
江苏地区荷斯坦牛乳蛋白率Wood模型的非遗传影响因素分析
2022-08-25 09:25王海洋郭梦玲杨章平毛永江
家畜生态学报 2022年7期
王 洁,梁 艳,张 强,王海洋,郭梦玲,杨章平,毛永江*
(1. 扬州大学 动物科学与技术学院,江苏 扬州225009;2. 教育部农业与农产品安全国际合作联合实验室,江苏 扬州225009)
乳蛋白是由乳腺上皮细胞合成并分泌的营养物质,在牛奶与乳制品中均起着重要作用。泌乳期指的是奶牛分娩后从开始泌乳到泌乳结束这一阶段。泌乳曲线是用来描述泌乳期内奶牛群体或个体泌乳性状随时间变化而变化的一个数学模型,在奶牛生产和育种方面的有较强的指导作用,而乳蛋白率是奶牛泌乳性状的重要指标之一,也是衡量牛奶品质的重要指标。相关研究表明,乳蛋白率在泌乳初期逐渐下降,一定时间后达到最低点,后又随时间推移而逐渐回升,在不同泌乳阶段呈规律性变化,许多学者通过建立或借鉴各种数学模型来分析、描述、量化这一变化规律,发现Wood不完全伽玛函数模型(Wood模型)、逆多项式模型(Nelder模型)、Wilmink模型、Ali-Schaeffer模型和Dijkstra模型等对泌乳曲线有较好的拟合效果。孙先枝等研究发现,Wood模型适用于长三角地区荷斯坦牛乳蛋白率变化曲线的拟合。毛永江等研究发现,Wood模型对南方中国荷斯坦牛乳蛋白率变化曲线拟合度均在0.99以上。
相关研究表明,奶牛泌乳曲线模型参数受场效应、胎次、产犊季节等多种非遗传因素影响。近年来,随着牧场硬件条件的改善和饲养管理水平的提升,奶牛生产性能得到大幅度提高,从而对奶牛乳蛋白率泌乳曲线产生一定影响。因此,本研究拟用Wood模型对江苏地区12个牧场2017-2019年荷斯坦牛乳蛋白率的变化进行曲线拟合,定量的描述其变化趋势,并分析不同牧场规模、胎次、产犊季节、产犊间隔和305 d产奶量等非遗传因素对荷斯坦牛乳蛋白率Wood泌乳曲线参数的影响,从而为江苏地区荷斯坦牛科学饲养等方面提供参考依据。
1 材料与方法
1.1 数据来源
