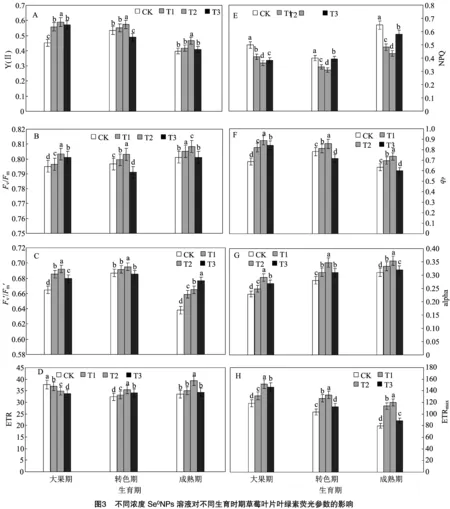
如图3-D所示,在草莓大果期,各浓度SeNPs溶液处理下叶片ETR值会出现随硒浓度提升不断下降的变化趋势,T1、T2、T3处理分别较对照组显著降低1.7%、7.2%、10.2%,其中以T3处理下降最为显著;在草莓转色期,不同浓度处理下该值则会较对照组显著提升,T1、T2、T3处理分别提升2.4%、9.6%、5.1%,其中以T2处理提升最为显著;在草莓成熟期,不同浓度处理下该值亦较对照组有所提升,T1、T2、T3处理分别较对照组提升4.4%、17.1%、2.3%,其中T2处理显著高于对照组,其他处理仅略高于对照组。表明SeNPs溶液处理会显著降低大果期草莓叶片ETR值,以T3处理最为显著,同时提升转色期、成熟期叶片ETR值,且该2个时期均以T2处理提升最为显著。
如图3-E所示,在草莓大果期,各浓度SeNPs溶液处理下叶片NPQ值均较对照组显著降低,T1、T2、T3处理分别降低17.8%、27.3%、23.4%,其中以T2处理下降最为显著; 在草莓转色期,不同处理叶片该值亦显著低于对照,T1、T2、T3处理分别较对照组降低16.4%、22.4%、1.2%,亦以T2处理下降最为显著;在草莓成熟期,T1、T2、T3处理叶片该值分别低于对照组10.7%、33.2%、25.7%,其中以T2处理下降最为显著。表明SeNPs溶液处理会显著降低各时期草莓叶片ETR值,且各时期均以T2处理下降最为显著。
如图3-F所示,在大果期,各浓度SeNPs处理下值均显著高于对照组,T1、T2、T3处理分别较对照组提升19.5%、29.3%、22.7%,其中以T2处理提升最为显著;在转色期,T1、T2处理值均显著高于对照,分别较对照组提升4.0%、9.7%,以T2处理提升最为显著,而T3处理则较对照组显著降低8.3%;在成熟期,不同浓度处理下该值变化趋势与转色期相似,即T1、T2处理较对照组显著提升10.1%、16.2%,而T3处理则显著低于对照组5.4%。表明SeNPs能够提升大果期值,同时T1、T2处理会提升转色期、成熟期的,而T3处理值在这2个时期则显著下降,且均以T2处理提升最为显著。
如图3-G所示,草莓各时期各浓度SeNPs处理下叶片alpha值均较对照组有显著提升。在大果期,T1、T2、T3处理分别较对照组提升8.7%、27.1%、17.0%;在转色期,T1、T2、T3处理分别较对照组提升10.6%、23.6%、10.2%;在成熟期,T1、T2、T3处理分别较对照组提升7.1%、13.8%、28.9%,且各时期均以T2处理提升最为显著。表明SeNPs能显著提升各时期草莓叶片alpha值。
