
辣椒品质研究进展
2022-08-26 07:48:48王楠艺付文婷涂祥敏杨万荣何建文
江苏农业科学 2022年16期
王楠艺, 付文婷, 吴 迪, 涂祥敏, 杨万荣, 何建文
[贵州省农业科学院蚕业(辣椒)研究所,贵州贵阳550006]
辣椒原产于南美洲,经中美洲和欧洲,再到非洲和亚洲,现在全球范围内栽培种植,中国是鲜辣椒产量最高的国家,世界上干辣椒产量最高的是印度。早在1万年前辣椒被人类熟知并作为食物,既可鲜食也可干食,辣椒富含多种重要的营养成分,如辣椒素类、辣椒素酯类、维生素、类胡萝卜素、酚类化合物、各种微量元素等。其中,辣椒所特有的辣椒素类物质具有降糖、抗肿瘤、抗疲劳、肝损伤保护等多种作用。辣椒维生素C含量是柑橘类水果的2倍。干红辣椒富含维生素A,并且是-胡萝卜素的重要来源。它们有助于人体健康,如减少肥胖和糖尿病。辣椒具有抗菌特性,含有生物类黄酮等最常见的抗氧化剂。辣椒还可以通过门冬酰胺酶有效预防癌症。这些营养成分对人体健康非常重要,决定着辣椒的营养品质。优良的营养品质有助于提升商品价值,相应地有利于种植推广,因而国内外都十分注重辣椒的营养品质研究,并取得了长足的进展。本文对辣椒营养品质的国内外研究进展进行概述,旨在为深入研究和进一步提高辣椒营养品质提供参考。
1 辣椒营养品质类型及功能
1.1 辣椒素类和辣椒素酯类
辣椒素类物质是一类生物碱化合物,为辣椒属植物所特有,是7个同源的支链烷基香草酰胺的混合物。Thresh于1876年首次分离出结晶形式的化合物,并将其命名为“辣椒素类”。辣椒素类物质包括5种:辣椒素(CAP)、二氢辣椒素(DHCAP)、降二氢辣椒素、高二氢辣椒素和高辣椒素,辣椒素是最重要的一种。辣椒素和二氢辣椒素含量最丰富,占总辣椒素的近90%,二者的差别在于支链脂肪酸的碳9处的不饱和双键。辣椒素决定辣椒辣味的强弱,国际上习惯用史高维尔热量单位(Scoville Heat Unit,简称SHU)表示辣度,SHU数值与辣椒辣度呈正相关。辣椒素酯类物质是在一种甜椒品种中被首次发现,其结构与辣椒素相似,存在于大部分辣椒种质资源中。基于脂肪酸部分的结构,辣椒素酯类物质包括辣椒素酯和二氢辣椒素酯3种。辣椒素和辣椒素酯类物质表现出相似的生理活性,包括增加耗氧量、提高体温以及抑制脂肪积累。辣椒素酯之所以无刺激性,是因为它是由香草醇和支链脂肪酸缩合而成的,具有高亲脂性或水不稳定性,这使得感觉神经元难以接触到它。