供试数据来自江苏地区12个奶牛场2017~2019年荷斯坦牛DHI测定日记录共580 025条,主要包括:牛号、测定日期、胎次、泌乳天数、乳蛋白率、305 d产奶量等。为保证最终结果的有效性,数据质控条件如下:胎次为1~5胎,泌乳天数≤305 d,乳蛋白率为1%~7%。最后合格记录数467 396条。根据江苏省气候特点,将产犊季节划分为春季(3-5月)、夏季(6-8月)、秋季(9-11月)、冬季(12月到次年2月);每30 d划分为1个泌乳月,最后一个泌乳月为271~305 d,共10个泌乳月。表1为数据的基本情况。
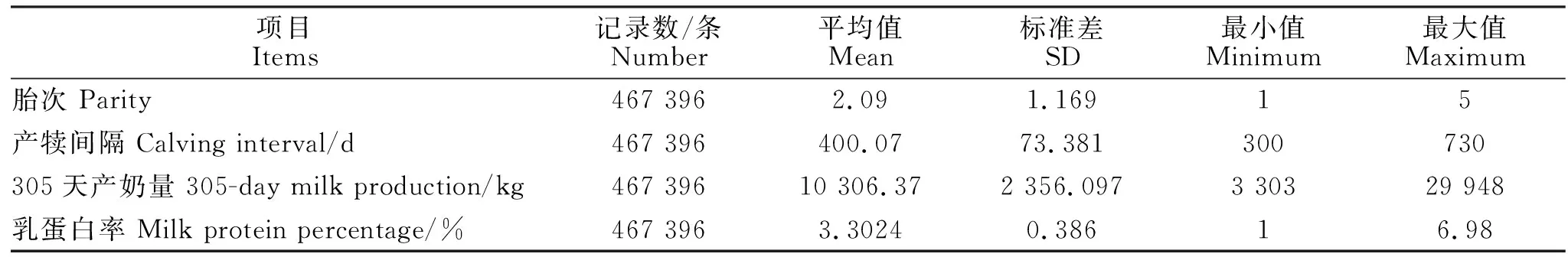
表1 泌乳数据的描述性统计Table 1 Descriptive statistils of lactating data
1.2 泌乳曲线拟合
用Wood不完全伽玛函数模型对测定日乳蛋白率变化进行拟合,基本模型如下:
Y=ate
式中:t为泌乳月;Y为时间t的乳蛋白率;a、b、c是模型参数。参数a是泌乳潜力;c是达到乳蛋白率最低点的速度;b是从乳蛋白率最低点上升的速度。a、b、c均由平均值±标准误形式表示。在进行曲线拟合时,各参数初始值的确定采用Olori等计算所得的结果。再使用如下公式换算出以下三个二级参数:
T= b/c
Y= a(b/c)e
Per= -(b+1)lnc
式中:T为达到乳蛋白率最低点所需的时间;Y为乳蛋白率的最低值;Per为乳蛋白率的持久力。先用Excel整合乳蛋白率数据,然后用SPSS(Ver 26.0)非线性回归子程序(Nonlinear Regression)进行拟合。拟合效果用拟合度R和误差均方进行评价。
采用最小二乘模型分析非遗传因素对奶牛乳蛋白率的影响,模型如下:
Y=μ+F+P+S+D+N+e
式中:Y为乳蛋白率的观察值;F为牛场规模的固定效应;P为胎次的固定效应;S为产犊季节的固定效应;D为产犊间隔的固定效应;N为305 d产奶量的固定效应;e为随机残差。各因素不同水平间的多重比较使用Duncan's法,显著性水平定义如下:≤0.01为极显著,≤0.05为显著,>0.05为不显著。根据上述影响测定日乳蛋白率的因素和水平,分别选取相应数据,利用如前所述方法分别进行Wood泌乳曲线拟合,并计算相应参数。
2 结果与分析
2.1 不同因素对荷斯坦牛乳蛋白率的影响
