
外源钙对干旱胁迫下不同抗性牡丹PSⅡ功能和光能分配的影响
2022-08-26 07:02:12李海霞米银法陈双臣
江苏农业科学 2022年16期
李海霞, 米银法, 陈双臣
(1.河南财政金融学院,河南郑州 450000; 2.河南科技大学园艺与植物保护学院,河南洛阳 471003)
土壤干旱是限制农业可持续发展的重要环境因素,干旱常常引起植物大量缺水、正常光合生理代谢紊乱,如气孔闭合,酶活性降解,光合同化力下降,光能过剩活性氧增加,损害光合机构。但许多研究表明,干旱胁迫时植物可启动热耗散调节途径,依靠自身生理调节机能降低过剩光能的伤害,保护光合机构。如干旱下,白及、增加,降低,光合机构受损;甜高粱净光合速率、气孔导度和蒸腾速率降低,升高,、/、逐渐降低,表明高粱幼苗受损加重;干旱时棉花净光合速率和气孔导度降低,叶绿素荧光指标荧光产量(Yield)和叶绿素荧光非光化猝灭(NPQ)比/对干旱更敏感。鼓节竹干旱胁迫时可通过降低、和增加来降低 PSⅡ反应中心的开放程度,从而降低PSⅡ干旱下的伤害,表现出一定的耐旱性。研究还表明不同抗旱型的植物品种应对胁迫下PSⅡ受损时的保护机制不同。如遭受干旱时,耐旱型辣椒品种正椒13号的/、、和NPQ显著高于敏感型品种弄口早椒。说明辣椒品种的抗高温干旱能力与较高的光化学能力和热耗散能力相关。
Ca是叶绿体PSⅡ必不可缺的重要组分,它在维持PSⅡ中心活性和稳定细胞膜结构方面发挥着重要作用。研究表明,干旱、高温等逆境时外源Ca能够调控PSⅡ中心活性,优化逆境下的叶绿素荧光能量分配,改善植物的光合性能。如干旱胁迫下施用外源钙后甘草叶片叶绿素含量、净光合速率、气孔导度、胞间CO浓度均有不同程度提高。高温胁迫下喷施CaCl后葡萄叶n显著提高,维持了较高的PSⅡ活性,、′/′、/极显著高于对照,极显著小于对照,有效缓解了高温对葡萄叶光合作用的抑制及PSⅡ系统的损害。高盐胁迫下外源Ca能显著提高巴西蕉幼叶/、光化学淬灭、和,增强光合活性、光保护能力和捕光能力,有效缓解高盐对光合系统的不利影响。干旱胁迫下喷施CaCl后烤烟叶片光合电子传递能量比例()降幅明显小于对照,电子转运效率明显高于对照,PSⅡ用于光合电子传递的捕获光能比例增加,提高了烤烟幼苗的抗旱能力。
综上所述,关于外源Ca对干旱胁迫下植物叶绿素PSⅡ功能的研究文献相对较多,但有关外源Ca对干旱胁迫下牡丹(Andr.)叶绿体PSⅡ荧光活性的影响,尤其是PSⅡ功能及吸收光能分配方面的研究尚未见报道,为此本研究以不同抗旱能力的牡丹品种为试材,研究干旱胁迫下外源CaCl对其PSⅡ功能及叶片吸收光能分配的影响,以期探求Ca的调控生理机制,为牡丹的栽培开发及引种管理等提供理论基础和依据。
1 材料与方法
1.1 试验材料
