
砷胁迫下接种丛枝菌根真菌对棉花光合特性和叶肉细胞超微结构的影响
2022-08-08 03:21:20龚明贵刘凯洋魏亚楠白娜邱智军张巧明
棉花学报 2022年3期
龚明贵,刘凯洋,魏亚楠,白娜,邱智军,张巧明
(1.河南科技大学食品与生物工程学院,河南洛阳 471023;2. 河南科技大学食品与生物工程学院微生物资源开发与利用重点实验室,河南 洛阳 471023;3. 安阳市疾病预防控制中心,河南 安阳 455000;4.河南科技大学园艺与植物保护学院,河南 洛阳 471023)
砷(arsenic, As)是一种在自然界中广泛分布的非(类)金属元素。 As 矿的开采利用,含As 农药、化肥的使用,工业生产及汽车排放产生的大量含As 有害气体等均会直接或间接地造成土壤As 污染[1]。 全球至少有5 000 多万人口正面临着地方性As 中毒的威胁,我国也是受As 毒害较为严重的国家之一,湖南郴州As 矿区,内蒙古、贵州等As 含量较高的地区以及云南、广西、广东等省区都存在大面积As 污染和As 中毒等问题[2]。As 极易在土壤- 植物- 人体系统中进行生物转移和积累,对生态环境和人类健康造成巨大的危害。 As 可与植物体内的咪唑、羧基等结构间接结合或与DNA、蛋白质直接结合,从而干扰植物体内一系列正常的酶促反应,在光合磷酸化和氧化磷酸化的循环中,As 与ADP 形成的ADP-As5+复合物可导致ATP 产能下降, 影响正常的光合代谢反应[3-4]。 过量的As 还可造成植物体内积累大量的活性氧,降低蒸腾速率,破坏叶绿体膜结构,进而损伤植物光合系统、抑制光合效率,影响植物的生长发育,严重时可导致植株死亡[5-7]。 植物的呼吸作用、光合作用以及荧光参数是检验植物对As 胁迫耐受性的重要生理指标, 通过对这些指标的评价,有利于我们更加深入地了解植物在As 胁迫下的抗性机理。
丛枝菌根真菌(arbuscular mycorrhizae fungi,AMF)广泛存在于土壤中,目前已有根孢囊霉属(如根内根孢囊霉Rhizophagus intraradices、异形根孢囊霉Rhizophagus irregularis) 和斗管囊霉属(如摩西斗管囊霉Funneliformis mosseae、地斗管囊霉Funneliformis geosporum)等27 属约300 种AMF 被 鉴 定 出 来(http://www.amf-phylogeny.com/)。 AMF 可与大多数陆生植物建立菌根共生体系,能够明显提高植物抵御生物和非生物胁迫的能力[8],使宿主的碳素循环过程和光合作用得到加强[9]。 在被As 污染的土壤中,AMF 可通过其菌丝网络、丛枝、泡囊等结构吸附、固定土壤中的As,从而缓解As 对宿主植物造成的伤害[10]。 研究表明,接种AMF 可提高光合作用电子传递效率,增强植物对光能的吸收利用,从而提高光合效率[9,11]。利用AMF 提高植物的重(类)金属耐受性已成为植物修复土壤技术研究与应用的重点,但AMF 是否能够通过影响植物光合作用减缓重(类)金属造成的损伤仍需进一步研究。
棉花为纤维类作物, 具有栽培历史悠久、抗逆性较强、生物量较大等优点,其较大的生物量可从土壤中吸收更多的重(类)金属,不易造成再次污染,且其纤维产品不进入食物链,是土壤重(类)金属污染修复的优势作物[12]。 近年来广大学者就植物对As 的耐受机理做了大量研究, 但主要集中在蔬菜作物和粮食作物上[7],有关As 胁迫对棉花影响的研究鲜有报道。 本研究团队发现100 mg·kg-1As 浓度下棉花表现出明显损伤,As胁迫下AMF 可与棉花形成良好的共生关系,提高棉花对As 的耐受性[13]。 本研究分析了不同As浓度处理下,AMF 接种对棉花的气孔形态特征、气体交换参数、叶绿素荧光参数以及叶肉细胞超微结构等的影响, 以进一步明确AMF 对棉花光合特性的影响,探究AMF 接种提高棉花对As 的耐受性的可行性,为AMF 在土壤As 污染修复中的应用提供一定的参考。