如图3-H所示,草莓各时期各浓度SeNPs处理下叶片ETR值亦均较对照组有显著提升。在大果期,T1、T2、T3处理分别较对照组提升10.8%、28.1%、23.8%;在转色期,T1、T2、T3处理分别较对照组提升22.7%、28.7%、8.9%;在成熟期,T1、T2、T3处理分别较对照组提升42.8%、50.6%、10.9%,且各时期均以T2处理提升最为显著。表明SeNPs能够提升各时期草莓叶片ETR值。
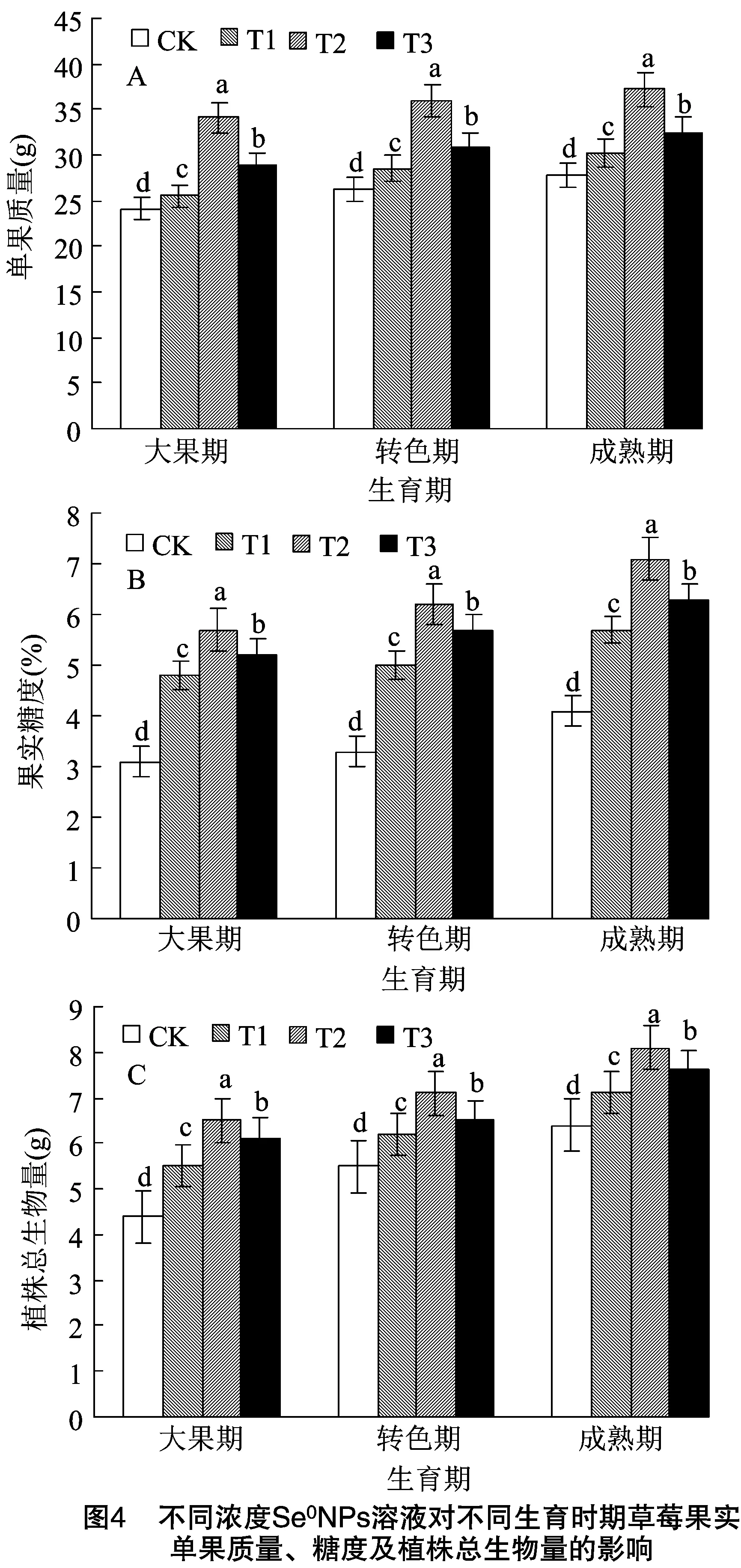
2.4 不同浓度Se0NPs溶液对不同生育时期草莓果实单果质量、糖度及植株总生物量的影响
相比对照组,SeNPs显著提升各时期果实单果质量,且各个时期均以T2处理提升最为显著,T3处理次之,T1处理提升幅度最小(图4-A)。此外,SeNPs处理下,各时期糖度及植株总生物量均呈现与单果质量相似的变化趋势,即先升高后降低,且均以T2处理效果最佳(图4-B、图4-C)。表明SeNPs能够显著提升不同生育时期草莓果实单果质量、糖度及植株总生物量。
3 讨论与结论
光合作用是植物有机物质形成的基础,与植物果实品质和产量的形成存在密切关系。因此,研究纳米硒溶液对草莓叶片光合特性的影响,对进一步探究其促进草莓果实品质及产量提升的成因具有重要意义。目前,有关于纳米态硒对植物光合特性影响的研究较少,关于纳米硒调控草莓叶片光合特性的研究尚未见报道。本研究结果表明,叶面喷施SeNPs溶液处理能够对不同生育时期草莓叶片不同光合指标起到积极作用,从而提升草莓光合效率,进而促进植株干物质、果实糖度的积累以及单果质量的提升。
史丽娟等研究表明,亚硒酸钠能够提升各时期谷子叶片SPAD值,这与本研究结果基本一致。然而,赵双玲等在水稻上的研究却出现了相反的结果,这可能是由于硒的形态以及供试材料差异导致的。迄今有关于硒对草莓叶片SPAD影响的研究仍鲜有见报道,本研究结果表明,SeNPs能够对不同生育时期草莓叶片的SPAD值起到显著的提升作用,说明SeNPs能够提升草莓叶片相对叶绿素含量,同时意味着SeNPs对植物氮素的吸收起到了一定促进作用。有研究曾经发现,有机态硒(SeMet)能够显著提升不同生育时期草莓叶片中各种光合色素的含量,这也进一步证实了硒对草莓叶片光合色素含量提升的有效性。造成这种现象的原因可能是由于硒的抗氧化作用延缓了叶绿体的衰老,同时亦有研究表明,硒能够促进植物对多个有益于叶绿素合成的微量元素的吸收。在这些生理作用下,草莓SPAD值得以提升,这也为草莓叶片光合效率的提升提供了基础。