1.2 类胡萝卜素
类胡萝卜素属于四萜类化学物质,它们的显色特性使生物体呈现出橙色、黄色和红色。类胡萝卜素对营养和健康至关重要。它们具有抗氧化和抗癌特性,并且是维生素A和视黄酸等其他关键物质的前体。依据化学结构的不同,类胡萝卜素可以分为胡萝卜素和叶黄素2类;基于显色特性可以分为黄色和红色2个色系。辣椒果实具有绿色、黄色、橙色和红色多种鲜艳的颜色,都是源自类胡萝卜素色素,存在于果实成熟过程中形成的有色体的类囊体膜中。其中,叶绿体a和叶绿体b呈现绿色;-胡萝卜素、-隐黄质、玉米黄质、花药黄质、紫黄质和叶黄素呈现黄橙色;辣椒红素、辣椒玉红素和隐辣椒素呈现红色,并且是辣椒果实中所特有的。辣椒果实呈现红色正是由于高含量的辣椒红素和辣椒玉红素所致,而呈现橙黄色果实的辣椒是因为含有-胡萝卜素和紫黄质。类胡萝卜素中辣椒红素占6成。辣椒红素和辣椒玉红素的积累速率随着成熟的后期呈指数增加。
1.3 维生素
辣椒富含维生素A、维生素C、维生素E和叶酸,有利于减少人体患多种癌症和心脏病的风险。辣椒被证明是维生素C的丰富来源。从绿色阶段到红色肉质阶段,辣椒果实可以提供足够的维生素C以满足成人60 mg/d的建议摄取量。辣椒富含维生素E(生育酚),不同品种含量差异显著,变幅从3.7~236 mg/100 g不等,红辣椒粉的-生育酚水平与菠菜和芦笋相当,其含量是西红柿的4倍。100克红色辣椒将超过普通成年人的营养素供给量(RDA)(8~10 mg)。包括-胡萝卜素和-隐黄质在内的类胡萝卜素在细胞内代谢为视黄醇和视黄酸,这是维生素A的活性形式,与年龄相关性黄斑变性(AMD)和肺癌的风险降低有关。
1.4 酚类化合物
酚类化合物是广泛存在于植物中的次生代谢物。它们作为一种防御机制,有助于通过苯丙烷途径在胁迫期间维持植物的生理过程。酚酸衍生物和黄酮类化合物是辣椒果实中主要的酚类化合物,它们作为抗氧化剂具有促进健康的功能。几项流行病学研究表明,食用类黄酮和酚酸与降低患神经退行性疾病和冠状动脉疾病、骨质疏松症、癌症和糖尿病的风险之间存在潜在联系。除了黄酮醇苷外,辣椒果实的果皮还含有一些芹菜素、杨梅素、木樨草素和山柰酚衍生物的苷和苷元。黄酮醇与抗真菌、抗菌、抗氧化和抗癌特性有关,因为在特定位置存在羟基和在C2~C3位置存在双键,如在辣椒中含有槲皮素3----鼠李糖苷。青椒的未成熟果实比成熟果实含有更高的黄酮醇(高达4.5倍)。红色和紫色的辣椒含有花青素,成熟的黄色果实和成熟的红色果实中的总花青素水平在 0.5~28 mg/100 g不等。飞燕草素-3-反式-香豆酰芸香苷-5-葡萄糖苷是在辣椒果实中发现的最丰富的花青素。
