
Functional diversity dominates positive species mixture effects on ecosystem multifunctionality in subtropical plantations
2022-08-11 04:11XiLiHuiWngJunweiLunSottChngBingGoYiWngShirongLiu
Forest Ecosystems 2022年3期
Xi Li, Hui Wng, Junwei Lun, Sott X. Chng, Bing Go, Yi Wng, Shirong Liu,*
a Institute of Resources and Environment, International Centre for Bamboo and Rattan, Key Laboratory of National Forestry and Grassland Administration/Beijing for Bamboo & Rattan Science and Technology, Beijing, 100102, China
b Key Laboratory of Forest Ecology and Environment of National Forestry and Grassland Administration, Ecology and Nature Conservation Institute, Chinese Academy of Forestry, Beijing, 100091, China
c Department of Renewable Resources, University of Alberta, 442 Earth Sciences Building, Edmonton, Alberta, T6G 2E3, Canada
Keywords:Ecosystem multifunctionality Species mixture effects Functional diversity Functional identity Mixed tree plantations Monospecific plantations
ABSTRACT Mixed-species plantations generally exhibit higher ecosystem multifunctionality than monospecific plantations.However, it is unclear how tree species functional composition influences species mixture effects on ecosystem multifunctionality. We selected 171 monospecific and mixed-species plantations from nine regions across subtropical China, and quantified 13 key ecosystem functional properties to investigate how species mixture effects on ecosystem multifunctionality are modulated by functional diversity and identity. We found that ecosystem multifunctionality was significantly higher (p <0.05) in mixed tree plantations than in monospecific plantations except the mixed-conifer species plantations. Across all regions, ecosystem multifunctionality was significantly higher (p <0.05) in mixed conifer-broadleaf plantations than in monospecific plantations of the corresponding species, but not different between mixed and monospecific coniferous plantations. The magnitude of species mixture effects on ecosystem multifunctionality varied greatly with tree species compositions. Taking Cunninghamia lanceolata Lamb. as an example, the effects varied from a range of 2.0%–9.6% when mixed with a conifer species to 36%–87% when mixed with a broadleaf species. The functional diversity was the dominate driver shaping ecosystem multifunctionality, while functional identity, as expressed by community-weighted mean of specific leaf area, also had a positive effect on ecosystem multifunctionality through the increased below-ground nitrogen and phosphorus stocks regulated by specific leaf area of the mixing tree species.Our study highlights the important role of functional diversity in shaping ecosystem multifunctionality across region-wide environmental conditions. Mixed conifer-broadleaf tree plantations with distinct functional traits benefit the enhancement of ecosystem multifunctionality,and the magnitude of species mixture effects is modulated by the functional identity of tree species composition; those relationships deserve a special consideration in multifunctional management context of subtropical plantations.
1. Introduction
Over the past decade, plantation management has shifted from pursuing the sole goal of wood production to multi-objective management aiming to improve multiple ecosystem functions,i.e.,multifunctionality(Liu et al., 2014). Establishing plantations with mixed tree species is viewed as one of the most effective silvicultural approaches to enhance the multifunctionality of plantations(Pretzsch et al.,2017;Schuler et al.,2017).A large body of literature suggests that mixed tree plantations can provide higher levels of multiple ecosystem functions compared to monospecific plantations, including productivity and soil nutrient storage and cycling (Gamfeldt et al., 2013; Lefcheck et al., 2015). These general patterns are mainly derived from positive relationships between tree species richness and ecosystem functioning. However, within each level of species richness,there is typically a high variation in functioning,mostly due to variations in the functional role played by different tree species (Baeten et al., 2019; Jonsson et al., 2019). The functional compositions of tree species,which are usually represented by a combination of species-specific functional traits(Diaz et al.,2016), may have similar or even greater effects on ecosystem functioning than species richness(Mouillot et al.,2011).Therefore,there is a need to understand how tree species functional composition influences the species mixture effects on ecosystem multifunctionality, in order to provide forest managers with practical tools for improving the multifunctionality of plantations.
Functional diversity and functional identity are two components representing functional composition(Tobner et al.,2016;Lin and Zeng,2018). In mixed tree plantations, different functional traits (functional diversity) may affect the resource acquisition strategy among tree species, leading to greater use of limited resources (Valencia et al., 2015;Turnbull et al., 2016; Barry et al., 2019) in view of the niche complementation hypothesis (Tilman et al., 1997; Gross et al., 2007). Complementary resource use favors higher ecosystem multifunctionality in mixed than in monospecific plantations. However, relatively low functional diversity of mixtures may support limited niche separation between species,with subsequent decreases in complementary resource use(Looney et al., 2021). In this case, the competition for resources among tree species may exceed the complementary effect, and consequently,leads to lower ecosystem functioning in mixed than in monospecific plantations (Forrester, 2014). The species mixture effects shift from negative to positive with increasing functional diversity.