1 材料与方法
1.1 试验材料
供试棉花种质为大铃棉69 号, 由中国农业科学院棉花研究所提供。 供试菌种为根内根孢囊霉(编号BGC BJ09)和摩西斗管囊霉(编号BGC XZ02A),由北京市农林科学院植物营养与资源研究所提供。 接种混合物包括沙子、孢子(每克土壤包含约23 个孢子)、 菌根碎片和根外菌丝体。供试土壤为黄土和砂土,均从河南科技大学植物园获得。
1.2 试验设计
选取饱满完整的棉花种子用75%(体积分数,下同)乙醇消毒15 min 后,用去离子水冲洗干净, 在30 ℃恒温条件下水浴泡发12 h 使其充分吸收水分,剥去种皮后,将种子置于恒温培养箱中,28 ℃条件下培养至萌发。
按照刘凯洋等[13]的方法配制土壤基质并进行灭菌处理。以喷洒Na3AsO4·12H2O 水溶液的方式加入外源As,使土壤As 浓度分别为100 mg·kg-1、200 mg·kg-1,将不喷洒砷盐溶液的土壤基质设为空白对照, 即0 mg·kg-1(原土壤基质中不含As元素)。 在塑料花盆(底径15 cm、口径18 cm、高度25 cm)中分别装入3.0 kg 上述土壤基质。试验设置接种根内根孢囊霉(以下简称RI)和接种摩西斗管囊霉(以下简称FM)处理,在花盆中距土壤表面1/3 深度层接种30 g 相应的菌根混合物,以接种经高温灭菌的同重量根内根孢囊霉菌种为对照(CK)。在距土壤表层2 cm 深度播种4~5粒萌发程度一致的棉花种子,出苗2 周后每盆保留2 株长势较好的棉花幼苗, 每组处理设10 盆重复。 本试验于2021 年7 月—10 月在河南科技大学温室内进行,棉花生长温度为20~35 ℃,相对湿度为50%~75%,光照/ 黑暗时间为12 h/12 h。 棉花生长过程中,每周浇灌Hoaglang's营养液[14]50 mL,为了减轻磷对AMF 菌丝生长和孢子萌发的抑制,将营养液中的磷含量调整为原配方的1/4。
1.3 测定指标及方法
棉花培养90 d 后取样,将各处理组随机分为3 份, 一部分用于棉花植株叶片气孔形态参数测定,一部分活体植株的叶片用于气体交换参数和叶绿素荧光参数的测定,一部分用于叶片细胞超微结构观察。
1.3.1气孔形态特征测定。 参照王凯丽等[15]的方法测定棉花倒3 叶下表皮气孔的长度、 宽度、开度和密度。
1.3.2叶片气体交换参数测定。 在室外晴朗无风条件下, 采用LI-6400 光合仪(LI-COR 公司,美国),使用开放式气路系统,设置内置红蓝光源光强为1 000 μmol·m-2·s-1,CO2浓度为400 μmol·mol-1,选取棉花顶部向下第一片完全展开的叶中部进行测定,获取净光合速率(net photosynthetic rate,Pn)、气孔导度(stomatal conductance,Gs)、胞间CO2浓度(intercellular CO2concentration,Ci)和蒸腾速率(transpiration rate,Tr)数据。