1.5 辣椒营养品质的检测
对于辣椒素成分的分离,一般采用薄层色谱法、超临界流体色谱法、气相色谱法和液相色谱法(LC)。在这些方法中,LC因其快速、可靠、准确和精确的特性而被广泛使用。用于辣椒素测定的最新技术可以采用LC与质谱联用或者超高效LC,因为它分析速度快,并且在流动相下溶剂消耗量减少了5~10倍。Douventzidis等利用Weber-Fechner定律来说明更复杂的辣椒量表,以替代传统使用的史高维尔热量单位,新的评级量表简单有效。由美国香料贸易协会(ASTA)制定的使用分光光度法对辣椒的可提取颜色值进行量化。ASTA参考值也可以通过美国官方分析化学师协会制定的方法评估。
2 辣椒营养品质育种现状
辣椒最早起源于美洲热带地区,有30多个种,其中只有4个栽培种,包括辣椒(L.)、黄灯笼辣椒(Jacq.)、风铃辣椒(L.)和绒毛辣椒(L.)。虽然驯化品种来源于热带品种,但大多在温带国家进行辣椒育种,研究人员最关注的是辣椒(L.)。辣椒属的第1批育种者是美洲的土著人,他们通过选择驯化辣椒品种。考虑到对营养品质的多样化需求,各国在世界范围内进行了种质资源探索和评估,以确定富含辣椒素类和类胡萝卜素的种质、育种系或地方品种。世界各地的辣椒育种者都试图评估和鉴定更有辛辣感的辣椒,这促使了对辛辣辣椒种质资源的收集。世界各国先后培育出很多不同辣度的辣椒品种,在不同地区推广种植(表1)。印度东北部辣椒以辛辣度高而闻名,印度东北部的Bhoot Jolokia,来源于和之间的自然杂交,因其超过百万SHU,注册了吉尼斯世界记录。然而,在几年之后,由澳大利亚的Butch Taylor培育的Trinidad Butch T Scorpion和由Bosland等鉴定的Trinidad Moruga scorpion,辣度超过了Bhoot Jolokia。2013年,该纪录再次被Carolina Reaper打破,该品种具有220万SHU的辣度。2017年,一种名为Pepper X的新辣椒培育成功,其斯科维尔评分为318万SHU。