试验于2018年8月至2019年6月在河南省林业科学研究院进行。选抗干旱能力较强的胡红和不耐干旱胁迫的乌龙捧盛牡丹三年生分株苗为试材,植株由中国洛阳牡丹国家基因库提供。2018年8月初选择长势良好、均匀一致的优良苗,统一修整后在内径30 cm高28 cm的瓦盆中进行盆栽,1盆1株,基质为大田肥沃土壤。每种牡丹栽种12棵,正常肥水管理。 2019年6月1—10日将盆栽分为不喷施CaCl的对照组和外施CaCl的处理组。每个处理6盆,6次重复。处理组喷施 0.3 mmol/L CaCl,对照组喷施蒸馏水。喷施时按早晚间隔 8 h以上的原则,每天对牡丹叶片正反面喷洒2次,叶片喷洒直至有水滴落,连续10 d。6月1—10日期间上述材料一边进行喷施试验,一边进行下列干旱胁迫试验,胁迫期间搭建防雨棚遮雨备用。6月1—10日内按文献[13]每天傍晚时分对盆栽进行称重、及时补水,使盆栽维持80%土壤含水量,6月11日后进行控水处理,让各盆栽自然干旱。测得3、6、9 d时的相对含水量分别为50%~55%(轻度干旱)、35%~40%(中度干旱)、25%~30%(重度干旱)。干旱控水10 d,各处理大部分叶片明显萎蔫,此时浇水至80%的相对土壤含水量进行复水并控水2 d。
1.2 试验方法
分别在试验期内0、3、6、9、12 d 10:00—12:00用美国Li-Cor6400XT V6.1荧光仪测定各处理叶绿素荧光参数,每次测定时固定选用基部上数第 4~5节位的完全展开叶。按照仪器操作说明并参照周艳虹等的方法测定各荧光指标,包括暗适应时叶片最大荧光()、初始荧光()、光下叶片初始荧光(′)、可变荧光()、最大荧光(′)、表观光合电子传递速率()、最大光化学效率(/)、实际光能转换效率(′/′)、非光化学淬灭系数()和叶绿素荧光光化学猝灭系数()等,指标可由仪器直接读出。各指标重复4次。叶片天线热耗散百分率()、非光化学耗散百分率()、光化学反应百分率()分别按照下列公式进行计算。试验数据用DPS软件进行分析,用Excel软件进行作图。
=1-′/′;
=×′/′;
=(1-)×′/′。
2 结果与分析
2.1 Fo、Fm的变化
反应中心PSⅡ开放时的荧光水平通常由叶绿素基础荧光()加以衡量。图1表明,抗性不同的牡丹品种胡红和乌龙捧盛遭受干旱胁迫时随胁迫程度的加重,对照及处理叶绿素基础荧光均整体呈逐渐升高趋势,趋势则整体逐渐降低。9 d时,重度干旱胁迫下值最大,胡红、乌龙捧盛分别比对照降低19.6%、14.1%(<0.05)。说明干旱胁迫对牡丹叶片光合作用产生了明显的光抑制现象,光合原初反应受到抑制,影响了植株正常生长。但与胁迫初期相比,耐性较强的胡红对照增加了48.1%,处理增加了25.9%。可见外施CaCl后耐性较强的胡红增幅减少了22.2百分点。耐性较差的乌龙捧盛对照比胁迫初期增加了64.5%;处理比胁迫初期增加了45.0%。因此外施CaCl后耐性较差的乌龙捧盛增幅被抑制了19.5百分点。