1.3.3叶绿素荧光参数测定。 测定前对棉花进行20 min 暗适应, 采用MINI-PAM 叶绿素荧光仪(Walz 公司,德国)测定棉花实际光合效率(actual photosynthetic efficiency,YⅡ)、最小荧光(minimum fluorescence,F0)、最大荧光(maximum fluorescence,Fm)、最大光化学量子产量(maximum photochemical quantum yield,Fv/Fm)、光化学淬灭系数 (photochemical quenching coefficient ,qP)、非光化学淬灭系数(non photochemical quenching coefficient,NPQ)。 在1.3.2 选取的同一叶片测定上述参数。
1.3.4叶片叶肉细胞超微结构观察。 取棉花倒数第4 片真叶,用蒸馏水冲洗干净。参照徐义昆等[16]的方法观察叶肉细胞内叶绿体、线粒体等超微结构并拍照。 具体操作流程如下:
(1)固定:将新鲜组织切成1 mm2小块置于2.5%戊二醛溶液中,固定4 h;将固定后的组织样本用0.1 mol·L-1磷酸缓冲液漂洗数次后置于1%锇酸中,4 ℃保存过夜。
(2)脱水:以乙醇-环氧丙烷为脱水剂对组织样本进行脱水,先用30%、50%、70%、80%、95%、100%乙醇梯度脱水(每次40 min),再用100%环氧丙烷脱水3 次(每次30 min)。
(3)包埋:分别用环氧丙烷-812 树脂2∶1 包埋液(体积比,下同)、环氧丙烷-812 树脂1∶2 包埋液进行渗透过渡,过渡时间分别为4 h、12 h;然后用812 树脂纯包埋液分别渗透12 h、4 h;将渗透完全的组织样本用纯812 树脂包埋液进行包埋,包埋完成后置于60 ℃干燥箱中干燥2~3 d 至树脂完全聚合。
(4)切片:对包埋块进行修整后置于超薄切片机(SM2010R,Leica,德国)上进行切片,厚度为70 nm。
(5)染色观察:先后用3%醋酸铀饱和酒精溶液和枸橼酸铅溶液分别染色5~8 min,染色后用超纯水漂洗,吸干其表面水分后在透射电子显微镜(Talos L120C TEM,Thermo Scientific,美国)下观察并拍照。
1.4 数据分析
以上所有试验均重复测量3 次。 数据以平均值±标准差形式表示。 利用SPSS 23.0 软件进行数据统计分析,采用新复极差法检验不同处理间各指标的差异显著性。 在Excel 2019 中制作图表。
2 结果与分析
2.1 As 胁迫下AMF 接种对棉花气孔形态特征的影响
随着As 胁迫的加重, 棉花叶片气孔形态特征发生明显变化,气孔的长度、宽度、开度和密度均呈下降趋势(表1)。不同As 浓度下,AMF 接种组的棉花各气孔形态指标数值相比于CK 均有不同程度的增加。 土壤As 浓度为0 mg·kg-1时,与CK 相比,RI 处理的气孔长度显著增加,FM 处理的气孔长度和开度均显著增加。100 mg·kg-1As胁迫下,与CK 相比,RI 处理的气孔长度显著增加,FM 处理的气孔长度、 开度和密度均显著增加。 200 mg·kg-1As 胁迫下,与CK 相比,RI 处理的气孔长度显著增加,FM 处理的气孔长宽、宽度和开度均显著增加。 不同As 浓度下,FM 处理的气孔长度、宽度、开度和密度均高于RI 处理。 以上结果表明,As 胁迫抑制了棉花叶片气孔的发育和开放程度,降低蒸腾作用,而AMF 接种可显著缓解这种抑制效应,且FM 较RI 效果更好。

表1 不同As 浓度下AMF 接种对棉花气孔形态特征的影响Table 1 Effects of AMF inoculation on morphological characteristics of cotton stomata under different As concentrations