由表2可知,牧场、胎次、产犊季节、产犊间隔和305 d产奶量对荷斯坦牛乳蛋白率有极显著影响(<0.01)。其中,规模在5 000头以上的牧场荷斯坦牛乳蛋白率最高(3.31%),规模在2 001~5 000头的牧场荷斯坦牛乳蛋白率最低(3.23%);第2胎荷斯坦牛乳蛋白率为3.32%,极显著高于3~5胎(<0.01),第5胎乳蛋白率为3.25%,极显著低于其它胎次(<0.01);冬季产犊的荷斯坦牛乳蛋白率为3.31%,极显著高于其他产犊季节(<0.01),春季产犊乳蛋白率为3.28%,极显著低于其它产犊季节(<0.01);荷斯坦牛乳蛋白率随着产犊间隔的增加而极显著下降,分别为3.32%(300~365 d)、3.30%(366~400 d)、3.29%(401~420 d)、3.28%(421~440 d)和3.27%(>441 d);305 d产奶量为3 000~5 000 kg和5 001~7 000 kg的荷斯坦牛乳蛋白率极显著高于其他305 d产奶量范围奶牛的乳蛋白率(<0.01)。

表2 不同因素对乳蛋白率的影响Table 2 Effects of different factors on milk protein percentage
2.2 不同因素对日乳蛋白率曲线拟合的影响
由图1和表3可知,规模在5 000头以上的奶牛乳蛋白率曲线拟合度最大(0.9932)且误差均方最小(0.010),规模在2 001~5 000头的奶牛乳蛋白率曲线拟合度最小(0.9921);规模在5 000头以上的奶牛乳蛋白率最低值最大(3.11%),规模在1 000头以下的奶牛乳蛋白率最低值最小(3.02%);规模在1 000头以下的奶牛达到乳蛋白率最低值的时间最大(87.43 d),规模在5 000头以上的奶牛达到乳蛋白率最低值的时间最小(77.00 d);规模在1 000~2 000头的奶牛乳蛋白率持久力最大(3.81),规模在1 000头以下的奶牛乳蛋白率持久力最小(3.69);规模在1 000~2 000头的奶牛乳蛋白率潜力最大(3.106),规模在1 000头以下的奶牛乳蛋白率潜力最小(3.038);规模在1 000头以下的奶牛达到乳蛋白率最低点的速度和最低点上升的速度最大,分别为0.035和0.102。
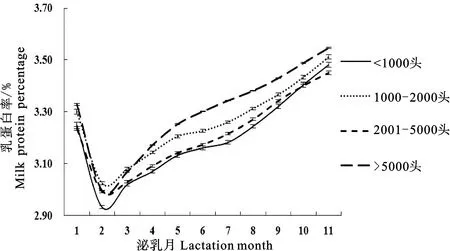