可见遭受干旱胁迫时外施CaCl后,2个品种的牡丹增幅均受到了显著抑制,缓解了因干旱逆境对2种牡丹叶绿素PSⅡ反应中心产生的破坏作用,减轻了失活。同时说明CaCl对耐性较强的胡红缓解光抑制效应,改善光合作用的能力明显优于耐性较差的乌龙捧盛。图1还表明,胁迫后期经CaCl处理的2个品种复水后叶片显著低于重度干旱时,与中度胁迫时无显著差异,可见复水一定程度地缓解了2个牡丹品种的光抑制,可恢复到中度干旱时的水平。

研究认为逆境胁迫时,最大荧光()降低是光抑制光合作用的另一显著表现。图1表明,随胁迫程度的加重2个牡丹品种逐渐降低,重度胁迫时最低,此时与胁迫初期相比,胡红对照及处理分别降低了19.5%、14.6%;乌龙捧盛则分别降低了22.4%、13.9%。可见外施CaCl后2个品种均显著抑制了胁迫期内的减少,就品种差别而言胡红减少程度相对较轻。外施CaCl复水后,胡红恢复到了轻度干旱时的水平(>0.05);乌龙捧盛则恢复到了中度干旱时的水平。因此CaCl抑制胡红降低的效果优于乌龙捧盛。
2.2 Fv和Fv/Fm的变化
逆境生理下叶绿素可变荧光的大小及变化,可用于反映PSⅡ最初电子受体质体醌Q的氧化还原情况。由图2可见,2种牡丹叶片随干旱胁迫程度的增加逐渐降低,直到后期复水后,外施CaCl的2种牡丹叶又回升至中度干旱时的水平(>0.05)。说明外施CaCl结合复水能够显著提高2个牡丹品种叶片光化学效率,改善光合性能从而获得较多的能量物质,以应对干旱逆境带来的伤害。图2还表明,外施CaCl后2个牡丹品种虽总体趋势降低,但同一干旱程度下,胡红、乌龙捧盛均显著高于各自的对照。9 d时,胡红、乌龙捧盛分别比对照提高了37.5%、34.7%。因此2种牡丹遭受干旱逆境时,光反应中心最初电子受体Q的氧化还原能力显著降低。但外施CaCl后2种牡丹显著提高,说明CaCl对Q的氧化还原能力起到了有效的保护作用。外施CaCl后6~9 d时,耐干旱能力较强的胡红品种显著高于抗性较差的乌龙捧盛(<0.05),品种间差异显著。2个品种之所以存在这种差异,可能与CaCl能维持胡红较低的和较高的(图1)有关。
叶绿素光能转化时,/的大小和光合电子传递活性成正比,可用于衡量原初光能转化率的高低。图2表明,随干旱胁迫程度的加重,抗性不同的2个牡丹品种叶绿素/均逐渐降低。但中度及重度干旱时喷施CaCl,2个牡丹品种叶片/都显著高于对照,其中胡红和乌龙捧盛分别比对照提高了10.7%~20.0%、9.7%~20.1%。可见遭受中度和重度干旱时,喷施CaCl能有效缓解牡丹叶绿素PSⅡ系统原初光能转化效率的受损程度,维持暗适应时较高状态的最大光化学效率。

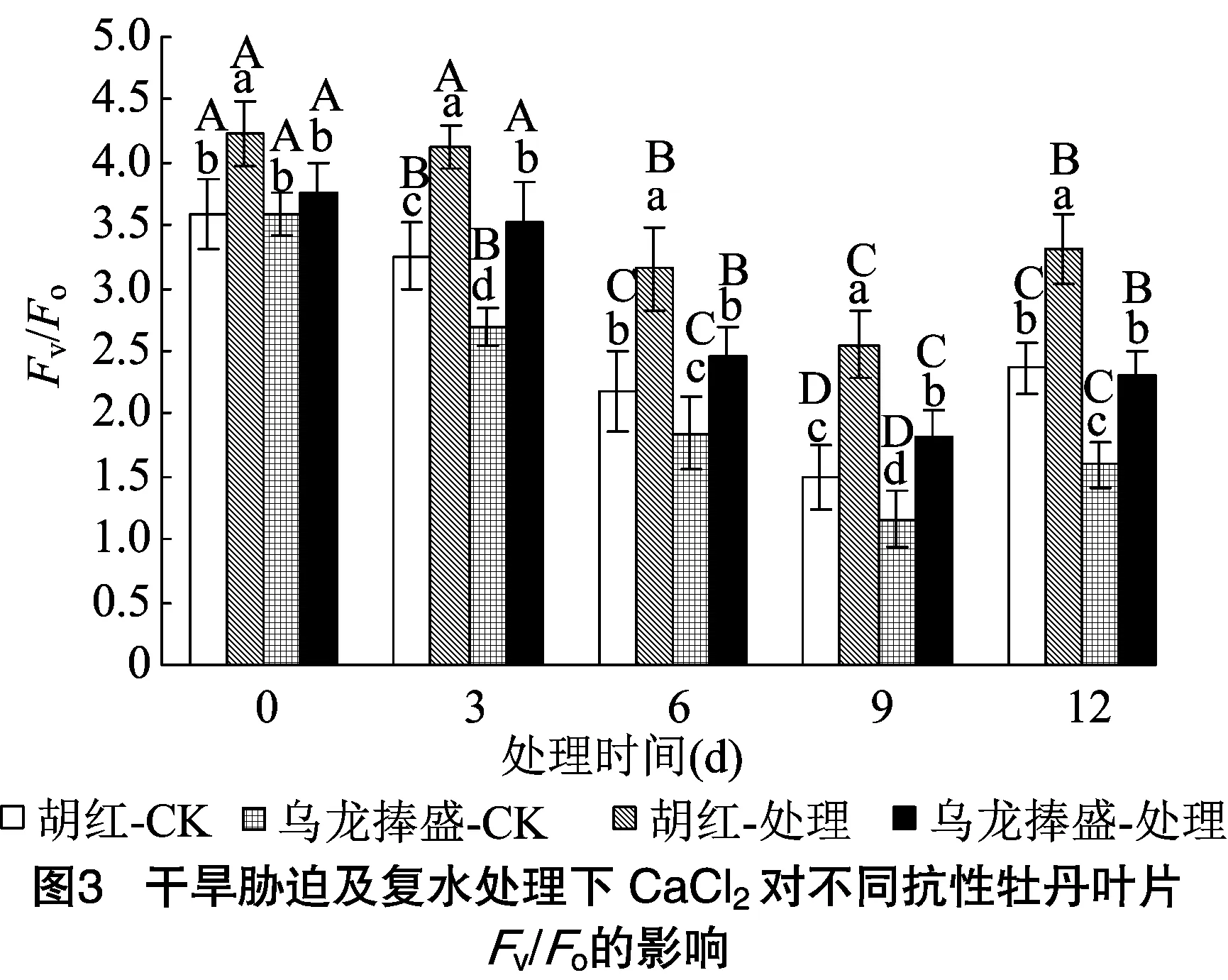
2.3 PSⅡ光潜在活性Fv/Fo的变化