目前已经确定了许多富含类胡萝卜素的种质资源(表1),例如基因型ICBD-10、EC-18和ICBD-8含有大量类胡萝卜素和较低的辣度(>200ASTA 颜色值)。Kt-Pl-19、PBC171、SSP-1999和ICBD-8/10也具有较高的ASTA颜色值。Bydagi Dabbi以其深红色(150~200 ASTA颜色值)和低辣度而享誉国际,于2010年9月获得地理标志证书。
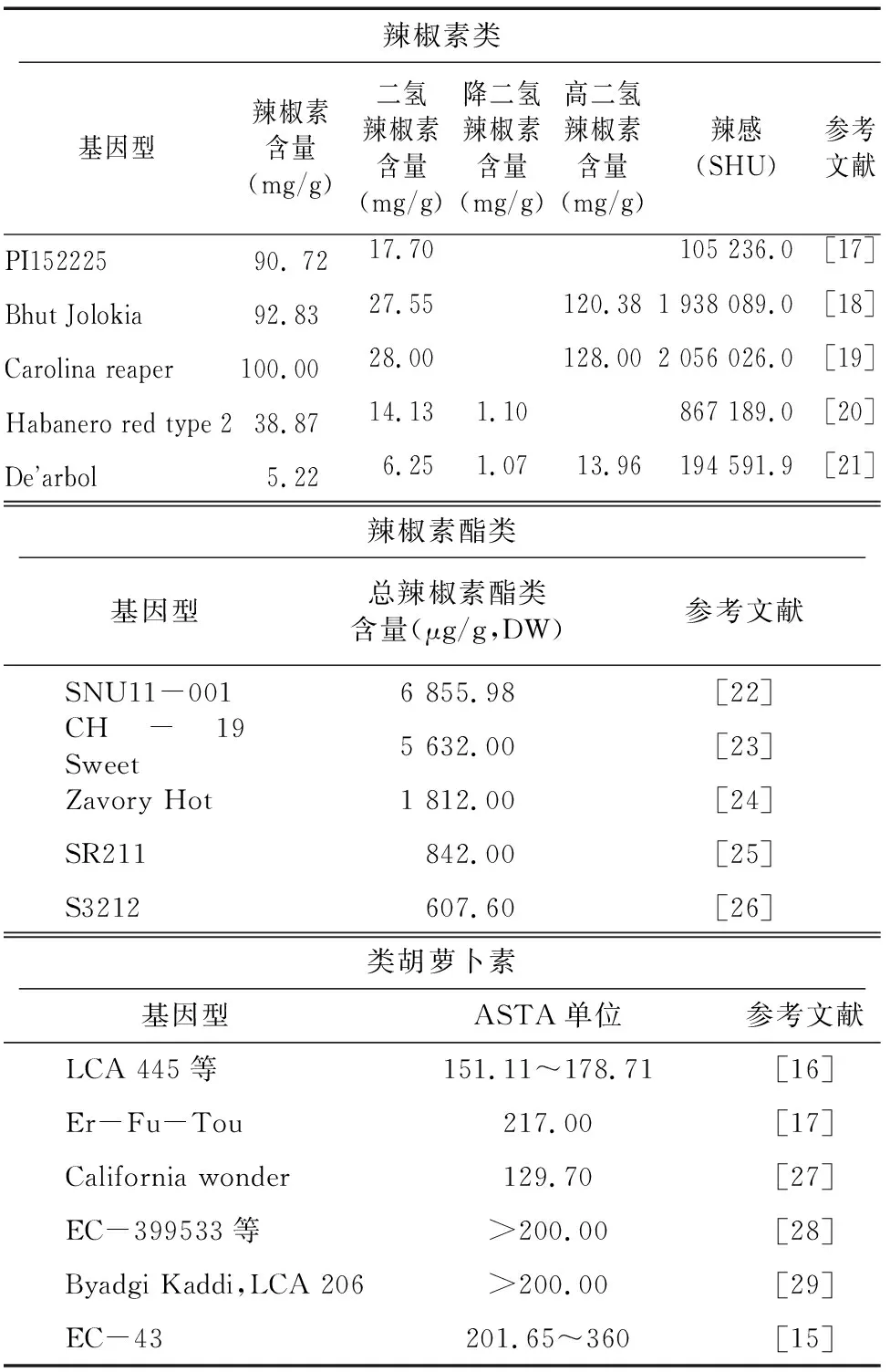
表1 富含辣椒素类、辣椒素酯类及类胡萝卜素的种质资源
我国的辣椒种植面积为78万hm,新鲜辣椒产量1 900万t,均位居世界第一。我国辣椒物种资源相对匮乏,然而在种内发生了部分独特的演化和分化现象。蓬桂华等通过测定贵州地方辣椒品质情况(辣椒素、粗脂肪、粗纤维和蛋白质含量),将贵州地区辣椒种质资源按照品质类型划分为4种:高、中、一般和特殊,为当地辣椒品质划分的依据和标准提供参考。张军等分别对来源于四川的辣椒种质83份和贵州的辣椒种质30份进行辣椒素含量的测定,分析辣椒种质资源的辣度情况,为培育不同辣度品质的辣椒品种提供种质资源。仅在2017—2019年期间,农业农村部非主要农作物品种登记就通过了多达2 801个辣椒新品种,其中包括高辣度、香辣、高维生素C含量等营养品质方面的品种。如高辣度辣椒品种金辣1号,辣椒素含量1.6%,辣味强,可为专用提取辣椒素的品种,现已在浙江一带广泛推广应用。
3 营养品质形成的生理机制
辣椒素类物质是在开花后大约20 d开始在果实中积累,其生物合成发生在胎座表皮细胞中,然后分泌到外细胞壁并在胎座表面的“小泡”中积累。种子不含辣椒素,但由于它们靠近胎座,所以会表现出一定的刺激性。辣椒素类物质是由香草胺分子缩合后产生的,香草胺分子是苯丙氨酸的衍生物,后者是一种由缬氨酸或亮氨酸合成的含有9~11个碳原子的支链脂肪酸(图1)。由于在氨基转移酶基因()中插入1个T碱基导致产生1个终止密码子,使其功能丧失,从而合成了辣椒素酯类物质。正是由于氨基转移酶不能催化香草醛合成香草胺,所以辣椒素酯类物质分子产生积累。辣椒素酯类物质的积累可能受到控制辣椒素积累的相同遗传机制的影响。辣椒红素在红辣椒果实成熟果皮的色素体的类囊体膜中积累,是类胡萝卜素生物合成途径的延伸产物,始于在辣椒果实发育和成熟期间通过八氢番茄红素合酶合成八氢番茄红素。几个去饱和反应将八氢番茄红素转化为橙色的-胡萝卜素,后者被氧化形成叶黄素。在辣椒中,辣椒红素/辣椒玉红素合酶(Ccs)合成辣椒红素和辣椒玉红素2种红色素。