图1 不同牧场规模下Wood模型拟合泌乳曲线Fig.1 Lactation curves of Wood model for different farm sizes
结合图2和表3可知,荷斯坦牛第1胎的乳蛋白率曲线拟合度最大(0.9937)且误差均方最小(0.009),第5胎拟合度最小(0.9918);荷斯坦牛第2胎乳蛋白率最低值最大(3.10%),第5胎乳蛋白率最低值最小(3.06%);荷斯坦牛第4胎达到乳蛋白率最低值的时间最大(90.00 d),第1胎达到乳蛋白率最低值的时间最小(63.75 d);荷斯坦牛第1胎乳蛋白率持久力最大(3.92),第3胎乳蛋白率持久力最小(3.69);荷斯坦牛第3胎乳蛋白率潜力最大(3.108),第1胎乳蛋白率潜力最小(3.050);荷斯坦牛第3、4胎达到乳蛋白率最低点的速度最大(0.035),第4胎由最低点上升的速度最大(0.105)。
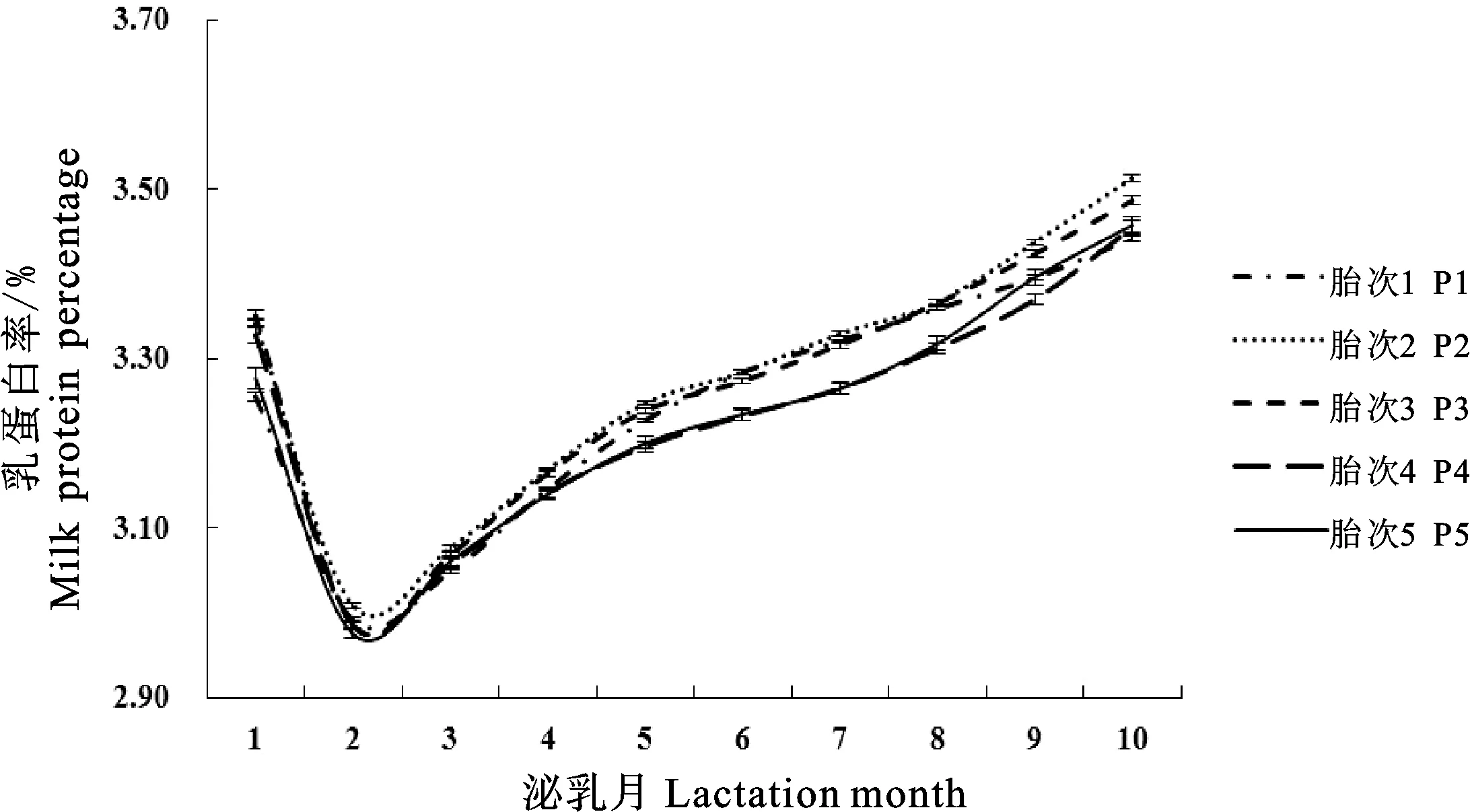
图2 不同胎次Wood模型拟合泌乳曲线Fig.2 Lactation curves of Wood model for different parities
结合图3和表3可知,荷斯坦牛冬季产犊乳蛋白率曲线拟合度最大(0.9933),夏季产犊乳蛋白率曲线拟合度最小(0.9925);冬季产犊乳蛋白率最低值最大(3.14%),春季产犊乳蛋白率最低值最小(2.99%);冬季产犊达到乳蛋白率最低值的时间最晚(99.33 d),夏季产犊达到乳蛋白率最低值的时间最早(28.57 d);秋季产犊乳蛋白率持久力最大(4.08),春季产犊乳蛋白率持久力最小(3.43);春季产犊达到乳蛋白率最低点的速度最大(0.047),冬季产犊乳蛋白率潜力和乳蛋白率由最低点上升的速度最大,分别为3.159和0.149。
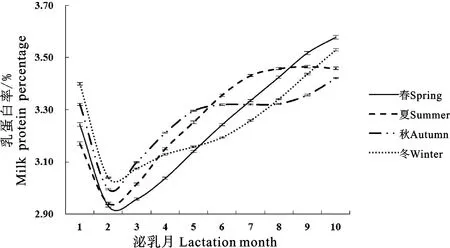
图3 不同产犊季节Wood模型拟合泌乳曲线Fig.3 Lactation curves of Wood model for different calving seasons