叶绿体光反应中心PSⅡ的潜在活性可用/表示。图3表明,对照及处理条件下的2种牡丹,叶片/都随干旱胁迫程度的增加呈下降趋势。2种牡丹对照叶片内/范围为1.5~3.6,外施CaCl后则维持在1.8~4.2之间。可见干旱胁迫逆境下2种牡丹叶片PSⅡ潜在活性均受到了明显抑制,但外施CaCl可以显著提高/,进而提高PSⅡ的潜在活性。进一步方差分析表明,干旱胁迫期间3~12 d,处理和对照之间以及外施CaCl后的2个品种之间,/均存在显著差异(<0.05)。尤其是重度干旱9 d时,胡红和乌龙捧盛分别比对照值提高了71.5%、56.9%。表明此时抗性较强的胡红PSⅡ潜在活性显著高于乌龙捧盛(<0.05)。12 d经复水处理后,2种牡丹/又迅速回升且与6 d时差异不显著,可见PSⅡ潜在活性经过3 d复水已经能够恢复到中度胁迫时的水平。
2.4 Fo′、Fm′、Fv′及Fv′/Fm′的变化
光下初始荧光(′)、光下最大荧光(′)、光下可变荧光(′)和PSⅡ实际光能转换效率(′/′)也是叶绿素荧光参数研究中的几个重要指标。图4表明,遭受干旱胁迫时,抗旱能力不同的2种牡丹随胁迫程度的增加,叶内′逐渐升高,′、′及′/′整体逐渐降低。表明遭受干旱胁迫时,胡红和乌龙捧盛 PSⅡ反应中心捕获激发能效率下降,PSⅡ 功能受到影响,致使牡丹植株叶片PSⅡ反应中心光化学效率降低。与各自对照相比,2种牡丹外施CaCl后′显著降低,′、′及′/′显著提高。外施CaCl后9 d的胡红、乌龙捧盛′/′分别为0.71、0.66,分别比对照提高了20.3%、17.4%。由此可见,外施CaCl后抗旱能力较强的胡红′/′无论是数值大小还是增加幅度,都显著高于耐旱能力较差的乌龙捧盛(<0.05)。表明外施CaCl可以显著提高干旱胁迫尤其是重度胁迫下牡丹植株PSⅡ的实际光能转换效率,而且抗旱能力较强的胡红PSⅡ实际光能转换效率提高效果显著优于耐性较差的乌龙捧盛。胁迫后期复水后2种牡丹的′/′又逐渐升高,表明PSⅡ实际光能转换效率又得以恢复。

2.5 qP、qN的变化
叶绿素荧光光能包含光化学猝灭()和非光化学猝灭()2种主要的能量耗散形式。图5表明,遭受干旱胁迫时,抗旱能力不同的2种牡丹随干旱时间的延长胁迫程度加剧,叶内耗散逐渐降低、耗散逐渐增加,复水时和得到改善。干旱胁迫下的减少和的增加意味着PSⅡ电子传递活性减弱、非光化学猝灭能量耗散增加,这2种因素造成Q的重新氧化能力减弱,植株光合碳同化能力受到严重影响。但与对照相比, 外施CaCl

显著提高了胡红和乌龙捧盛中度和重度干旱胁迫下的值,显著降低了重度干旱胁迫下的值(图5)。9 d时胡红处理和乌龙捧盛处理的分别为0.53、0.46,比各自的对照提高了23.3%、24.3%;分别为0.75、0.87,比各自的对照降低了17.9%、28.9%。因此2个牡丹品种外施CaCl可以通过提高降低的方式,进一步稳定PSⅡ的电子传递活性,保持它们较高的Q重新氧化能力。就2个牡丹品种而言,遭受重度干旱时抗性较强的胡红仍能保持相对较高的值和相对较低的值,说明胡红的能量耗散比乌龙捧盛损失较轻,维护光合反应中心免受破坏的能力相对更强,2个品种间差异显著(<0.05)。
