
桑天牛纤维素酶编码基因的克隆及分泌表达
2022-08-26 09:02潘宇杰梁笑玲张楚玥陈少珍林晓玲李虹仪
江苏农业科学 2022年16期
张 茂, 潘宇杰, 梁笑玲, 张楚玥, 陈少珍, 林晓玲, 李虹仪
(韶关学院英东生物与农业学院,广东韶关 512005)
饲料原料的短缺问题一直影响着畜牧业的发展,成为我国畜牧业面临的一大挑战,因此需要不断开发饲料资源。纤维素是自然界年产量巨大的可再生资源,然而这些宝贵的纤维素资源并没有被有效地利用。此外,猪和家禽日粮中也存在大量纤维素因含有非淀粉多糖(non starch polysaccharides,NSP)等抗营养因子而降低了其他营养物质的吸收利用率而不能被畜禽利用。饲用酶制剂被认为是目前能有效解决养殖领域中饲料安全、饲料原料缺乏的新型饲料添加剂。为了促进不同类型饲粮纤维的合理开发和利用,可通过添加纤维素酶来提高动物机体对饲粮纤维的消化利用率,改善动物肠道健康,提高动物生产性能,以缓解饲料资源危机。纤维素酶来源有细菌、真菌、昆虫等,同时也有大量不同来源的基因被克隆,如芽孢杆菌、木霉、青霉、牛瘤胃微生物、天牛、白蚁、白蚁体内细菌、福寿螺等。动物源(如天牛、白蚁、福寿螺等)消化道分泌的纤维素酶对纤维素展现出较好的水解能力,其应用亦受到广泛关注。桑天牛消化道内源表达的纤维素酶在分解木质纤维素方面具有较高的活性,可进一步将其研究应用于饲料资源的开发。本试验以桑天牛为来源,克隆桑天牛肠道纤维素酶基因,构建体外表达载体并在PK15细胞中进行表达,以期得到高活性的纤维素酶基因用于酶制剂生产或转基因研究,拓宽畜牧业的饲料来源。
1 材料与方法
1.1 主要试剂
TRIzol试剂、反转录试剂盒PrimeScriptRT reagent Kit with gDNA Eraser、LA酶、pMD18-T载体、连接试剂盒DNA Ligation Kit Ver.2.1、限制性内切酶(Ⅰ、Ⅰ)、DL 2000 DNA Marker、PCR检测试剂Ex,均购自TaKaRa公司;胶回收Gel Extraction Kit D2500、质粒提取Plasmid Mini Kit Ⅰ,均购自Omega公司;转染试剂Lipofctamine、细胞培养试剂胎牛血清(FBS)、达尔伯克(氏)改良伊格尔(氏)(DMEM)培养基、磷酸盐缓冲液(PBS),均购自Life公司;羧甲基纤维素钠(CMC-Na)、-葡聚糖,均购自Sigma公司。PK15细胞、表达载体pCMS-EGFP于笔者所在实验室保存。试验于2017年5月至2020年12月在华南农业大学动物科学学院及韶关学院英东生物与农业学院进行。
