
中华绒螯蟹4个养殖群体遗传多样性与遗传结构分析
2022-08-27 10:59胡玉婷潘庭双周华兴
江苏农业科学 2022年16期
胡玉婷, 凌 俊, 江 河, 汪 焕, 潘庭双, 周华兴
(安徽省农业科学院水产研究所/水产增养殖安徽省重点实验室,安徽合肥 230031)
中华绒螯蟹()别称河蟹,俗称大闸蟹,是我国久负盛名的名贵水产品,自然分布于我国长江、黄河、辽河和瓯江等河川及附近地区。20世纪80年代初,随着河蟹育苗技术的突破和养殖技术的开发和推广,我国河蟹养殖进入前所未有的崭新阶段,产量逐年增加,已成为世界首个产蟹大国,同时从蟹苗一直到成蟹培育,全部采用人工方法,这在世界上也是绝无仅有的。目前,中华绒螯蟹养殖已遍布我国从辽宁至浙江的各沿海省份及广大内陆地区,是我国最重要的经济养殖蟹类,2020年全国养殖总产量为77.6万t。但随着中华绒螯蟹养殖产业的规模化发展,跨流域引种和增殖放流等生产活动,造成不同水系中华绒螯蟹种质资源严重混杂和养殖性能退化,尤其受利益驱使和技术所限,中华绒螯蟹育苗场一般选择100 g左右小规格亲本繁殖,且不考虑亲本遗传差异,导致蟹苗质量差,已成为当前养殖生产发展的瓶颈问题。如苏雨等采用微卫星和线粒体标记(COⅠ、b和D-loop)分析了来自长江、黄河和辽河野生与养殖群体的遗传多样性和遗传结构,结果一致表明,三个水系群体间的遗传分化程度较低,野生群体的遗传分化与地理距离具有不完全相关性,提示不同水系的中华绒螯蟹可能与其跨流域引种及养殖群体逃逸造成其种质混杂有关。
一般认为,遗传多样性的大小及其群体遗传结构跟一个物种的进化潜力和抵御不良环境的能力密切相关。鉴于此,本研究拟在分析养殖群体遗传背景的基础上,从中选取遗传多样性高且遗传距离适度的群体作为育种基础群,采取大规格亲本进行繁育的方法培育中华绒螯蟹良种。江苏和安徽两省是目前中华绒螯蟹重要的养殖区域,成蟹养殖产量分别位列全国第一和第三,占比分别为46.3%、12.9%。本研究利用10对微卫星标记对偶数年的江苏、安徽4个主要养殖群体的遗传多样性和遗传结构进行分析,旨在为中华绒螯蟹的遗传选育、种质资源保护与利用工作提供基础资料。
1 材料与方法
1.1 样品来源
2020年8月,中华绒螯蟹样品采集于安徽省芜湖市无为县泉塘镇、江苏省南京市高淳区桠溪镇、江苏省宜兴市杨巷镇和江苏省张家港市凤凰镇魏庄村,4个养殖群体的亲本是当地养殖场于2018年底采集(或购买)的大规格野生长江蟹(雌蟹约 300 g,雄蟹约400 g)。每个养殖群体随机选取30只成蟹样品。
1.2 总基因组DNA提取
剪取上述中华绒螯蟹的附肢肌肉样品,保存于无水乙醇中备用。提取河蟹总基因组DNA,提取方法参照基因组提取试剂盒[DP324,天根生化科技(北京)有限公司]说明书进行。提取的基因组DNA分装后于-20 ℃冰箱保存备用。
1.3 微卫星位点的扩增
由表1可知,经预试验,从参考文献[7,9]中选取能够稳定扩增、等位基因个数较多、多态性较好的10对微卫星引物,由生工生物工程(上海)股份有限公司合成荧光引物。

表1 中华绒螯蟹微卫星引物信息