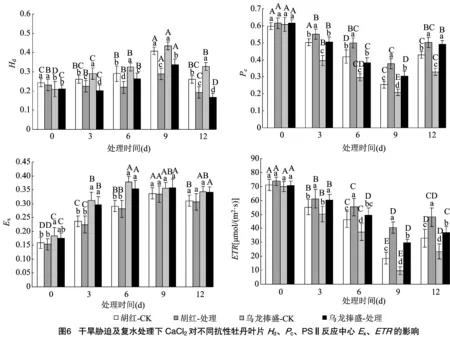
2.6 Pc、Hd、PSⅡ反应中心Ex、ETR的变化
由图6可见,随胁迫程度的加剧,胡红和乌龙捧盛及反应中心呈逐渐升高的趋势,和逐渐降低(<0.05)。但与各自的对照相比,外施CaCl条件下2个牡丹品种除外,显著降低,、均显著提高。9 d时胡红和乌龙捧盛分别比对照降低29.3%、22.5%,增加48.3%、45.9%,增加118.3%、208.6%。因此CaCl处理能够显著提高干旱胁迫下2个牡丹品种叶绿素PSⅡ的光化学反应能量百分率和表观光合电子传递速率,CaCl的这种调控效果非常有利于PSⅡ光化学效率的升高,结论与王晓黎等的研究结果一致。CaCl的这种调控作用可能与黄素循环受到抑制或D1蛋白活性升高相关。这也是经外源CaCl调控下牡丹自行适应干旱胁迫的应对调节机制之一。就2种牡丹品种差异而言,虽然CaCl调控下胡红的表观光合电子传递速率增幅低于乌龙捧盛,但其光化学反应的能量百分率增加的幅度及天线热耗散的百分率减少的幅度,都好于耐旱性较差的乌龙捧盛。
2.7 叶绿素荧光能量分配的变化
由图7可见,遭受干旱胁迫0~6 d时,与抗性较差的乌龙捧盛相比,胡红PSⅡ反应中心用于叶绿素荧光能量分配的吸收光能各份额所占比例较为稳定,能量分配基本没有受到较大影响。但总体上胁迫期内,2种牡丹PSⅡ吸收光能用于叶绿素荧光能量分配的和所占份额整体逐渐加大、所占份额逐渐减少,直到复水时、有所降低、增加。9 d时,胡红对照∶∶=33.7 ∶25.4 ∶40.9,乌龙捧盛∶∶=35.5 ∶20.9 ∶43.6。外施CaCl后9 d,胡红∶∶=33.4 ∶37.7 ∶28.9、乌龙捧盛∶∶=35.8 ∶30.5 ∶33.7。9 d时,胡红、乌龙捧盛分别比对照增加了48.3%、45.9%,降低了29.3%、22.6%。胁迫中后期比值的增大,说明牡丹叶绿体捕捉到的光能分配比例进入PSⅡ反应中心的能量减少,天线热耗散的能量损失份额加大,叶片光合保护机制严重受损。经外施CaCl调控后减少、增大,可见外施Ca能够提高进入PSⅡ反应中心的能量,缓解并降低天线热耗散的能量损失,提高PSⅡ反应中心的光化学反应百分率,起到了很好地保护牡丹叶片光合机制的作用。

3 讨论与结论