1.2 引物设计
参考GenBank发布的桑天牛纤维素酶编码基因(AY741064)、(AY451326)的序列,用DNAstar软件进行引物设计,扩增其cDNA序列(长度分别为711、720 bp),分别加入酶切位点Ⅰ、Ⅰ和Kozak序列,引物由深圳华大基因科技有限公司合成,引物序列见表1。

表1 纤维素酶基因扩增引物序列信息
1.3 RT-PCR扩增
将采集的桑天牛在超净工作台中的冰上分离出肠道组织,用TRIzol法提取肠道组织的总RNA,用PrimeScriptRT reagent Kit with gDNA Eraser进行cDNA的合成,反转录反应条件:37 ℃ 15 min,85 ℃ 5 s,4 ℃保存。以cDNA为模板进行PCR扩增,反应体系:0.5 μL LA(5 U/μL),5 μL 10×LABuffer Ⅱ,8 μL dNTP Mixture,各1 μL上、下游引物,4 μL模板,加ddHO至50 μL。反应条件:95 ℃ 3 min;95 ℃ 30 s,56 ℃ 30 s,72 ℃ 1 min,35个循环;72 ℃ 10 min。反应结束后用琼脂糖凝胶电泳进行检测。将PCR产物分别与 pMD18-T 进行连接,然后进行转化,再将单克隆菌液送至测序公司测序,将测序结果与参考序列进行比对分析,选择无移码、缺失、终止突变的克隆进行后续试验。
1.4 酶切
分别以含有、基因的载体为模板进行PCR扩增,然后用限制性内切酶Ⅰ、Ⅰ分别对表达载体pCMS-EGFP及PCR扩增产物进行酶切,酶切体系:5 μL 10×QuickCut Green Buffer,各2.5 μL QuickCutⅠ、Ⅰ,30 μL DNA,加ddHO至50 μL。37 ℃保温1 h,酶切结束后用Gel Extraction Kit进行目的片段的回收。
1.5 表达载体的构建
将回收的载体片段pCMS-EGFP与插入基因DNA片段按照连接试剂盒(DNA Ligation Kit Ver.2.1)说明书比例混合制备成体积为10 μL 的DNA 溶液,加入等体积的10 μL Solution Ⅰ,充分混匀后于 16 ℃ 连接反应30 min,反应结束后将连接液进行转化、涂板,挑单克隆菌落进行培养,用通用引物T3、T7进行菌液PCR,将PCR检测阳性的菌液送往深圳华大基因科技有限公司测序,对测序结果进行比对,分析构建质粒的准确性。