2.2 As 胁迫下AMF 接种对棉花气体交换参数的影响
由表2 可知,棉花叶片净光合速率随外源As浓度增加显著降低,在200 mg·kg-1As 浓度下达到最低水平。AMF 接种(200 mg·kg-1RI 除外)能显著提高叶片净光合速率,与CK 相比,在0、100、200 mg·kg-1As 浓度条件下,RI 处理的净光合速率分别提高了54.04%、42.76%和12.15%,FM 处理的净光合速率显著分别提高了108.33%、148.15%和91.71%。 在不同As 浓度下,FM 处理的净光合速率均显著高于RI 处理。 上述结果表明, 虽然AMF 可显著提高棉花叶片的净光合速率,但在As 的毒害作用下,其光合作用依旧会随As 胁迫的加重而降低,且FM 接种表现出更强的促进作用。
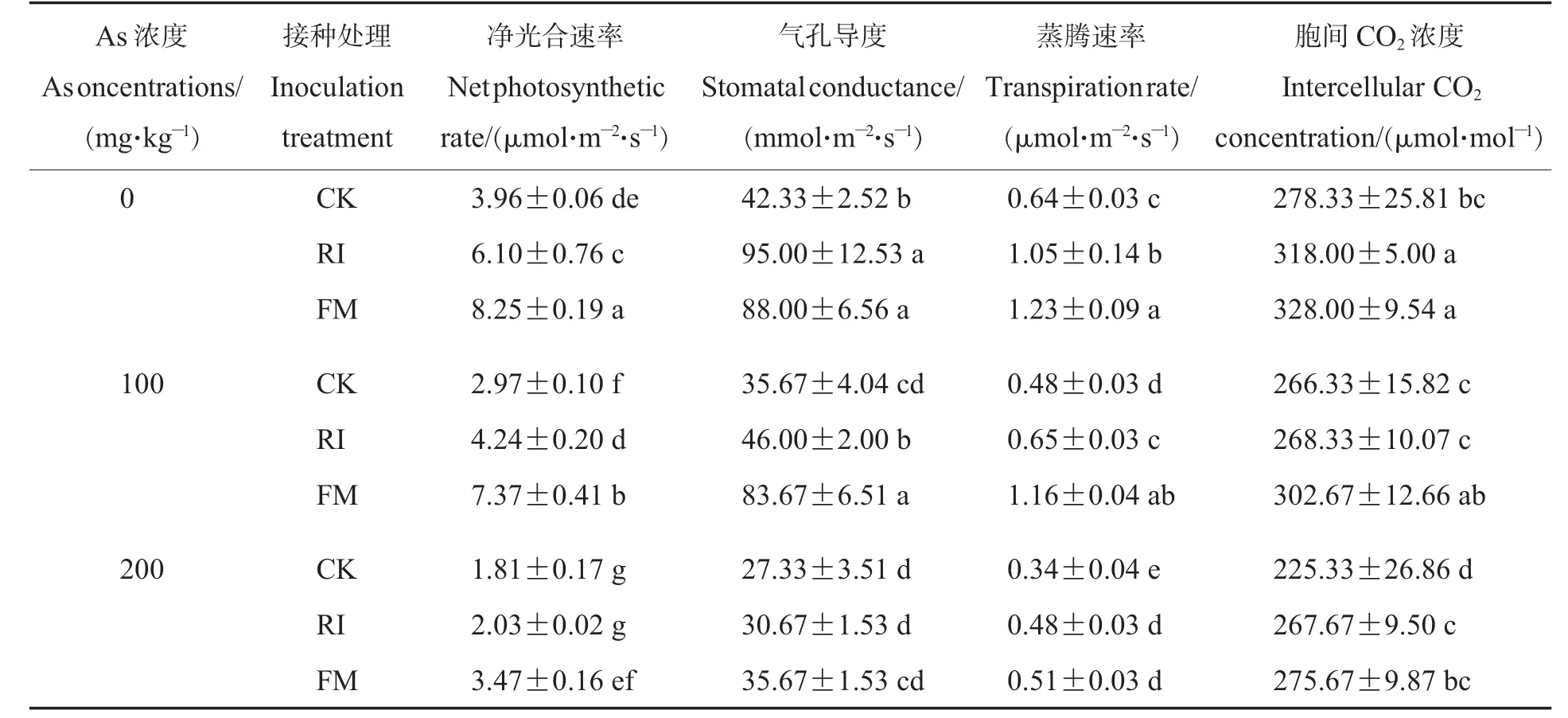
表2 不同As 浓度下AMF 接种对棉花气体交换参数的影响Table 2 Effects of AMF inoculation on gas exchange parameters of cotton under different As concentrations
蒸腾速率、胞间CO2浓度与净光合速率的变化趋势一致,As 胁迫可显著降低其参数值,AMF接种能减轻这种胁迫作用。 气孔导度也有相似的趋势,不同的是,无外源As 处理时,与CK 相比,RI 处理的气孔导度显著提升了124.43%, 较FM的增幅(107.89%)更高;但在100、200 mg·kg-1As胁迫下,FM 处理对气孔导度的提升程度更高,分别为134.57%和30.52%。 以上结果表明,As 可对棉花的光合系统造成一定的损伤,减弱其光合能力;AMF 接种则可缓解As 对光合作用的危害,从而提升棉花耐As 能力。 且FM 处理的缓解能力较RI 处理的更强。
2.3 As 胁迫下AMF 接种对棉花叶绿素荧光参数的影响
由图1 可知,棉花最小荧光与最大荧光参数变化趋势相反,随着As 浓度增加,最小荧光参数逐渐增大, 最大荧光参数逐渐减小。 0、200 mg·kg-1As 浓度下,2 种AMF 接种处理均使最大荧光参数显著提升。而不同As 浓度下,各接种处理间最小荧光参数均无显著差异。

图1 不同As 浓度下AMF 接种对棉花叶绿素荧光参数的影响Fig. 1 Effect of AMF inoculation on chlorophyll fluorescence parameters of cotton under different As concentrations
实际光合效率和最大光化学量子产量变化趋势一致,二者均随As 浓度的增加而降低;不同As 浓度下,与CK 相比,AMF 接种可提高实际光合效率和最大光化学量子产量。 外源As 浓度为0、100、200 mg·kg-1时, 与CK 相比,RI 处理分别使实际光合效率显著提升11.44%、25.83%和87.90%, 最大光化学量子产量分别提高9.72%、1.35%和111.59%; 相对应地FM 接种处理则使实际光合效率分别显著提升19.63%、52.40%和153.23%,最大光化学量子产量分别显著提高10.29%、8.25%和105.96%。 这表明As 胁迫会损伤棉花叶片光系统II(PSⅡ),阻碍叶绿体
内电子链传递,而AMF 接种可降低As 对光合系统的毒害作用, 较CK 处理有效提升光合效率,且FM 处理下光合速率的增幅较RI 的更高。