由图4和表3可知,荷斯坦牛产犊间隔为401~420 d时乳蛋白率曲线拟合度最大(0.9928)且误差均方最小(0.010),产犊间隔441 d以上时乳蛋白率曲线拟合度最小(0.9919);产犊间隔为300~365 d时乳蛋白率最低值最大(3.13%),产犊间隔为441 d以上时乳蛋白率最低值最小(3.02%);产犊间
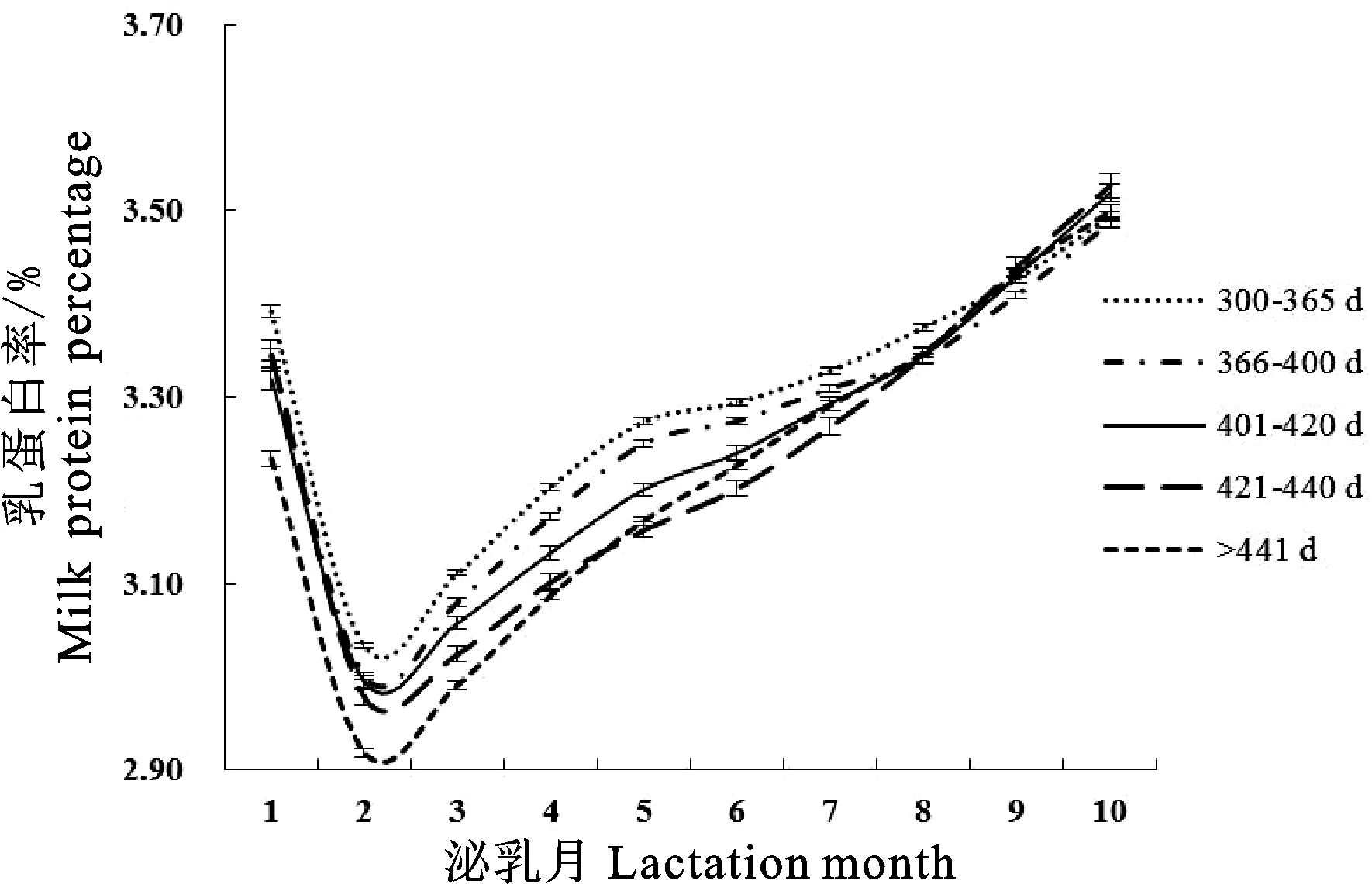
图4 不同产犊间隔Wood模型拟合泌乳曲线Fig.4 Lactation curves of Wood model for different calving interval
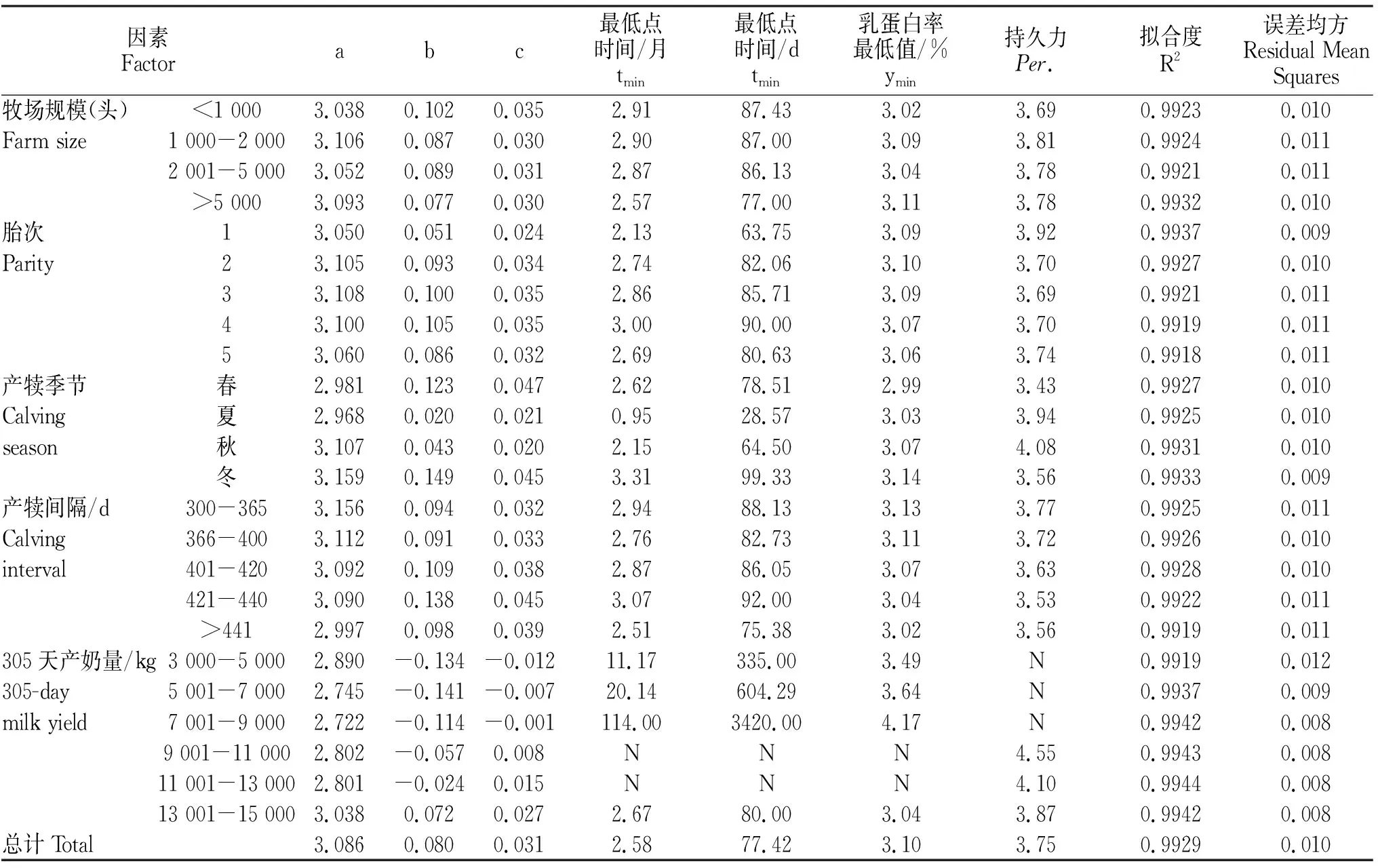
表3 不同因素日乳蛋白率Wood模型拟合参数Table 3 Wood model fitting parameters of daily milk protein ratio by different factors
隔为421~440 d时达到乳蛋白率最低值的时间最晚(92.00 d),产犊间隔441 d以上时达到乳蛋白率最低值的时间最早(75.38 d);产犊间隔为300~365 d时乳蛋白率持久力最大(3.77),产犊间隔为421~440 d时乳蛋白率持久力最小(3.53);产犊间隔为300~365 d时乳蛋白率潜力最大(3.156),441 d以上时乳蛋白率潜力最小;产犊间隔为421~440 d时达到乳蛋白率最低点的速度和由最低点上升的速度最大,分别为0.045和0.138。
结合图5和表3可知,305 d产奶量为11 001~13 000 kg的荷斯坦牛乳蛋白率曲线拟合度最大(0.9944)且误差均方最小(0.008),3 000~5 000 kg的荷斯坦牛乳蛋白率曲线拟合度最小(0.9919);305 d产奶量为7 001~9 000 kg的荷斯坦牛乳蛋白率最低值最大(4.17%),13 001~15 000 kg的荷斯坦牛乳蛋白率最低值最小(3.04%);305 d产奶量为7 001~9 000 kg的荷斯坦牛达到乳蛋白率最低值的时间最大(3420.00 d),13 001~15 000 kg的荷斯坦牛达到乳蛋白率最低值的时间最小(80.00 d);305 d产奶量为9 001~11 000 kg的荷斯坦牛乳蛋白率持久力最大(4.55),13 001~15 000 kg的荷斯坦牛乳蛋白率持久力最小(3.87);305 d产奶量为13 001~15 000 kg的荷斯坦牛乳蛋白率潜力、达到乳蛋白率最低点的速度和由最低点上升的速度最大,分别为3.038、0.027和0.072。