Functional identity can be quantified by community-weighted mean trait values (Mokany et al., 2008), which are reformulated from the selection effect hypothesis(Grime,1998).The community-weighted mean of trait values is usually calculated using the traits measured and weighed by the relative abundance of each species(Violle et al.,2007).Functional traits are directly related to multiple ecosystem processes(Kunstler et al.,2016; Funk et al., 2017), and tree species with distinct functional traits can significantly amplify differences in their ecosystem functional effects(Kirwan et al., 2009; Kunstler et al., 2012). A positive (or negative)relationship of a community-weighted mean trait with multifunctionality suggests that mixed plantations composed of species with a high(or low)value of a trait promote multifunctionality. However, we know little about the relative contribution of functional diversity and functional identity to ecosystem multifunctionality, which constrains our understanding on how tree species mixture effects on ecosystem multifunctionality are modulated by tree species functional composition.
Tree species functional composition can also influence species mixture effects on ecosystem multifunctionality via changes in soil microbial diversity. Increasing evidence suggests that soil microbial diversity may play a key role in shaping ecosystem multifunctionality(Wagg et al., 2014; Delgado-Baquerizo et al., 2020). The functional structure of plant communities can directly (e.g., via litter quality and inputs)account for variations in soil microbial diversity(de Vries et al.,2012; Delgado-Baquerizo et al., 2018; Leff et al., 2018). Thus, to gain a mechanistic understanding of tree species mixture effects on ecosystem multifunctionality,we need to consider both direct and indirect effects of tree species functional composition on ecosystem functioning, by identifying changes in soil microorganisms and their relative contribution to ecosystem multifunctionality.
Here we used monospecific and mixed plantations composed of widely planted tree species in subtropical China to examine how tree species functional compositions influence the mixture effects on ecosystem multifunctionality and the underlying mechanisms. We test the following two hypotheses: 1) functional diversity promotes the positive species mixture effects on multifunctionality; and 2) functional identity has a similar role in regulating tree species mixture effects on multifunctionality.
2. Materials and methods
2.1. Study area and experimental design
In summer 2018, a total of 171 plots (each of 20 m × 20 m in size)from mature plantations composed of widely planted tree species were selected from nine regions across subtropical China with region-wide compositional and environmental gradients: Pingwu, Pengzhou,Yichang, Changsha, Taihe, Xinfeng, Longnan, Lechang, Pingxiang(Fig. S1). Mean annual temperatures ranged from 8°C to 20.5°C and mean annual precipitation ranged from 867 to 2,155 mm (Table 1). In each region, monospecific plantations and mixed plantations were selected for paired comparative analysis (see Table 1 for tree species composition in each region). We focused on the tree species of Cunninghamia lanceolata Lamb., Pinus massoniana Lamb., Schima superba Gardn., Castanopsis hystrix Miq., Pinus elliottii Engelm. and Cryptomeria japonica Linn. f., which are dominant plantation tree species in subtropical China. We selected mixed plantations and the corresponding monospecific plantations in similar environmental conditions (e.g., geology, soil type and topography) to minimize discrepancies among the abiotic conditions. Within each 20 m × 20 m plot, we randomly established three 5 m × 5 m subplots and five 1 m × 1 m quadrats for measurements of shrubs and herbs, respectively. Composite topsoil samples from 12 soil cores(0–10 cm depth)were collected and separated into two parts for biochemical and microbial analysis, and fine roots were separated from the composite soil for further analysis.
2.2. Ecosystem functional properties
We measured 13 properties of the above-and belowground ecosystem functions, including biomass of trees, understory and fine roots; aboveground wood production; concentrations of soil total carbon, total nitrogen,total phosphorus,nitrate nitrogen,and ammonium nitrogen;and activities of β-1,4-Glucosidase, cellobiosidase, β-1,4-N-Acetyl-glucosaminidase, and acid phosphatase. These variables are good proxies for biomass production and nutrient storage and cycling(Jax,2010;Hautier et al.,2018;Delgado-Baquerizo et al.,2017;Schuldt et al.,2018;van der Plas et al., 2018). We calculated biomass of trees using allometric equations (Luo et al., 2015). The aboveground wood production data were derived from tree ring measurements. On each 20 m × 20 m plot,we used 5.15 mm-diameter increment borers (Haglof CO300–5.15/4.3,Sweden)to extract wood cores from 5 individual trees randomly selected for each species, and the annual ring images were obtained after wood cores were mounted and sanded.The annual ring width was measured by LINTAB TM-6 measuring instrument (Germany). Annual radial increments were converted to annual biomass growth using allometric equations (Luo et al., 2015). Biomass of understory and fine roots were directly determined by harvesting and oven-drying.All shrubs within the three 5 m×5 m subplots and herbs within the five 1 m×1 m quadrats on each 20 m×20 m plot were harvested.The samples were dried at 70°C for 48 h and weighed for determination of the dry mass. Similarly, the fine roots collected within each 20 m×20 m plot were washed,dried to a constant weight, and then weighed.Total soil nitrogen and carbon concentrations were measured by a Vario ELIII elemental analyzer (Germany). The HClO4–H2SO4digestion method was used to measure total phosphorus. Soil nitrate nitrogen and ammonium nitrogen were measured by Continuous Flow Analysis(Smartchem 300,AMS,Italy).All extracellular enzyme activities were determined using the modified methods described in Saiya-Cork et al.(2002)and DeForest(2009).We also calculated the correlation coefficients between all measured variables for confirmation that there was neither a potential trade-off nor a high redundancy between the proxies of ecosystem function(Fig.S2).