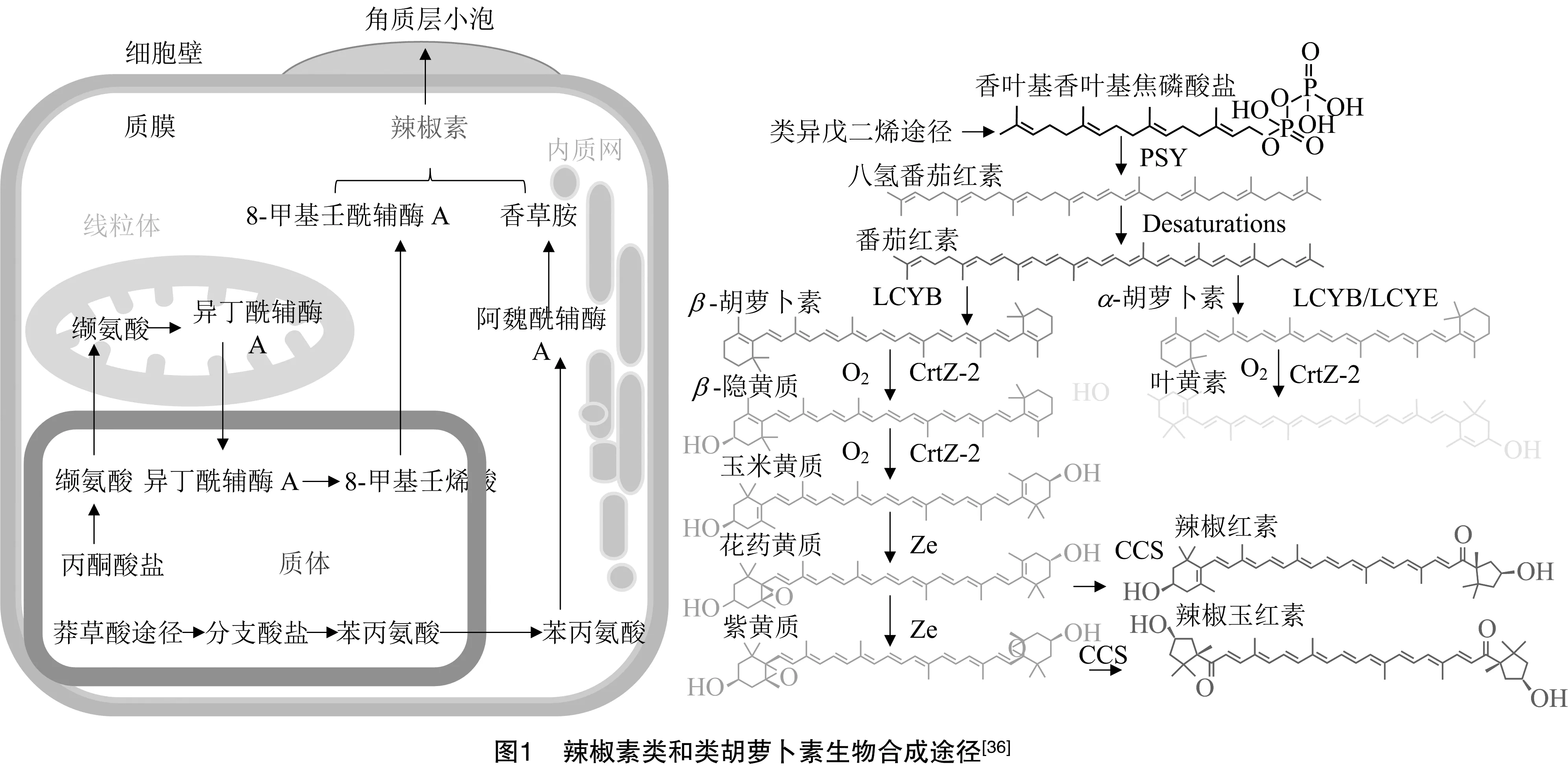
4 营养品质性状的分子生物学研究
关于辣椒遗传图谱的开发有多个报道,包括RAPD、RFLP、AFLP和SSR标记。不同类型的标记用于构建连锁图谱,以便可以使用传统的双亲作图群体识别控制特定数量性状的染色体区域。单核苷酸多态性(SNP)标记的引入为密集连锁图谱开辟了更多新机会,可用于绘制QTL和识别重要数量性状的候选基因。转录组序列、测序基因分型和全基因组测序方法加速了大量基于DNA的SNP标记的可用性。Li等开发了145对基于NGS重测序的高分辨率熔解(HRM)标记的引物对。在2个辣椒自交系中发现了indels(1~5 bp),并验证并绘制了252个InDel标记。这些研究共同表明,标记辅助选择策略可以应用于辣椒育种计划,以便于利用遗传变异。刘周斌在通过对转录组和蛋白质组的整合分析中发现在辣椒中,辣椒素的合成主要受牻牛儿基牻牛儿基焦磷酸合成酶 GGPS1(Capana04g000412)和辣椒红素-辣椒玉红素合成酶 CCS1(Capana06g000615)的调控。全基因组关联作图是使用双亲作图群体的另一种新方法,用于识别复杂性状的QTL。Nimmakayala等首次在辣椒使用GWAS,发现编码锚蛋白样蛋白、IK13家族蛋白、ABC转运蛋白G家族和五肽重复蛋白的基因中的SNP是辣椒素的主要标志物。2014年,实现了第一个栽培和野生辣椒的全基因组测序。辣椒的基因组比其他茄科作物的基因组要大得多:它的基因组大小为3 480 Mb,比马铃薯(840 Mb)、番茄(950 Mb)和茄子(1 127 Mb)大几倍。已确定76%~81%的辣椒基因组由转座因子组成。尽管基因组异染色质区域大量扩增的功能意义尚不清楚,但辣椒基因组是研究该现象的理想模型。虽然在拟南芥、番茄和马铃薯中发现了可能参与辣椒素类生物合成的基因同源物,但辣椒在13个基因家族中具有特定的重复,表明基因重复和新功能化。对编码已知控制辣椒素合成的酰基转移酶的基因分析表明,该基因的3个拷贝在高辣度种质中串联复制,而在该基因的启动子区域中存在缺失。Ou等比较了368个、4个和11个品种的测序(NGS)数据,并分析了基因变异。由此产生的辣椒泛基因组信息将成为辣椒重要的遗传资源。