1.6 转染PK15细胞
将PK15细胞在含有5% FBS、1%双抗的DMEM中于37 ℃、5%CO培养箱中培养,用Plasmid Mini Kit Ⅰ试剂盒抽提表达质粒DNA,用Lipofectamine 3000 进行转染。将PK15细胞传代于6孔板中,当细胞汇合度约为60%时进行转染,分别用125 μL Opti-MEM将质粒DNA(2.5 μg)和P3000Reagent(5 μL)进行稀释,然后将稀释的DNA加入到稀释的转染试剂中轻轻混匀,在室温下孵育15 min,将孵育好的混合物加入培养细胞中混匀。将构建的表达载体及空载体分别转染3次,转染后48 h在荧光显微镜下观察增强绿色荧光蛋白(EGFP)的表达情况。
1.7 纤维素酶活性的测定
培养72 h后收集细胞培养液作为粗酶液,采用二硝基水杨酸法(DNS法)在温度为40 ℃、pH值为4.5和5.5的条件下测定其水解活性。以1% CMC-Na 溶液作为底物,并参考NY/T 912—2004《饲料添加剂 纤维素酶活力的测定 分光光度法》测定纤维素酶活性;以0.8%-葡聚糖溶液作为底物,并参考NY/T 911—2004《饲料添加剂-葡聚糖酶活性的测定 分光光度法》测定-葡聚糖酶活性,计算出不同纤维素酶基因表达的粗酶液的纤维素酶活性、-葡聚糖酶活性。
1.8 纤维素酶基因的RT-PCR检测
用细胞刮片将PK15细胞刮下,用TRIzol法提取培养细胞总RNA并反转录成cDNA,用纤维素酶基因、扩增引物分别进行PCR扩增,检测纤维素酶基因在PK15细胞中的表达情况。
1.9 统计分析
酶活性以“平均值±标准差”表示,对不同纤维素酶基因(、)表达的酶活性进行独立样本检验,用SPSS 19.0进行统计学分析,<0.05表示差异显著。
2 结果与分析
2.1 Agc Ⅰ、Agc Ⅱ基因的扩增
以桑天牛肠道组织总RNA反转录得到的cDNA为模板,经PCR扩增后进行琼脂糖凝胶电泳检测,结果获得大小约为730、740 bp的预期条带(图1)。测序结果与目的序列进行比对发现,克隆的基因(711 bp)有4个碱基发生突变,基因(720 bp)发生了8个碱基突变,相似度分别为99.4%(基因)和98.3%(基因);基因编码236个氨基酸,基因编码239个氨基酸,氨基酸比对分析结果显示,与目的基因氨基酸序列的相似度为100.0%,基因与目的基因氨基酸序列的相似度为99.6%,在第56位氨基酸位置上发生了S→N的突变,表明克隆的、基因没有发生移码、缺失、终止突变,可用于后续基因的表达试验。