PCR反应总体系20.0 μL:先加入超纯水,依次补足模板DNA,10×Buffer缓冲液(TaKaRa) 2.0 μL,dNTP(2.5 mmol)1.5 μL,MgCl(2.5 mol/L)2.0 μL,上下游引物(5 μmol/L)各1.0 μL,DNA聚合酶0.3 U。PCR反应条件:96 ℃预变性3 min;94 ℃变性45 s,由表1可知,最佳退火温度下保持45 s,72 ℃延伸1 min,32个循环;循环后72 ℃延伸10 min,4 ℃结束反应,-20 ℃保存备用。扩增产物经ABI-3730XL全自动DNA测序仪进行测定,然后使用软件Genemarker 2.2读取等位基因大小进行基因分型,内标为LIZ500。
1.4 数据分析
运用软件CONVERT 1.31把基因型数据转换为各种软件格式。使用Popgene 1.32计算观测等位基因的数目、有效等位基因数目、观测杂合度、期望杂合度、近交系数及群体间Nei’s遗传距离,采用MEGA 7.0软件构建前述遗传距离的UPGMA系统树;用PIC_CALC 0.6小程序计算各位点及群体的多态信息含量。运用Arlequin 3.5软件计算成对群体之间的遗传分化系数(genetic differentiation coefficient,),并采用分子方差分析(analysis of molecular variance,AMOVA)统计变异来源。哈迪-温伯格平衡检验通过Genepop在线软件(http://genepop.curtin.edu.au/)进行。根据各位点等位基因频率,应用Bottleneck软件中的符号检验(Sign test)和符号秩次检验(Wilcoxon sign-rank),检验在3种突变模型(IAM、TPM和SMM)假设下是否偏离突变-漂移平衡(Mutation-drift equilibrium)。利用软件Strcture 2.3.4对4个养殖群体进行遗传聚类分析,确定最佳值,结果使用软件CLUMPP打开,利用小程序DISTRUCT绘图。
2 结果与分析
2.1 微卫星位点和群体遗传多样性
10对微卫星位点和4个养殖群体的遗传信息比较见表2。有效等位基因数(:4.980~26.620)均>4,观测杂合度(:0.585~0.990)、期望杂合度(:0.803~0.967)及多态信息含量(PIC:0.778~0.961)值均>0.5,符合群体遗传评估的多样度要求。

表2 微卫星位点的遗传多样性参数

表2(续)
各养殖群体均显示出较高的遗传多样性水平(=0.722~0.812,=0.904~0.918,PIC=0.874~0.891),无为泉塘群体期望杂合度及多态信息含量相对较高,宜兴杨巷群体期望杂合度及多态信息含量相对较低。综合各遗传多样性参数结果,认为各群体的遗传多样性大小顺序为:无为群体>张家港群体>高淳群体>宜兴群体。40个群体-位点的组合中,大部分组合(31个)近交系数()为正值且相应的<,所有群体的10个微卫星位点平均近交系数()值均为正。
2.2 群体遗传距离和分化