2.3. Ecosystem multifunctionality
We employed averaging as a method to calculate the value of multifunctionality index(Zavaleta et al.,2010;Byrnes et al.,2014).To obtain an average value of ecosystem multifunctionality index,we first standardized each of the 13 ecosystem functional properties to a range from 0 to 1,using the formula (rawFunction - min(rawFunction)/(max(rawFunction) -min(rawFunction))),then the mean value was calculated for all standardized functions.This average value of ecosystem multifunctionality index is statistically robust,providinganintuitive wayandeasy-to-interpretdatafor assessing multifunctionality(Maestre et al.,2012).

Table 1 Basic information on the sample plots by region.
2.4. Functional identity and functional diversity
On each 20 m × 20 m plot, we obtained the increment cores and healthy leaves from 5 trees randomly selected for each species to measure the following functional traits: specific leaf area (SLA), leaf nitrogen content (LNC), leaf phosphorus content (LPC), wood density (WD), and leaf dry matter content (LDMC). These functional traits correlate with plant relative growth rate, nutrient acquisition and utilization (Wright et al., 2004), and competitiveness (Niinemets and Valladares, 2006;Anderegg et al., 2018). The functional diversity (FD) was calculated as the functional dispersion with the five functional traits,which measures the mean distance in a multidimensional trait space of individual species to the weighted centroid of all species (Lalibert'e and Legendre, 2010).Functional identity (FI) is usually estimated as community-weighted mean (CWM) of traits. Since some CWM of trait values were highly correlated with each other, a principal component analysis was performed for the CWM values of five traits.The first component explained 61.7%of the total variation,and was closely associated with CWM-LPC,CWM-LNC, CWM-LDMC, and in particularly CWM-SLA (Fig. S3). The second component explained 14.4%of the total variation and was closely related to CWM-WD. Therefore, CWM-SLA and CWM-WD were used to represent FI in our study.
2.5. Microbial diversity
Soil DNA extraction: Soil DNA was extracted from 0.25 g of soil samples using an MO BIO Powersoil®DNA extraction kit(MO BIO Laboratories,Carlsbad,CA,USA),accordingto themanufacturer's instruction.The bacterial 16S rRNA gene and fungal ITS1 region were amplified by a 515F (5′-GTGCCAGCMGCCGCGG-3′)/926R (5′-CCGTCAATTCMTTTGAGTTT-3′)and a ITS1-F(5′-CTTGGTCATTTAGAGGAAGTAA-3′)/ITS1-R(5′-GCTGCGTTCTTCATCGATGC-3′) primer, respectively. And then sequencing of the obtained PCR amplicons was performed on an Illumina MiSeq platform(Illumina,San Diego,CA,USA).
Bioinformatics analysis: The PE reads sequenced by Miseq were distinguished for each sample based on the barcode,followed by quality control and filtering of the sequences. We then spliced the sequences according to the overlapping relationships, and then performed quality control and filtering again to obtain optimized sequences. A Usearch software was used to perform OUT clustering analysis for optimized sequences based on a 97%similarity level.
We used the phylogenetic diversity of fungi(PDF)and bacteria(PDB)to calculate microbial biodiversity. Phylogenetic diversity is an important factor driving ecological functions (P'erez-Valera et al., 2015; Delgado-Baquerizo et al., 2017), and thus the phylogenetic diversity index can be used as a diversity index based on phylogenetic tree.To determine the phylogenetic diversity,the representative sequence of OTUs in each sample was used to calculate the distance of constructing phylogenetic tree, and then the branch lengths of all representative sequences were summed to generate the index value (Goberna and Verdú, 2016). We analyzed the relationship between community functional composition and microbial diversity(Fig.S4).
2.6. Statistical analysis
One-way analysis of variance was used to evaluate the differences in the 13 ecosystem functional properties and average ecosystem multifunctionality between monospecific and mixed plantations in each region, and heat-maps were then used for visualization of the differences.Linear mixed effects model was employed to test how species richness(SR), CWM-SLA, CWM-WD and FD affect the 13 ecosystem functional properties and ecosystem multifunctionality.In the analysis,a full set of models based on maximum likelihood estimation was developed as:
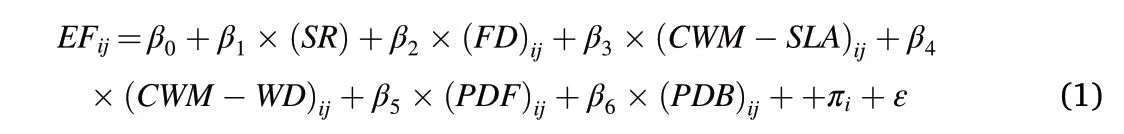
where the subscript“ij”designates the ith plot in the jth region;EFijis the predicted ecosystem functional properties or multifunctionality for the given plot; β0–β6are parameters to be estimated; πiis a random effect accounting for the study region;and ε is the residual error.Evaluating the optimal model was performed by Akaike information criterion (AICc).Prior to modelling, all data were normalized. Variance partitioning was used to quantify the proportion of total variation explained by each of the 13 functional properties and ecosystem multifunctionality.In addition to analyses using linear mixed effects models, we conducted a piecewise SEM to test direct and indirect relationships between plant species richness,functional composition of tree species,microbial diversity,and ecosystem multifunctionality.All statistical analyses were performed in R 3.6.2, with ‘FD’, ‘ggplot2’, ‘lme4’, ‘lmerTest’, ‘partR2’, ‘multifunc’,‘piecewiseSEM’and‘vegan’libraries/packages.