与茄科的其他成员(例如烟草和番茄)相比,从辣椒的培养组织进行植物的组织培养再生有些困难。小孢子胚胎发生是一种可用在育种中产生双单倍体的方法,并已得到优化。使用直接基因转移方法或农杆菌介导的基因转移方法在转基因辣椒植物中表达外源基因存在成功案例,使用CRISPR/Cas9系统在特定靶基因中引入突变的基因组编辑尚未在辣椒中实现。随着通过基因组工具在不同物种中鉴定出更多与园艺相关性状的候选基因,可以预期,未来以育种改良品种为目的的基因转移和基因组编辑的使用将会增加。
涉及植物组织培养和重组DNA技术的生物技术是强大的工具,可以弥补传统育种技术的不足,加快辣椒营养品质的改良。孙国胜等创建了一套适用于辣椒的遗传转化体系,该体系的优点是易操作、无选择性、周期短。以纳米磁性颗粒为载体,与质粒DNA等比例混匀,再与培养基中的辣椒花粉利用磁场的作用混合,经过人工授粉(花粉携带载体DNA),转化效率可达63.70%。利用该技术体系,结合最新的基因编辑技术,有望为辣椒营养品质的育种工作在效率和精准度方面提升一个新的台阶。
病毒诱导的基因沉默(virus-induced gene silencing,简称VIGS)技术,可应用于辣椒基因功能研究。通过VIGS体系抑制茉莉酸代谢信号途径中的基因的表达,使基因(辣椒素生物合成相关)表达量下降,也降低了辣椒素的含量。利用VIGS技术,沉默氨基酸转移酶基因(),使辣椒素类和辣椒素酯类物质含量减少。采用相同的技术研究辣椒中的、、和基因,都参与到辣椒素生物合成途径中,其表达量与辣椒素含量呈正相关。因此,利用VIGS体系调控辣椒营养品质相关成分,可获得优质的辣椒品种。
辣椒果实中营养成分的积累取决于基因型、果实发育阶段和环境因素,如光照、温度、水、CO、海拔、伤害和营养等。基因组学、转录组学、蛋白质组学和代谢组学是研究这些营养成分生物合成途径的有效方法。
5 栽培措施对辣椒品质影响的相关研究
辣椒品质与水、肥和介质等环境因素密切相关。中等灌溉量和中等施肥量耦合处理可在节水省肥的基础上使辣椒高产并且显著提高辣椒果实营养品质,高子星等研究发现灌水量和营养液的浓度显著影响辣椒的果实品质,游离氨基酸、可溶性还原糖以及维生素C含量等部分果实品质随营养液浓度的增加达到饱和。马国礼等的研究表明,水氮耦合对辣椒品质及产量影响显著,增加施氮量提高了辣椒各品质的含量,且施氮对辣椒品质的影响大于基质含水率,同时发现水肥最优处理的组合为中水高肥(W2F1)处理,即灌水量为田间持水量的60%。