对4个养殖群体间的成对遗传距离和遗传分化指数()进行分析。由表3、表4可知,遗传距离较小(0.121 0~0.189 0),均低于0.3;群体间遗传分化指数(0.001 5~0.008 2)较小,远低于0.05,其中,遗传分化程度最大的为无为群体和宜兴群体(0.008 2),其遗传距离也最远(0.189 0)。AMOVA结果显示, 群体之间的遗传变异占总变异的比例仅为0.42%,而来自每个养殖群体内的遗传变异占比高达99.58%,表明4个中华绒螯蟹养殖群体遗传分化程度极低,无显著遗传分化。
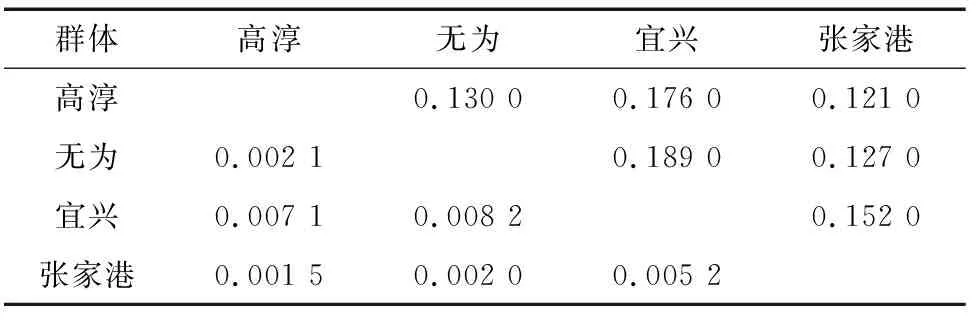
表3 中华绒螯蟹4个群体的遗传距离(对角线上)和遗传分化指数(对角线下)

表4 中华绒螯蟹4个养殖群体的分子方差分析(AMOVA)
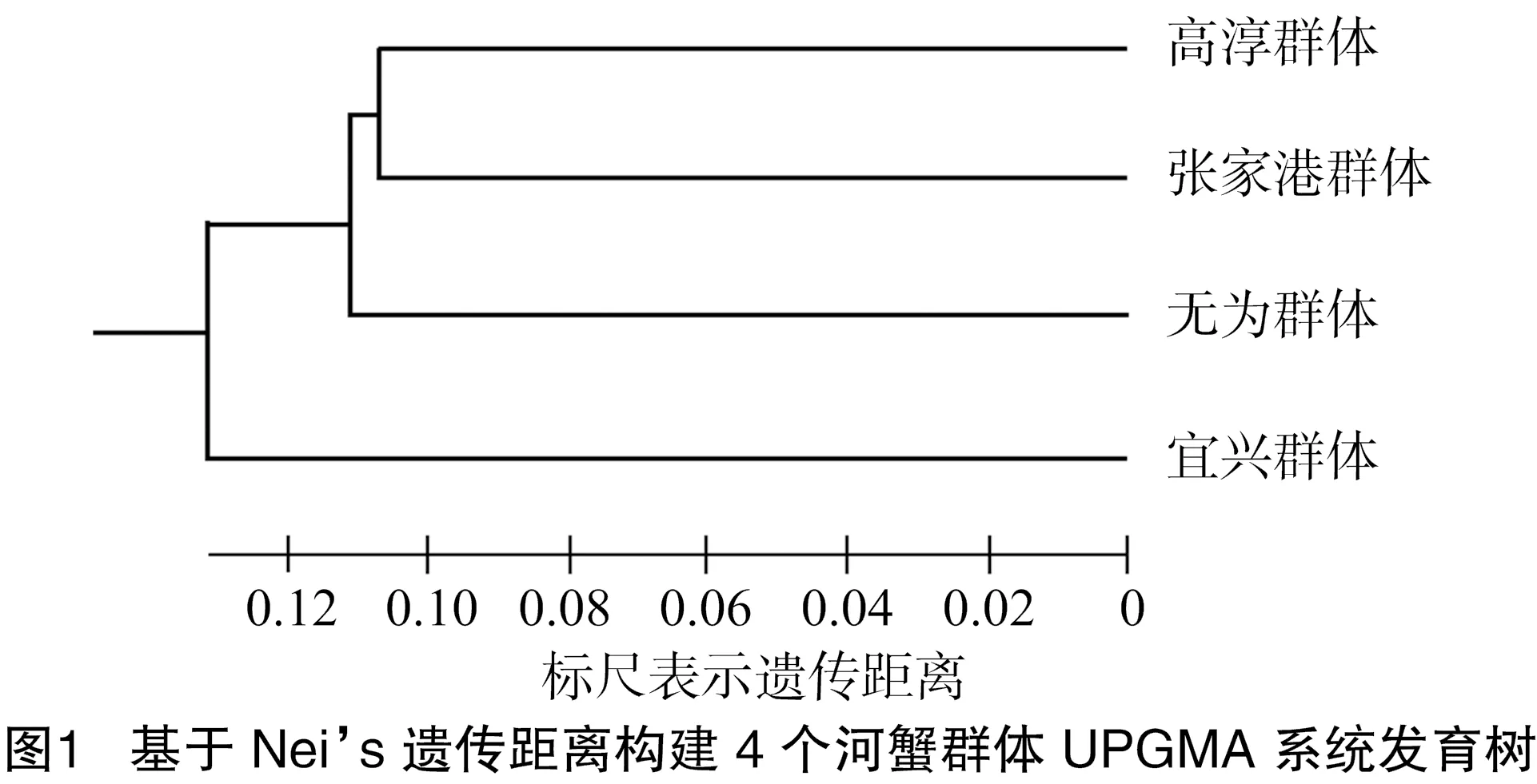
基于Nei’s遗传距离用UPGMA法构建的4个群体的系统发育树表明,4个养殖群体具有共同的祖先型,高淳和张家港群体最先聚类为一支,互为姊妹群,然后与无为群体聚类,最后与宜兴群体聚类,表明高淳和张家港群体亲缘关系最近、宜兴群体与其他群体间亲缘关系较远(图1)。
2.3 瓶颈效应分析
中华绒螯蟹4个养殖群体的突变-漂移平衡分析结果见表5。IAM假设下,Sign test显示高淳和无为2个群体偏离了突变-漂移平衡,表现出杂合显著过剩;Wilcoxon sign-rank显示所有群体均偏离了突变-漂移平衡。SMM假设下,所有群体均符合突变-漂移平衡。TPM假设下,Sign test显示所有群体均符合突变-漂移平衡;Wilcoxon sign-rank显示仅无为群体偏离突变-漂移平衡,表现为杂合过剩。

表5 中华绒螯蟹4个群体的突变-漂移平衡分析
2.4 群体遗传结构分析
设置预设聚类组数为2~5,=4时为最佳聚类分组。遗传结构图见图2,每个养殖群体均包含4个遗传聚类组,说明4个养殖群体存在相似的遗传组成,无明显的遗传结构差异。
3 讨论与结论
3.1 中华绒螯蟹的遗传多样性
遗传多样性是种质鉴定的重要部分,一般认为遗传多样性的高低与物种或群体生存和进化潜力呈呈正相关,遗传多样性越高,物种或群体适应环境变化的能力越强。而有效等位基因数、预期杂合度和多态信息含量,均是衡量遗传多样性高低的重要参数。中华绒螯蟹4个养殖群体的为9.810~11.660,为0.904~0.918,为0.874~0.891,分别高于4.0、0.5、0.5的高度多样性标准,均显示出较高的遗传多样性水平。如,高于奇数年中华绒螯蟹3个人工选育群体(“长江1号”“光合1号”和七里海群体)和1个海河自然群体的遗传多样性(为4.79~5.87,为0.720~0.745,为0.687~0.716);高于偶数年长江水系中华绒螯蟹3个育种基础群(崇明团结沙、扬中江段、靖江与六合、江浦江段)(:3.90~3.95,:0.696~0.705,:0.746~0.792);但与偶数年(2012年)辽河、黄河及长江3个水系的野生和养殖6个群体的遗传多样性结果(:13.08~14.10,:0.870~0.881,均值0.864)比较,较小、略小、值相近;大小还与长江水系野生和人工繁殖大眼幼体群体的遗传多样性水平(0.898、0.893)相近。另外,基于线粒体等其他分子标记的中华绒螯蟹研究也显示多数野生和养殖群体均具有较高的遗传多样性。综上所述,中华绒螯蟹具有高遗传多样性应该是普遍现象;本研究的中华绒螯蟹4个养殖群体遗传多样性处于高水平,具有较大的育种潜力。