3. Results
3.1. Variations of ecosystem multifunctionality
Ecosystem multifunctionality was significantly higher (p <0.05) in mixed plantations than in monospecific plantations except the mixed coniferous tree plantations (Fig. 1a–b). Across all regions, ecosystem multifunctionality was significantly higher (p <0.05) in mixed coniferbroadleaf plantations than in corresponding monospecific coniferous or broadleaf plantations, but no significant difference was found between mixed and monospecific coniferous plantations (Fig. 1b). However, the positive mixture effects on multifunctionality were more profound in mixed conifer-broadleaf plantations than in monospecific coniferous plantations(Fig.1b).
The magnitude of tree species mixture effects on multifunctionality varied greatly with the tree species composition (Fig. 2). Taking an example of C.lanceolata,the magnitude of the mixture effects varied from a range of 2.0%–9.6%when mixed with a coniferous tree species to 36%–77% when mixed with a broadleaf tree species (Fig. 2). Across regions,the magnitude of tree mixture effects on ecosystem multifunctionality varied between 63%and 87%(Fig.2).
Almost all the specific functional property values were significantly higher in mixed conifer-broadleaf plantations than in monospecific coniferous plantations, especially the above-ground wood production,total soil nitrogen and phosphorus (Fig. 3). Most specific ecosystem functional property values in relation to biomass production were significantly higher in mixed conifer-broadleaf than in monospecific broadleaf plantations, especially the aboveground wood production(Fig. 3). However, there was no significant differences in most of the specific functional property values in relation to soil nutrient storage between mixed conifer-broadleaf and monospecific broadleaf plantations and between mixed coniferous and monospecific coniferous plantations,respectively(Fig.3).The magnitude of species mixture effects on aboveground wood production,soil total nitrogen and phosphorus varied with tree species compositions,e.g.,the respective values varied from a range of 8.7%–37%to 71%–184%,from-7.9%-37%to 54%–121%,and from-10%-54%to 95%–157%,when mixing C.lanceolata with a coniferous tree species and with a broadleaf tree species,respectively (Fig.4a–c).
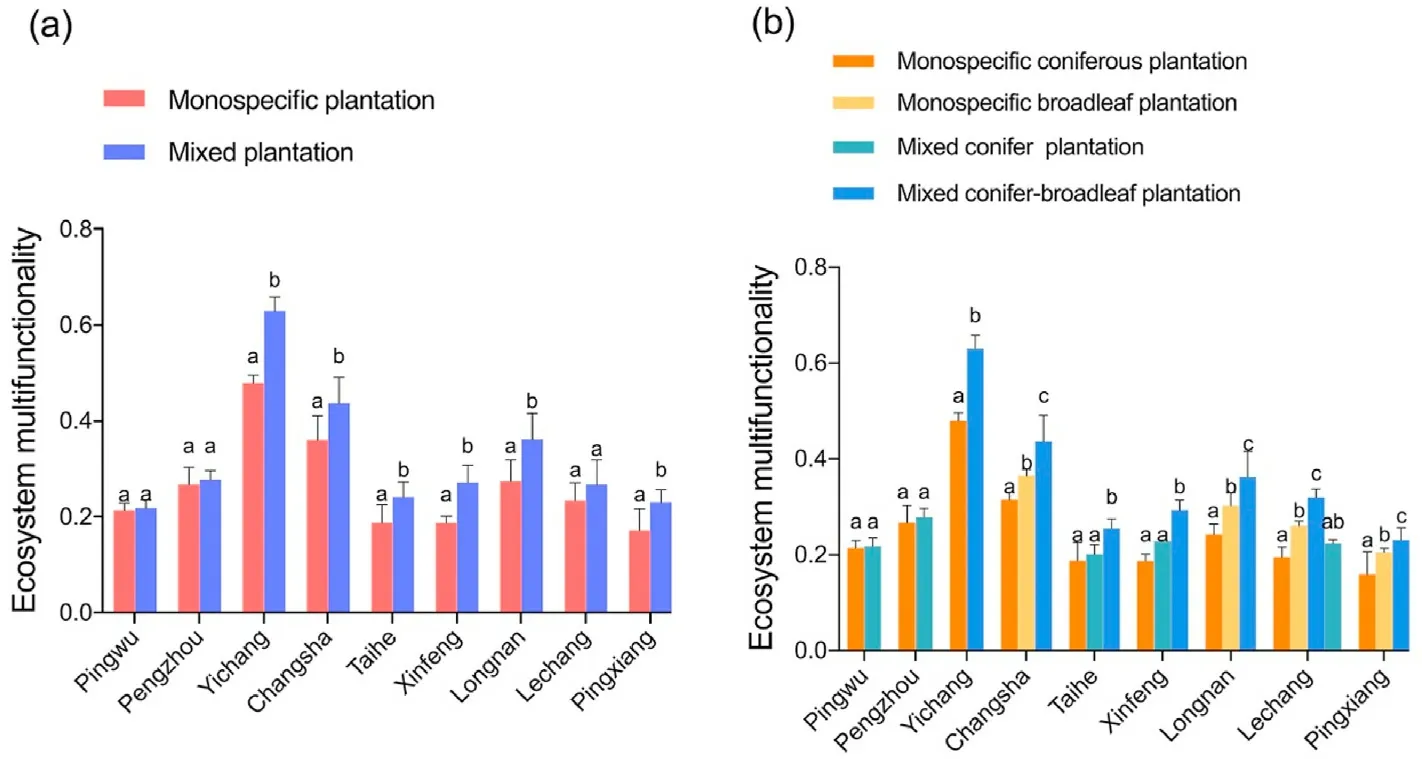
Fig. 1. Variation of ecosystem multifunctionality between mixed and monospecific plantations: a) comparison between mixed and monospecific plantations, and b)comparisons of monospecific coniferous plantation,monospecific broadleaf plantation,mixed conifer-broadleaf plantation and mixed conifer plantation.Error bars are for standard errors. Different letters indicate significant differences between stand types (α = 0.05).