在正常生长条件下,植物叶绿体中光合反应中心PSⅡ活性较强,能够及时将捕获天线收集到的光能用于光化学反应,实现电子传递藕连光合磷酸化助推碳同化并生成同化力。但遭受逆境胁迫后,PSⅡ 光化学反应中心将发生可逆性失活,即便能吸收光能也推动不了电子传递,只有等逆境解除后反应中心活性才能恢复。其中Ca在PSⅡ调控中起着重要作用, 譬如Ca通道可抑制叶绿素荧光的猝灭。Ca还可参与PSⅡ放氧系统中的重要氧化还原反应,外施Ca进入细胞质后,会对叶绿体内的酶代谢过程进行调控,譬如碳同化相关Rubp竣化酶的活性等。目前认为干旱胁迫条件下外施Ca的生理调控作用,主要体现在它能维持类囊体膜的稳定性,并显著增强类囊体膜上的电子传递能力,提高叶片光能的转换效率。Ca还担负着细胞第二信使的功能,当植物感受到发育或生物、非生物胁迫信号的刺激后,Ca泵和Ca通道得以激发诱导细胞内Ca浓度变化产生钙信号。借助于Ca-CaM信使系统参与并保护细胞膜双脂层结构的稳定和完整性。Ca还能够结合许多受体细胞,接收外界环境信号并进行传递和放大,触发一系列增强植物抗逆特性相关的生理反应,如内源渗透性调节物质的增加、保护酶活性的增强等。为此本研究以CaCl为外源调控物质,以便探求Ca对不同抗性牡丹品种干旱胁迫下的叶PSⅡ功能及叶绿素荧光能量分配的影响。
本研究中2种不同抗性的牡丹叶片随干旱胁迫程度的加深,逐渐升高、逐渐降低(图1),致使显著降低(=-),进而/(图2)和/(图3)也显著下降。的逐渐升高说明2种不同抗性的牡丹叶片PSⅡ 反应中心光化学已受到了明显的损害。同时,研究认为植物叶片PSⅡ 反应中心遭受逆境时,还可以通过增加热耗散来降低逆境伤害,并伴随另一种猝灭过程,这个猝灭过程可由叶绿素荧光光化学猝灭系数进行衡量,且越大电子传递活性越强,氧化形成的QA量越多。外施CaCl后与对照相比,2个牡丹品种均显著抑制了各胁迫程度时的增幅和的减少,提高了以及/,证明外施CaCl能够有效减轻牡丹叶绿素光反应中心PSⅡ系统中原初光能转化效率受损程度,维持暗适应时较高状态的最大光化学效率。与各自的对照相比,外施CaCl后2种牡丹内′显著降低,′、′及′/′显著提高。′及′/′即光下可变荧光和PSⅡ实际光能转换效率,表明CaCl可以显著提高干旱胁迫尤其是重度胁迫下牡丹植株内PSⅡ的实际光能转换效率,维持较高的碳同化产物。
图5表明遭受干旱胁迫时,2种牡丹随时间的延长叶内逐渐降低、逐渐增加。逐渐降低说明叶绿素PSⅡ氧化反应中心的电子传递Q-Q的过程中,受到了较为严重的抑制,最终导致进入并用于光合作用的电子量减少,同时也提高了耗散掉的无效光能份额,这和本试验中升高的结论吻合(图6)。但外施CaCl后2个牡丹品种又显著提高、显著降低,表明CaCl可以通过调控干旱胁迫时的PSⅡ氧化侧向反应中心Q-Q的传递过程,改善光合电子传递效率,利于提高牡丹叶片NADPH和ATP的合成能力。