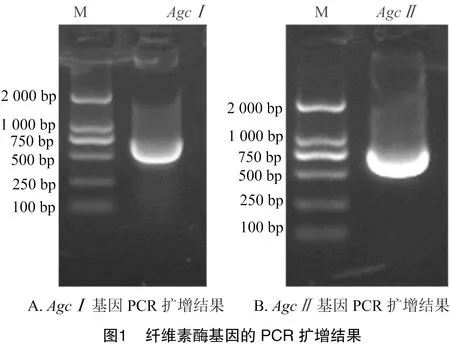
2.2 表达载体的构建
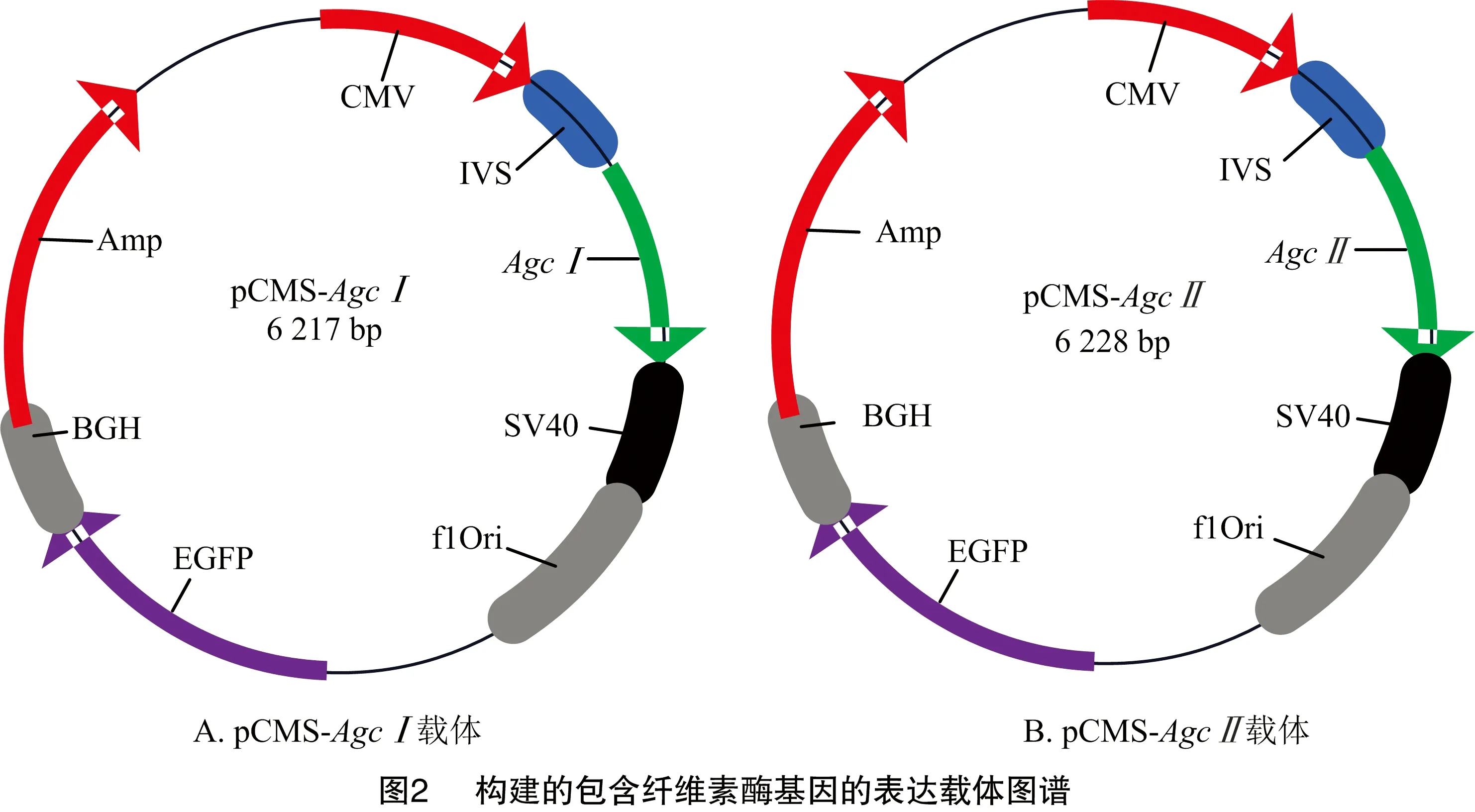
通过酶切、连接、转化、菌液PCR检测,构建出包含纤维素酶基因的表达载体pCMS-、pCMS-(图2),测序结果正确,表明表达载体构建成功。
2.3 转染后EGFP的表达
利用脂质体将表达载体pCMS-EGFP、pCMS-和pCMS-分别转染PK15细胞,48 h后于荧光显微镜下检测转染效果。结果(图3)显示,转染的PK15细胞中均观察到EGFP的表达,视野中绿色荧光细胞数量较多,表明转染效果良好。

2.4 纤维素酶活性的测定
利用DNS法在40 ℃测定纤维素酶基因表达粗酶液的活性。结果(图4)显示,以CMC-Na为底物,在pH值为5.5的环境下,、基因表达的纤维素酶活性分别为(0.09±0.01)、(0.24±0.02) U/mL;在pH值为4.5的环境下,纤维素酶活性分别为(0.07±0.00)、(0.20±0.01) U/mL。以-葡聚糖为底物时,在pH值为5.5的环境下,、基因表达的-葡聚糖酶活性分别为(0.27±0.02)、(0.43±0.05) U/mL;在pH值为4.5的环境下,-葡聚糖酶活性分别为(0.05±0.00)、(0.16±0.01) U/mL。空载体转染的PK15细胞培养液中未检测到纤维素酶、-葡聚糖酶。此外, 在以-葡聚糖、CMC-Na为底物及不同pH值条件下,基因表达的纤维素酶、-葡聚糖酶的活性都显著高于基因(<0.05)。
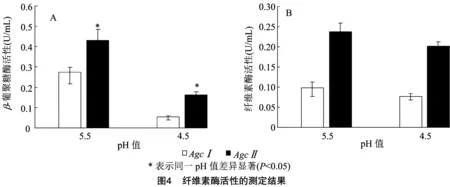
2.5 纤维素酶基因的表达
通过提取转染后PK15细胞总RNA进行RT-PCR检测纤维素酶基因的转录表达,结果(图5)显示,在pCMS-、pCMS-转染PK15细胞cDNA中扩增出、基因目的条带,而在转染空载体pCMS-EGFP的阴性对照PK15细胞中未检测到有纤维素酶基因表达,表明纤维素酶基因在PK15细胞中表达成功。
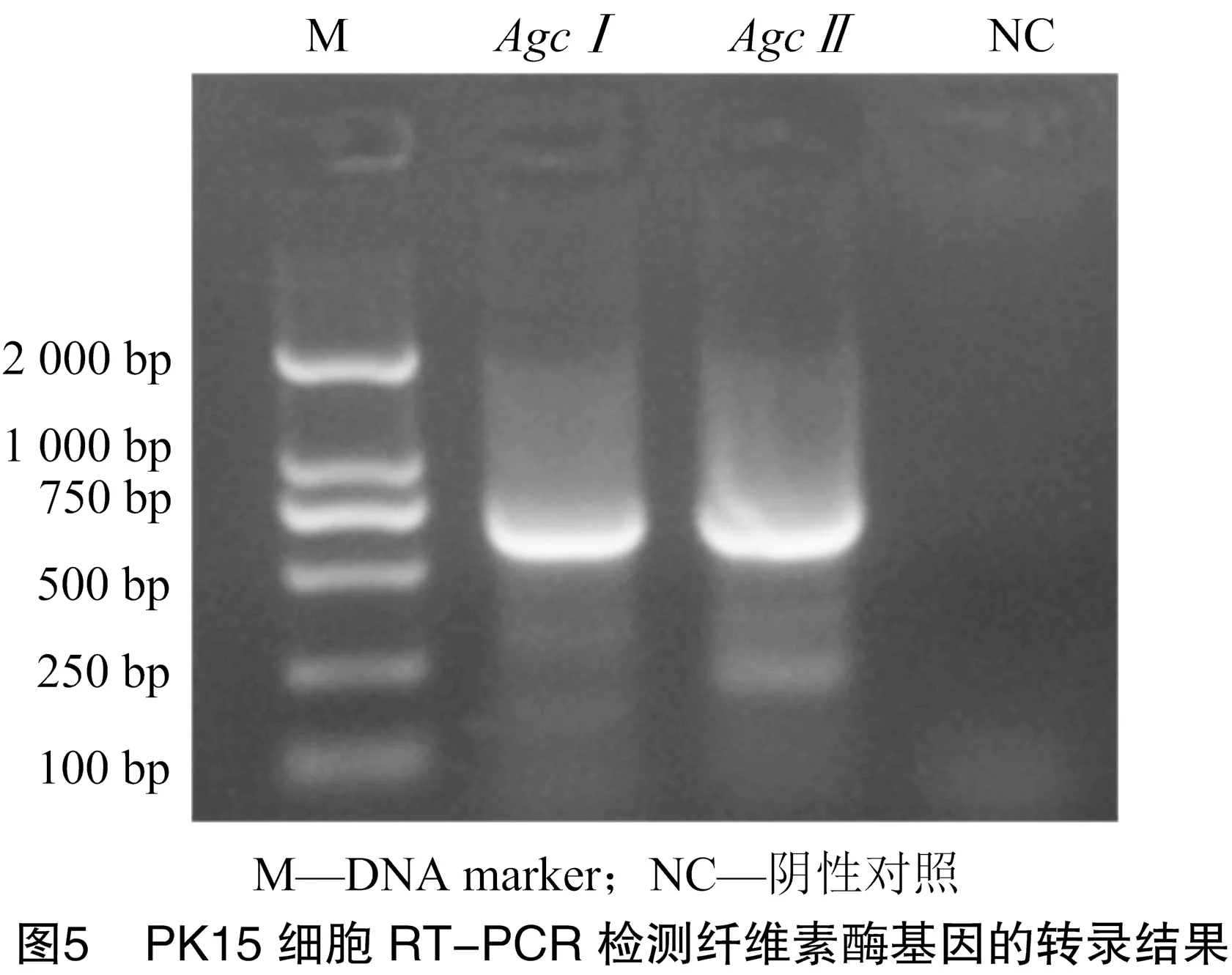
3 讨论