Fig.2. Species mixture effects on ecosystem multifunctionality in relation to tree species composition.Cl:Cunninghamia lanceolata Lamb.;Ch:Castanopsis hystrix Miq.;Ss:Schima superba Gardn.;Pm:Pinus massoniana Lamb.;Pe:Pinus elliottii Engelm.;Cj:Cryptomeria japonica Linn.f.;Mg:Metasequoia glyptostroboides Hu&W.C.Cheng;Lf: Liquidambar formosana Hance; Cc: Castanopsis carlesii Hemsl.; Ns: Nyssa sinensis Oliv.; Cc2: Cinnamomum camphora Linn.; St: Sassafras tzumu Hemsl.
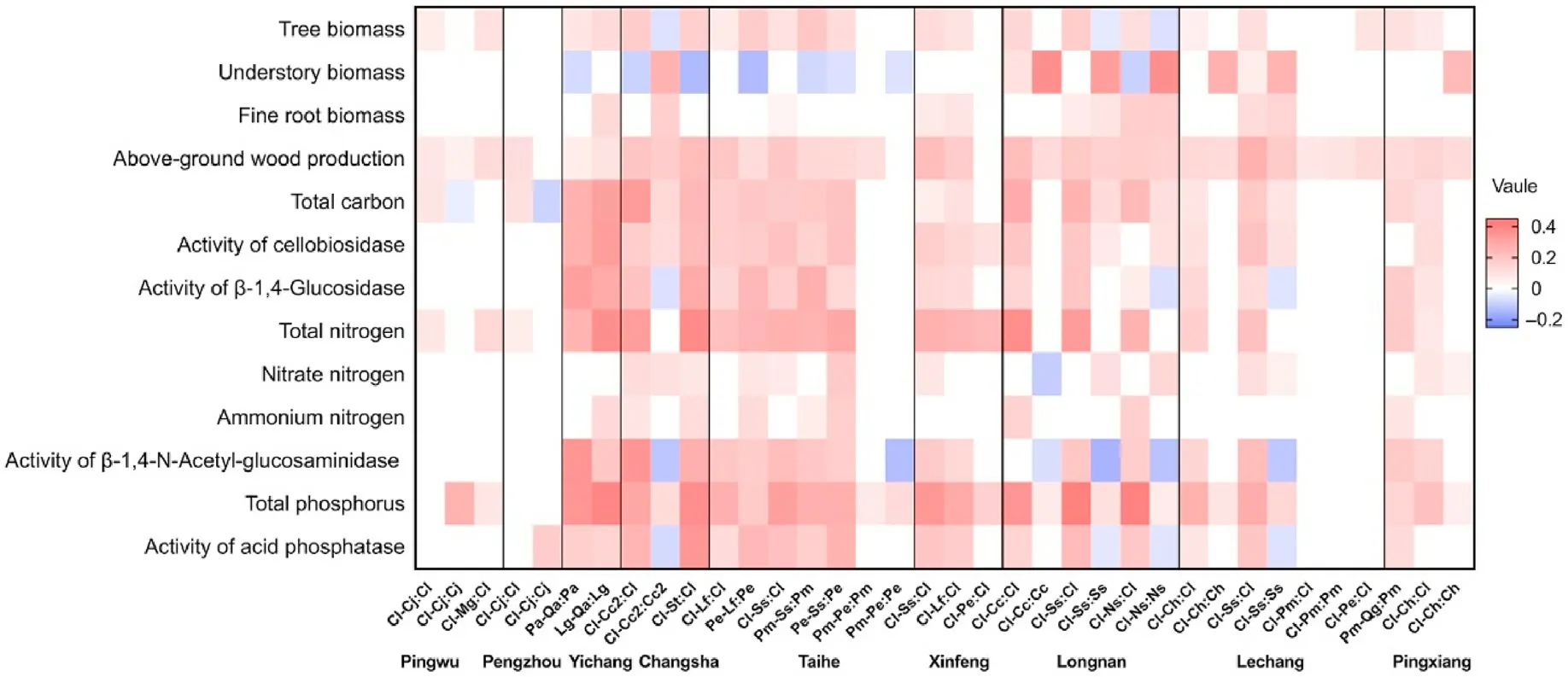
Fig.3. Differences in the 13 specific functional properties between mixed and monospecific plantations.The magnitude of differences is represented on a color scale:red and purple values represent significantly higher or lower specific ecosystem functional property values of mixed relative to monospecific plantations,respectively(p <0.05); darker the color, greater the difference. White values represent no significant difference. Horizontally, SppX-SppY:SppX represents comparison between mixed SppX-SppY plantation and monospecific SppX plantation,respectively.Species code:Cl,Cunninghamia lanceolata Lamb.;Ch,Castanopsis hystrix Miq.;Ss,Schima superba Gardn.; Pm, Pinus massoniana Lamb.; Pe, Pinus elliottii Engelm.; Cj, Cryptomeria japonica Linn.f.; Mg, Metasequoia glyptostroboides Hu & W. C. Cheng; Lf,Liquidambar formosana Hance;Cc,Castanopsis carlesii Hemsl.;Ns,Nyssa sinensis Oliv.;Cc2,Cinnamomum camphora Linn.;St,Sassafras tzumu Hemsl.;Pa,Pinus armandiiFranch.; Qa, Quercus alien Bl.; Lg, Larix gmelinii Rupr.; Qg, Quercus griffithii Hook. f. et Thoms. ex Miquel. (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
3.2. Key factors regulating ecosystem multifunctionality