相较单作栽培模式,间作模式呈现出明显的产量优势。辣椒、玉米、芋头间套作栽培模式研究表明此栽培模式不仅可以大幅降低辣椒病毒病的发病率,同时还能提高单位面积的综合产值,结果表明,3年间套种单位面积产值与单种相比,其涨幅分别达到59.71%、52.37%和56.15%。同时曾国璠在研究间作栽培措施对辣椒主要品质指标的影响中发现玉米种植密度28 500株/hm、辣椒种植密度39 000墩/hm、不剪枝剪叶、喷施0.2%硫酸锌溶液的玉米辣椒间作栽培措施组合可有效提高辣椒维生素C、辣椒素以及干物质含量,研究结果可为研发提高辣椒品质的栽培模式提供支撑。
6 展望
随着辣椒全基因组序列的公布,破解了辣椒丰富的遗传密码,这些基因组信息为研究者提供了大量潜在的靶标基因,有助于对辣椒营养品质进行精准高效改良。值得关注的是,这种巨大遗传多样性与观察到的表型变异性之间相关性研究还有待深入开展,尤其是基因组资源与辣椒中重要营养品质相关性状之间的关联是一个重要的研究领域。辣椒转基因技术体系的开发也很缓慢,在辣椒的遗传转化和组织培养再生过程中的效率不高。随着基因编辑技术的出现和不断升级,可以探索最新的基因组编辑技术及其在辣椒营养品质遗传改良中的潜在应用。新育种技术将成为辣椒营养品质育种必不可少的工具,有机融合传统的遗传育种选择和生物育种技术,将大大提高辣椒营养品质改良效率,满足人们对不同营养品质辣椒品种的需求,在增加食欲的同时,增进人类的健康水平。
猜你喜欢
老年博览·上半月(2024年2期)2024-03-15 00:05:24
天然产物研究与开发(2018年9期)2018-10-08 03:25:54
中国调味品(2018年9期)2018-09-15 04:54:34
天然产物研究与开发(2018年6期)2018-07-09 06:01:46
时代英语·高一(2017年5期)2017-11-14 15:52:20
中国果菜(2016年9期)2016-03-01 01:28:39
西南军医(2016年1期)2016-01-23 02:22:26
消费者报道(2015年7期)2015-12-05 02:32:16
食品科学(2013年22期)2013-03-11 18:29:34
食品科学(2013年19期)2013-03-11 18:27:54