郭美俊等研究表明,硒能够提升各时期谷子叶片Y(Ⅱ)、ETR、和的值;高尚等的研究中也曾经发现,有机态硒能够显著提升不同生育时期草莓叶片Y(Ⅱ)、′/′、/、ETR以及的值,同时显著降低叶片NPQ值。本研究结果表明,除高浓度SeNPs处理外,适量浓度SeNPs处理能够提升不同生育时期草莓叶片Y(Ⅱ)、/、′/′以及值,同时显著降低不同生育时期叶片NPQ的值,这与先前的研究结果基本一致。此外,本研究还发现,SeNPs处理下转色期、成熟期的草莓叶片ETR均有所提升,这与前人研究结果一致,但大果期叶片ETR值却呈现随硒浓度提升逐渐下降的变化趋势,有关这种现象的原因还有待进一步研究阐明。另外,本研究还发现,SeNPs处理能够提升各时期叶片alpha和ETR的值,该2个指标分别反映了叶片光能利用效率和最大电子传递速率,该2个指标的提升进一步证实了硒对叶片光能利用及电子传递能力的提升作用。上述现象说明SeNPs不仅提升了草莓叶片中光能及电子传递的能力及效率,同时还减少了能量损耗,从而达到提升叶片光合效率的目的。闫帅等研究表明,硒能够改善植物叶片组织结构,进而提升其光合能力,硒还会提升叶片光合色素的含量,保护光合系统不受伤害。这些都是本研究中叶片光合荧光参数指标得到提升的原因。
蔡天革等研究表明,通过喷施适量的硒显著提升了荞麦叶片以及。本研究结果表明,经过SeNPs处理后草莓叶片的及总体上呈现上升趋势,这与前人研究结果基本一致,但在草莓成熟期,40 mg/L浓度SeNP处理草莓果实叶片的上述2个指标则出现了显著下降,这可能是由于过量的硒对草莓叶片造成了毒害,导致其光合系统受到损害,进而导致光合能力的下降。此外,本研究结果发现,除高浓度处理外,草莓大果期、成熟期,硒对叶片光合能力的提升主要是依靠调控气孔因素所导致的,而在转色期则是由于非气孔因素导致的,类似的现象在我们之前的研究中也有发现。造成这种现象的原因,很可能是由于草莓叶片在不同生育时期固有的生理活动差异导致的。武林楠等研究表明,硒能够提升葡萄水分利用率,本研究表明SeNPs显著提升不同生育时期草莓植株水分利用率,这与相关研究结果基本一致,造成这种现象的原因可能是因为SeNPs对气孔开放程度的提升,增加了CO通量,同时提升了叶片光合能力,最终导致更多干物质的合成。
猜你喜欢
山东农业科学(2022年10期)2022-11-17 06:28:02
花卉(2022年17期)2022-09-06 02:19:02
食品安全导刊(2021年21期)2021-08-30 08:21:56
落叶果树(2021年6期)2021-02-12 01:29:28
房地产导刊(2020年9期)2020-10-28 08:37:24
中国果树(2020年2期)2020-07-25 02:14:18
食用菌(2020年1期)2020-02-18 08:17:22
中国农资(2019年50期)2020-01-02 08:52:54
今日农业(2019年14期)2019-01-04 08:57:40
浙江柑橘(2016年4期)2016-03-11 20:13:01