The functional diversity, FD, was the dominant driver regulating ecosystem multifunctionality (Table 2, Fig. 5a), explaining a large proportion of total variations(Figs.5b and 6).Among the five indices of the functional identify, CWM-SLA significantly affected ecosystem multifunctionality (Table 2, Fig. 5a). Ecosystem multifunctionality significantly increased with the level of FD and CWM-SLA.FD and CWMSLA indirectly affected ecosystem multifunctionality by increasing PDF and PDB,respectively(Fig.5a,Fig.S4).FD and CWM-SLA were the main factors regulating the 13 specific functional properties (Table S1), of which FD played the dominant role in driving aboveground wood production relative to CWM-SLA(Table S1).Both FD and CWM-SLA had positive effects on soil nutrient storage and cycling(Table S1).

Fig. 4. Species mixture effects on specific ecosystem functional properties in relation to tree species composition. (a) Above-ground wood production; (b) Total nitrogen;(c)Total phosphorus.Species code:Cl,Cunninghamia lanceolata Lamb.;Ch,Castanopsis hystrix Miq.;Ss,Schima superba Gardn.;Pm,Pinus massoniana Lamb.;Pe,Pinus elliottii Engelm.;Cj,Cryptomeria japonica Linn.f.;Mg,Metasequoia glyptostroboides Hu&W.C.Cheng;Lf,Liquidambar formosana Hance;Cc,Castanopsis carlesii Hemsl.; Ns, Nyssa sinensis Oliv.; Cc2, Cinnamomum camphora Linn.; St, Sassafras tzumu Hemsl.
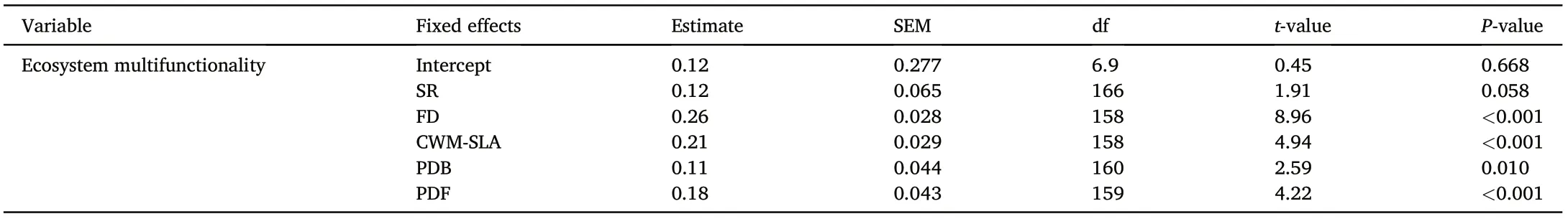
Table 2 The best-fitting mixed-effects model for ecosystem multifunctionality,including species richness(SR),functional diversity(FD),community weighted mean of specific leaf area(CWM-SLA), phylogenetic diversity of fungi (PDF), and phylogenetic diversity of bacteria (PDB). SEM:standard error of the mean;df: degree of freedom.
4. Discussion
This study evaluated the role of tree species functional composition in regulating species mixture effects on ecosystem multifunctionality by comparative analysis of functional diversity and functional identity between mixed and monospecific plantations.