木质纤维素是植物细胞壁的主要组成成分,是地球上最丰富的可再生生物质,占据了地球90%以上光合作用产生的生物质资源,同时也是一种饲料资源。在单胃动物日粮中添加纤维素酶可以有效消除饲料中纤维素的抗营养作用、提高饲料营养物质的消化利用率,同时还能开发饲料资源。为了缓解畜牧业饲料资源短缺的局面,近几年来,大量木本饲料如桑叶、辣木、构树等被开发用于猪和家禽饲料中,这些饲料中纤维素含量过高不易消化,更需要添加高活力的纤维素酶来降解含量较高的木质纤维素,以更好地将这些非常规饲料用于畜禽养殖中。桑天牛主要以树木纤维素为食源,它主要依靠肠道和唾液中的各种纤维素酶、半纤维素酶的协同作用分解食物中的纤维素、半纤维素,桑天牛自身能够分泌纤维素酶并能有效分解木质纤维素,因此本研究选择从桑天牛肠道组织中克隆纤维素酶基因。本试验成功从桑天牛肠道组织中克隆到了纤维素酶编码基因、的cDNA序列,长度分别为711、720 bp,与目的基因序列相比,没有发生移码、缺失、终止突变,2个基因翻译的氨基酸序列与目的基因序列相比相似度都在99%以上。此外,本研究克隆的纤维素酶基因的cDNA序列与基因的相似度为76%,同属于GHF45家族,再次证明桑天牛自身消化道能够分泌纤维素酶体系。
通过构建表达载体分别转染PK15细胞进行表达,在细胞培养液中检测到了纤维素酶活力和-葡聚糖酶活性,表明克隆的纤维素酶基因兼具-葡聚糖酶活性。酶活性测定结果显示,在40 ℃、pH值为4.5及pH值为5.5的条件下,纤维素酶基因表达的酶活性都优于基因表达的酶活性,在pH值为5.5的条件下的酶活性优于pH值为4.5的酶活性,所以基因可作为候选基因进行下一步研究。此外,对PK15细胞进行的RT-PCR检测结果发现了纤维素酶基因、的表达,表明纤维素酶基因在PK15细胞中实现了分泌表达。
本研究克隆了桑天牛纤维素酶基因并构建出真核表达载体,将其转染PK15细胞进行了分泌表达,在基因表达的粗酶液中测定到了较高的纤维素酶及-葡聚糖酶活性,可以为桑天牛纤维素酶基因在转基因及酶制剂制的生产方面提供参考。
猜你喜欢
环球时报(2022-09-20)2022-09-20
纺织科技进展(2021年3期)2021-06-09
陶瓷学报(2021年1期)2021-04-13
今日农业(2020年24期)2020-12-15
兽医导刊(2016年12期)2016-05-17
现代农业(2016年6期)2016-02-28
现代检验医学杂志(2015年4期)2015-02-06
应用化工(2014年11期)2014-08-16
食品工业科技(2014年15期)2014-03-11
食品工业科技(2014年9期)2014-03-11