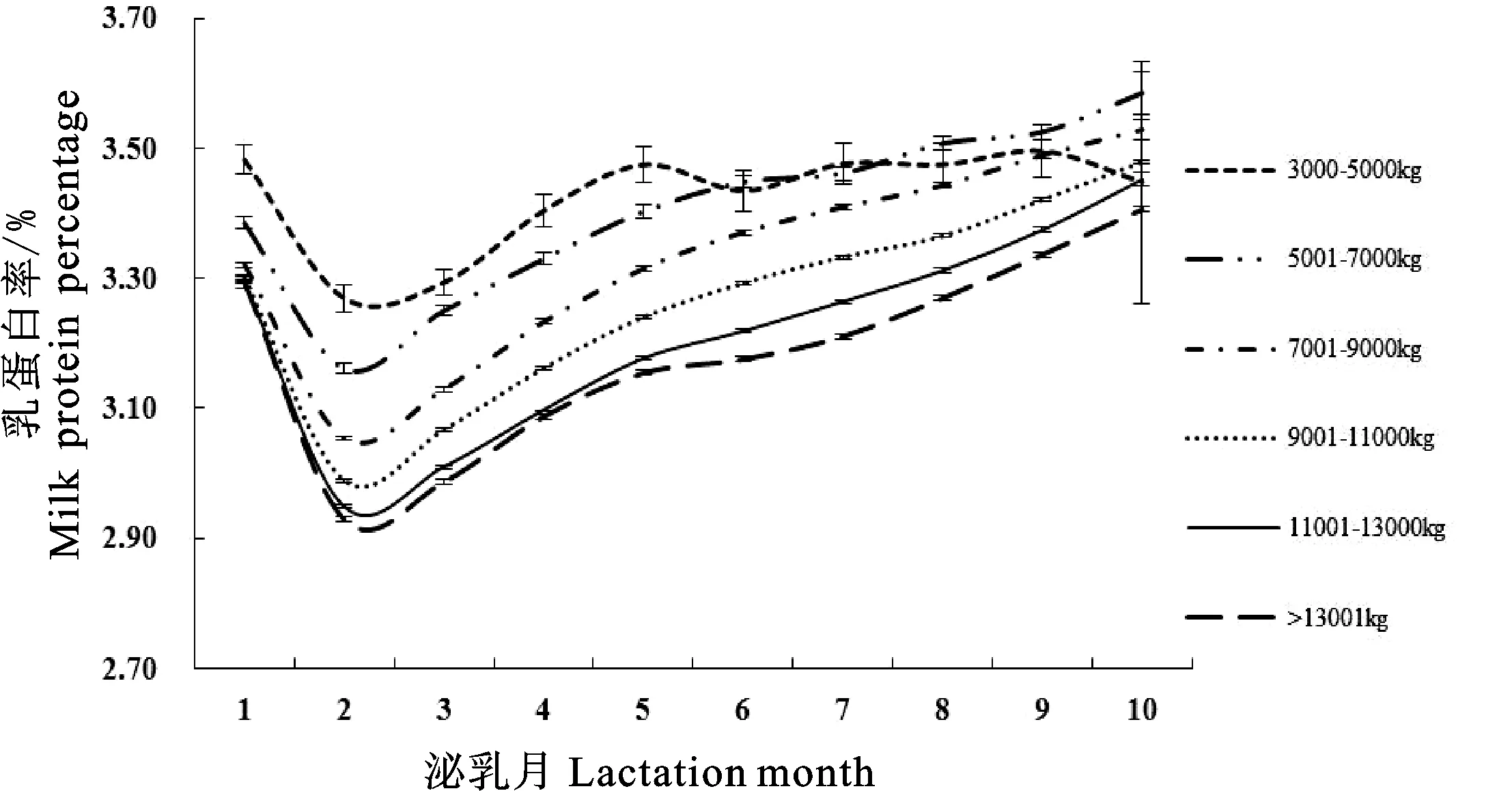
图5 不同305 d产奶量Wood模型拟合泌乳曲线Fig.5 Lactation curves of Wood model for different 305-day milk production
3 讨 论
3.1 牧场规模对Wood泌乳曲线模型的影响
王瑞军等和张佳兰等研究表明,场效应对荷斯坦牛泌乳曲线模型参数有显著影响,不同牧场泌乳曲线模型参数的差异显著。本研究发现,不同规模的牧场对荷斯坦牛乳蛋白率Wood泌乳曲线模型参数有一定影响,其中规模在5000头以上的牧场荷斯坦牛乳蛋白率曲线拟合度和乳蛋白率最低值最大,达到乳蛋白率最低值的时间最快。相关研究表明,规模在5 000头以上的牧场属于大规模牧场,场中设备机械化程度高,操作技术力量强,并且对不同泌乳期和产奶量水平的奶牛使用不同饲料配方的全混合日粮(Total mixed rations, TMR)进行分群饲养,进而提高了奶牛泌乳性能和泌乳效率,因此平均乳蛋白率和乳蛋白潜力较高。此外,大规模牛场较之中小型牛场具有更完善的饲养管理措施,并经过规范且科学的选育提高,因此泌乳效率高于小型牧场。
3.2 胎次对Wood泌乳曲线模型的影响
本研究发现,第5胎奶牛乳蛋白率曲线拟合度最低,乳蛋白率最低值最小;第1胎牛乳蛋白率持久力最大,潜力最小;第3胎牛乳蛋白率潜力最大,持久力最小。Roberto等通过其研究发现,Wood模型拟合群体泌乳曲线参数极显著受到胎次影响,头胎牛泌乳潜力较低。孙先枝等研究发现,1胎奶牛乳蛋白率持久力最高,3胎牛乳蛋白率潜力最大,持久力最小。上述研究与本研究基本一致。对于头胎牛,奶牛机体发育尚不完善,乳蛋白率潜力较低,泌乳后期乳蛋白率上升较慢,乳蛋白率持久力强。奶牛泌乳性能、乳蛋白潜力随着胎次的增加逐渐提升,第3胎时乳蛋白率潜力达到最大。就乳蛋白率而言,持久力越大,说明其维持较低值的能力越强,乳蛋白率越低,第3胎乳蛋白率持久力最小,泌乳性能最佳,乳蛋白率潜力最大。