4.1. Effects of functional diversity on ecosystem multifunctionality
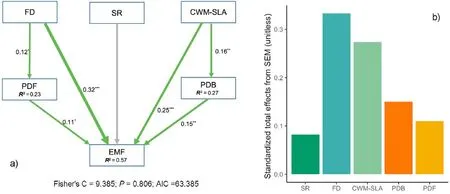
Fig. 5. Direct and indirect effects of plant species richness, functional composition, and microbial diversity on ecosystem multifunctionality. (a) Green solid arrows indicate significant (p <0.05) positive relationships. Grey arrows represent non-significant relationships. (b) The standardized total effects of each individual driver. SR: species richness; FD:functional diversity; CWM-SLA: community weighted mean of specific leaf area; PDF: species phylogenetic diversity of fungi;PDB:species phylogenetic diversity of bacteria; CWM-WD: community weighted mean of wood density, EMF: ecosystem multifunctionality.(For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)

Fig. 6. Relative importance of species richness (SR),functional diversity (FD), and functional identity (FI)including community weighted mean of specific leaf area(CWM-SLA),community weighted mean of wood density (CWM-WD), species phylogenetic diversity of fungi (PDF) and bacteria (PDB) on 13 specific functional properties and ecosystem multifunctionality,based on variance partitioning analysis on the bestfitting mixed-effects model for each ecosystem functional property and ecosystem multifunctionality.
In our study, ecosystem multifunctionality significantly increased with FD, and multifunctionality was significantly higher in coniferbroadleaf mixtures than in the corresponding coniferous and broadleaf monocultures, highlighting the importance of FD in dominating ecosystem multifunctionality (Valencia et al., 2015; Finney and Kaye,2017;Huang et al.,2019,2020).Our results suggest that the differences in functional traits among tree species in mixed plantations promote the complementary utilization of resources, which is a key mechanism underlying the positive mixture effect on ecosystem multifunctionality(Mouillot et al.,2011;Gross et al.,2017a;Mensah et al.,2020).Species mixture effects are often more pronounced in conifer-broadleaf mixtures,where FD in light use may promote stand-level efficiency (Ishii and Asano,2010;Lu et al.,2018).Coniferous and broadleaf tree species with divergent leaf morphology often have greater differences in SLA, LNC,LPC and LDMC, showing dissimilarity in the life history and light resource capture (Falster and Westoby, 2003; Wright et al., 2004). In general, trait-related niche differences among species could change the level of competitive interactions by facilitating niche partitioning and,consequently, reduce interspecific competition among coniferous and broadleaf species(Hooper et al.,2005;Díaz et al.,2007).As a result,the limited light resource can be used more efficiently in mixed conifer-broadleaf plantations (Sapijanskas et al., 2014; Jucker et al.,2015; Niklaus et al., 2017), leading to positive effects on ecosystem functions.
Although the heterogeneity of functional traits relating to light capture and utilization (e.g., SLA, LPC and LNC) can directly increase productivity rather than nutrient storage and cycling (Jucker et al., 2014;Vargas-Larreta et al., 2021), FD enhanced soil fertility by producing greater biomass and high-quality litter (Cong et al., 2014; Lange et al.,2015;Steinauer et al.,2015).In addition,our results showed that FD had a positive effect on PDF, possibly because of increased niche differentiation and resource partitioning. PDF promotes litter decomposition and the return of nutrients to soil (Nannipieri et al., 2010; Pei et al., 2016),which in turn facilitates tree growth, ultimately leading to greater ecosystem multifunctionality. This suggests that maintaining the ecosystem multifunctionality requires biodiversity at different trophic levels (Jing et al., 2015; Soliveres et al., 2016), and that the cascading effects between plant functional diversity and soil biodiversity contribute to the differences in ecosystem multifunctionality between monospecific and mixed plantations (Valencia et al., 2018). Moreover, mixed conifer-broadleaf plantations with higher ecosystem functions may be explained by complementary use of resources through the mechanisms that have not been explored, e.g., complementary foliar phenology(Forrester and Pretzsch,2015).
In our study, all coniferous trees are evergreen species, and most broadleaf trees are deciduous species, leading to temporal complementarity in light use (Forrester et al., 2017). Previous studies showed that mixing tree species would lead to overyielding in evergreen-deciduous species mixtures but not in deciduous-deciduous mixtures (Lu et al.,2016).Mixing tree species with distinct functional traits may also reduce the occurrence of pathogens and enhance ecosystem functions (Keesing et al., 2006; Haas et al., 2011). We found no evidence indicating a positive species mixture effect with the mixed conifer plantations.The result is consistent with the finding in the Interior West USA that there is a lack of species mixture effect in mixed conifer plantations (Looney et al.,2021), suggesting low FD restraining niche partitioning among tree species, and consequently, decreasing the chances for complementary resource use.