3.2 中华绒螯蟹的遗传结构
群体遗传距离和遗传分化指数分别是用于衡量群体间亲缘关系、遗传分化程度的重要指标。本研究中,中华绒螯蟹成对群体间的遗传距离范围为0.121 0~0.189 0、远低于0.3的物种标准,表明这些群体亲缘关系较近。成对群体间值为0.001 5~0.008 2,均远低于0.05的遗传分化程度中等以上标准。进一步的分子变异分析(AMOVA)结果显示,群体间遗传变异占比极小(0.42%)。Structure遗传聚类分析结构显示,所有个体可分为4个遗传组,但每个养殖群体中均包含这4个遗传组,这些一致表明4个中华绒螯蟹养殖群体间遗传组成相似,群体间无显著遗传分化。
近交系数大小常用于评估群体内个体间的近交程度。本研究中,10个微卫星位点在每个群体及所有个体中的虽然均为正值(0.093~0.183、0.143)但数值较低,且40个群体-位点组合中的9个值为负值,这些结果表明,中华绒螯蟹4个养殖群体不仅均存在一定程度的近交,而且还存在少量的远缘繁殖。考虑到这些群体高度的遗传多样性水平,分析可能其苗种或亲本群体来源较为复杂。事实上,已有不少研究报道,中华绒螯蟹在我国不同地区相互引种,导致不同水系的中华绒螯蟹种质相互混杂,致使养殖群体遗传多样性偏高。
基于Nei’s遗传距离构建的4个河蟹群体UPGMA系统发育树表明,4个养殖群体具有共同的祖先型,高淳和张家港群体亲缘关系最近,其次是无为群体,最后是宜兴群体,这表明宜兴群体与其他群体间遗传距离较远。
瓶颈效应分析结果显示,3种突变模型(IAM、TPM、SMM)假设下的4个中华绒螯蟹养殖群体的突变-漂移平衡评估结果不同。由于现有研究认为微卫星数据更符合TPM模型,Wilcoxon符号秩次检验比符号检验的统计效率相对较高,并可用于4个位点以上任意样本数群体的分析。因此,在TPM假设下,符号秩次检验结果显示仅无为群体偏离了突变-漂移平衡,且表现为杂合显著过剩,这与其遗传多样性最高的结论相吻合。因此,虽然无为群体经历了瓶颈效应,但由于有远缘繁殖的引种效果,其遗传多样性依然很高。
综上所述,中华绒螯蟹4个养殖群体的遗传多样性水平高,群体间无显著遗传分化,群体间可能存在少量的种质混杂。从开发利用的角度来说,在今后的选育中应该结合采用分子遗传学、形态学等多种方法,将传统选育技术与分子辅助育种技术相结合,对长江水系中华绒螯蟹进行提纯复壮,培育长江水系中华绒螯蟹良种。
猜你喜欢
特产研究(2022年6期)2023-01-17
清华金融评论(2022年4期)2022-04-13
种子(2021年3期)2021-04-12
国际放射医学核医学杂志(2021年10期)2021-02-28
房地产导刊(2020年7期)2020-08-24
四川动物(2017年4期)2017-07-31
外语教学理论与实践(2016年1期)2016-06-11
集美大学学报(自然科学版)(2015年4期)2015-02-28
河北遥感(2014年3期)2014-07-10
中国药理学通报(2014年2期)2014-05-09