对于荷斯坦牛第5胎乳蛋白率曲线拟合度最低和乳蛋白率最低值最小,Knaus等和Oltenacu等研究表明奥地利和美国奶牛的平均淘汰胎次为3.3胎,鄢新义等研究发现中国荷斯坦牛平均淘汰胎次为2.86胎;高胎次奶牛机体免疫力降低,泌乳机能下降,但也不排除部分5胎以上奶牛生产性能较好,因此造成5胎奶牛乳蛋白率变化差异较大,从而出现泌乳曲线拟合度最低和乳蛋白率最低值最小。但这一结果还有待扩大样本量,进一步深入研究。
3.3 产犊季节对Wood泌乳曲线模型的影响
亓建刚等研究发现,产犊季节对Wood泌乳曲线模型参数均有一定影响,其中冬季产犊拟合度最高。本研究中,不同产犊季节对荷斯坦牛乳蛋白率和Wood泌乳曲线模型参数均有相应程度的影响,其中冬季产犊乳蛋白率变化曲线拟合度、乳蛋白率最低值和乳蛋白率潜力最大;夏季产犊拟合度最小,达到乳蛋白率最低值的时间也最快。相关研究表明,奶牛在夏季产犊机体免疫力下降,同时最易受外界环境影响,在热应激情况下奶牛的各项生理机能出现紊乱,为补足能量和糖的需求,糖原分解和糖异生增加,这使得蛋白质代谢增强,曲线拟合度下降。同时,奶牛夏季产犊受热应激影响,降低了奶牛干物质采食量和脂肪组织动员能力,并通过激素信号通路促进了葡萄糖的利用,从而使维持需求增加,用于泌乳的能量减少,乳蛋白率快速降低,达到乳蛋白率最低值的时间也最小。因此,从维持奶牛身体健康和提高泌乳性能方面而言,牧场应做好有针对性的防暑降温工作,同时在可能的范围内合理安排牛群配种时间,尽量避免奶牛在酷暑季节产犊,以获得更高的养殖效益。
3.4 产犊间隔对Wood泌乳曲线模型的影响
本研究发现,奶牛产犊间隔对乳蛋白率和Wood泌乳曲线模型参数有一定影响,其中产犊间隔为401~420 d时泌乳曲线拟合度最大,产犊间隔为441 d以上时乳蛋白率变化曲线拟合度、乳蛋白率潜力、乳蛋白率最低值和达到乳蛋白率最低值的时间最小。研究表明,奶牛下丘脑-垂体-卵巢轴受到生长激素(GH)、胰岛素样生长因子-I(IGF-I)、胰岛素、甲状腺激素和瘦蛋白等多种代谢物和激素调节。对于产犊间隔为441 d以上的荷斯坦牛,体内分泌信号因子IGF-I和瘦蛋白处于负平衡状态,机体的新陈代谢能力和生殖激素的分泌能力下降,繁殖疾病增多,从而使产妊间隔延长,泌乳天数也随之延长,从而导致产犊间隔较长,因此产犊间隔441 d以上的奶牛乳蛋白率变化曲线拟合度最低,乳蛋白率潜力和乳蛋白率最低值最小。