4.2. Effects of functional identity on ecosystem multifunctionality
An important result of this study showed that the magnitude of species mixture effects differs between coniferous and broadleaf monocultures due to differences in SLA, and this is consistent with our understanding that ecosystem functioning is also driven by FI (Grime,1998; Tobner et al., 2016). Our results revealed that CWM-SLA as the index of FI is also an important factor affecting ecosystem multifunctionality.CWM-SLA was as important as FD in explaining ecosystem functional properties relating to soil nutrient storage and cycling. Previous studies have shown that soil fertility is positively correlated with SLA(Ordo~nez et al.,2009).Broadleaf tree species tend to have a higher specific leaf area than coniferous tree species due to different leaf morphology, leading to a higher CWM-SLA in monospecific broadleaf plantations than in mixed conifer-broadleaf plantations. Monospecific broadleaf plantations with a higher CWM-SLA produce easily decomposable litter, increasing nitrogen and phosphorus availabilities in the soil(Kazakou et al.,2006;Cornwell et al.,2008;Zukswert and Prescott,2017). Additionally, CWM-SLA positively impacts soil nutrient storage by increasing PDB,as easily decomposable litter provide more substrates for microorganisms and improve microbial diversity (Liu et al., 2018).Therefore, compared with monospecific broadleaf plantations, mixed conifer-broadleaf plantations with a lower CWM-SLA did not show overperformance on soil nutrient storage. Conversely, the aboveground wood production was significantly higher in mixed conifer-broadleaf plantations than in monospecific broadleaf plantations, attributable to FD being more important in driving functional properties relating to biomass production compared to CWM-SLA. The results suggest that greater resources partitioning and facilitation tend to benefit aboveground production rather than underground ecosystem processes (Sapijanskas et al., 2014; Jucker et al., 2015). However, compared with monospecific conifer plantation, mixed conifer-broadleaf plantations have a higher CWM-SLA,and thus,both functional properties relating to soil nutrient storage and stand productivity were higher in mixed conifer-broadleaf plantations than in monospecific conifer plantation.As a result, the positive species mixture effects on ecosystem multifunctionality were more apparent in mixed conifer-broadleaf plantations than in monospecific conifer plantations. This implies ecosystem multifunctionality can be improved through optimizing tree species composition for enhancing SLA.
It was also observed that the species mixture effects varied among regions for the given two-species mixtures, indicating that species mixture effects are influenced by regional environmental factors,such as air temperature, water availability, and other soil properties (Liu et al.,2014), because relationships between complementarity and resource availability can vary greatly with environmental conditions (Forrester,2014).
5. Conclusions
In conclusion, ecosystem multifunctionality was higher in mixed conifer-broadleaf plantations than in corresponding monospecific plantations.Positive species mixture effects on ecosystem multifunctionality increased with FD and FI-expressed by CWM-SLA that are dependent on optimal tree species composition.Our study highlights the important role of tree species functional composition in shaping ecosystem multifunctionality of subtropical plantations. Mixtures of conifer-broadleaf tree species with distinct functional traits were beneficial to maintaining a high level of ecosystem multifunctionality, and the magnitude of species mixture effects depended largely on the tree species composition.
Ethics approval and consent to participate
Not applicable.
Funding
This study was funded by the National Natural Science Foundation of China(No.31930078)and the National Key Research and Development Program of China(No.2021YFD2200405).
Authors’contributions
Shirong Liu,Hui Wang and Junwei Luan conceived the study;Shirong Liu provided project support;Xi Li performed the research and analyzed the data; Xi Li, Shirong Liu and Junwei Luan wrote the manuscript;Shirong Liu, Scott X. Chang, Junwei Luan, Hui Wang and Yi Wang contributed to editing. Xi Li and Bing Gao conducted the fieldwork and collected the data.All authors gave final approval for publication.
Availability of data and materials
The datasets used or analyzed during the current study are available from the corresponding author on reasonable request.
Consent for publication
Not applicable.
Declaration of competing interest
The authors declare that they have no competing interests.
Acknowledgements
We are grateful to Experimental Center of Tropical Forestry,Chinese Academy of Forestry and Qianyanzhou Subtropical Forest Ecosystem Observation and Research Station,Chinese Academy of Sciences for their technical supports and helps. This study was funded by the National Natural Science Foundation of China (No. 31930078), the Ministry of Science and Technology of China (No. 2021YFD2200405), and Fundamental Research Funds for ICBR(Nos.1632019006,1632021023).
Abbreviations
FD functional diversity
FI functional identity
CWM-SLA community-weighted mean of specific leaf area
CWM-LNC community-weighted mean of leaf nitrogen content
CWM-LPC community-weighted mean of leaf phosphorus content
CWM-LDMC community-weighted mean of leaf dry matter conten
CWM-WD community-weighted mean of wood density
PDF phylogenetic diversity of fungi
PDB phylogenetic diversity of bacteria
SR species richness
EMF ecosystem multifunctionality.
Appendix A. Supplementary data
Supplementary data to this article can be found online at https://do i.org/10.1016/j.fecs.2022.100039.

- Forest Ecosystems的其它文章
- Vegetation structure and edaphic factors in veredas reflect different conservation status in these threatened areas
- Black locust coppice stands homogenize soil diazotrophic communities by reducing soil net nitrogen mineralization
- Responses of soil CH4 fluxes to nitrogen addition in two tropical montane rainforests in southern China
- Importance of Quercus spp. for diversity and biomass of vascular epiphytes in a managed pine-oak forest in Southern Mexico
- Novel evidence from Taxus fuana forests for niche-neutral process assembling community
- Using machine learning algorithms to estimate stand volume growth of Larix and Quercus forests based on national-scale Forest Inventory data in China