2个牡丹品种遭受干旱尤其是重度胁迫时,外施CaCl后对干旱敏感的乌龙捧盛/、/、′/′、无论是增加幅度还是数值大小均显著低于抗性较强的胡红,同时的降低幅度也高于胡红。因而抗性较强的胡红较高的和′/′增幅及数值,维持了其胁迫下较好的叶片PSⅡ反应中心开放程度和较高的光合电子传递速率。即为非辐射能量耗散诱发的荧光猝灭,外施CaCl后胡红的降幅显著低于乌龙捧盛,说明外施CaCl后胡红用于保护光合机构而消耗的非辐射能量相应少于乌龙捧盛。由此可见就2个牡丹品种来说遭受重度干旱时,外施CaCl能促使抗性较强的胡红调控至相对较高的值和相对较低的值状态(<0.05),保持较少的能量耗散和较强的ATP、NADPH合成力,维护光合反应中心免受破坏的能力更强,损失显著轻于乌龙捧盛。
外施CaCl后与各自对照相比,2个牡丹品种除差异不显著外,显著降低,、均显著提高(图6)。减少、增大,表明外施CaCl能够减少天线热耗散的能量损耗,使得进入PSⅡ反应中心的能量增多,提高PSⅡ反应中心的光化学反应百分率,进而促进光合磷酸化以及NADPH和ATP的生成,以便更好地保护牡丹叶片光合机制向暗反应配送更多的还原力。这与低温弱光下黄瓜光反应中心的能量分配结论类似,其作用机理还与黄素循环受到抑制或D1蛋白活性升高相关。而没有施加CaCl的对照牡丹叶内,在胁迫中后期份额增大,说明重度胁迫加大了天线热耗散,从而抑制了PSⅡ反应中心能量的进入,2个牡丹品种叶片光合保护机制受损严重。文献表明引起增加的因素,主要有以下几种:(1)干旱胁迫时叶内PSⅡ反应中心接受源于捕光色素捕获能量的距离增加,加大了“线耗”,也随之增高;(2)的增减还密切关联于叶黄素的循环过程和光合单位叶绿素分子数量的多寡;(3)逆境条件下植物中的减少也会促进份额的增加。
就2个牡丹品种PSⅡ反应中心光能分配差异而言,干旱胁迫中后期(6 d后),外施CaCl后不耐干旱的乌龙捧盛和所占份额均显著高于抗性较强的胡红(图7),但是较高份额也会引起 PSⅡ 反应中心激发压的迅速增大,这种状况将会直接导致叶绿素分子处于能量过剩状态,过剩的能量一旦传给O,将会形成极具破坏性的单线态O。因此乌龙捧盛较高的份额引起的光合机构失活程度和破坏作用明显高于胡红。可见干旱逆境时,Ca对抗性较弱的乌龙捧盛PSⅡ叶绿素荧光能量分配的调控机制要劣势于抗性较强的胡红。总之这种比较复杂的Ca调控机制有待于进一步探讨。
猜你喜欢
小哥白尼(军事科学)(2020年12期)2021-01-18 06:10:02
阅读(科学探秘)(2020年8期)2020-11-06 06:22:48
学生天地(2020年2期)2020-08-25 09:03:00
小哥白尼(军事科学)(2020年1期)2020-06-16 03:10:26
中国果业信息(2019年1期)2019-01-05 17:41:42
现代园艺(2017年13期)2018-01-19 02:28:13
生物学教学(2017年9期)2017-08-20 13:22:32
连环画报(2017年1期)2017-07-13 09:01:55
北方音乐(2016年12期)2016-08-23 03:20:03
太空探索(2016年2期)2016-07-12 09:57:46