3.5 305 d产奶量对Wood泌乳曲线模型的影响
白音塔拉研究表明,不同305 d产奶量对乳蛋白率Wood泌乳曲线模型有一定影响,其中经产牛平均305 d产奶量为11 328.8 kg时Wood泌乳曲线模型拟合效果最好。本研究发现,305 d产奶量为11 001~13 000 kg的荷斯坦牛乳蛋白率变化曲线拟合度最大,3 000~5 000 kg的荷斯坦牛乳蛋白率变化曲线拟合度最小;305 d产奶量为13 001~15 000 kg的荷斯坦牛泌乳潜力、达到乳蛋白率最低点的速度和由最低点上升的速度最大,且乳蛋白率持久力最小。此外,305 d产奶量为3 000~5 000 kg的荷斯坦牛部分因为早产,部分可能为遗传和疾病等原因,导致泌乳性能下降,乳蛋白率曲线拟合度降低。对于305 d产奶量为13 001~15 000 kg时Wood泌乳曲线模型参数变化,其原因可能是由于本研究中荷斯坦牛305 d产奶量多集中在7001~13 000 kg之间,而产奶量在13 001~15 000 kg的样本量过少所致,因此还有待扩大样本量,以进行更深入的研究。
4 结 论
Wood模型适用于江苏地区荷斯坦牛测定日乳蛋白率变化曲线的拟合分析,且牧场规模、胎次、产犊季节、产犊间隔、305 d产奶量等非遗传因素对乳蛋白率Wood泌乳曲线模型参数均有显著影响。该结果为江苏地区荷斯坦牛精准养殖、选种选配等方面提供了参考依据。
猜你喜欢
作文·初中版(2020年10期)2020-11-25
食品界(2019年2期)2019-03-10
价值工程(2017年31期)2018-01-17
汽车科技(2016年6期)2016-12-19
食品安全导刊(2016年9期)2016-05-14
科教导刊·电子版(2016年3期)2016-03-14
科技与创新(2015年21期)2015-12-01
湖北畜牧兽医(2015年3期)2015-06-05
湖北畜牧兽医(2014年9期)2014-12-02
湖南饲料(2014年4期)2014-08-26