光化学淬灭系数随着As 浓度增加而减小,非光化学淬灭系数的变化趋势则与之相反。0 mg·kg-1As 浓度下,FM 处理的光化学猝灭系数显著高于CK;100 mg·kg-1As 浓度下, 与CK 相比,FM 处理的光化学猝灭系数显著提高,2 种AMF 处理的非化学猝灭系数均显著降低;200 mg·kg-1As 浓度下,与CK 相比,2 种AMF 处理的光化学猝灭系数均显著提升,非光化学猝灭系数均显著下降;CK 处理的非光化学淬灭系数急剧增长, 较0 mg·kg-1As 浓度下增加了133.47%。 且3 种As 浓度下,FM 处理的光化学猝灭系数均显著高于RI 处理;100、200 mg·kg-1As 浓度下,FM 处理的非光化学猝灭系数均显著低于RI 处理。 由此可知,As 胁迫会导致棉花PSⅡ电子传递活性降低,而以热形式耗散的光能增加;AMF 接种可重新分配As 胁迫下的激发能,降低非光化学的淬灭能量,从而提高光能利用效率,且FM 的效果强于RI。
2.4 As 胁迫下AMF 接种对棉花叶肉细胞超微结构的影响
透射电镜观察结果(图2)表明:外源As浓度为0 mg·kg-1时,CK 处理的棉花叶肉细胞内叶绿体结构完整, 类囊体片层排列整齐有序;线粒体形态饱满,内部结构清晰完整,腔内突嵴排列紧密。 外源As 浓度为100 mg·kg-1时,CK处理的棉花叶肉细胞内的一些叶绿体形态逐渐肿胀,整体结构遭到破坏,基粒类囊体排列紊乱且分布不均;线粒体内嵴排列松散变形,部分线粒体结构内部有空洞出现, 双层膜结构模糊。200 mg·kg-1As 浓度下, 以CK 处理的棉花叶肉细胞内叶绿体形态发生明显变化, 肿胀加剧,类囊体片层十分模糊,乃至消失;线粒体数目减少或结构溶解, 以致未观察到明显的线粒体结构。此外,随着外源As 胁迫的加重,大部分叶肉细胞空泡化逐渐加剧,液泡周围出现许多电子致密颗粒, 这是As 逐渐渗透进入棉花叶肉细胞并沉积的表现。

图2 不同As 浓度下AMF 接种对棉花叶肉细胞超微结构的影响Fig. 2 Effect of AMF inoculation on ultrastructure of cotton mesophyll cells under different As concentrations
100 mg·kg-1As 胁迫下,RI 和FM 处理的叶绿体形态未发生明显变化,部分基粒片层仍清晰可见; 与CK 相比,AMF 接种处理的线粒体内嵴结构排列较为紧密,双层膜结构清晰可见,未见空洞。外源As 浓度为200 mg·kg-1时,AMF 接种使得叶绿体结构损伤得到一定的缓解,但基粒类囊体片层模糊,FM 处理后,可在棉花叶肉细胞中观察到完整的线粒体结构。同时,FM 处理使得液泡周围电子致密颗粒密度降低, 说明As 在棉花叶肉细胞内的沉积减少。 由此可知,As 胁迫可使棉花叶肉细胞超微结构受损, 而AMF 接种可降低As 对叶绿体、线粒体等超微结构的损伤,并减少As 在棉花叶肉细胞内的积累沉淀, 其中,FM处理缓解棉花叶肉细胞受As 毒害的作用强于RI处理。
3 讨论
As 胁迫下植物光合系统会遭到不同程度的损伤,从而降低光合效率[17]。气孔在调节植物与外界水分和气体的交换中发挥着重要作用,植物张开气孔吸收CO2时也会造成水分蒸腾,因此其调节功能与植物呼吸作用和光合能力密切相关,气孔特征也常作为一种指标来阐明植物在逆境中的适应性对策[6]。 本研究结果表明,随着As 浓度的增加,棉花叶片净光合速率、气孔导度、蒸腾速率及胞间CO2浓度均受到As 的抑制作用而降低,气孔密度也随As 浓度的增大而减小。这是棉花为适应As 胁迫环境, 通过调节气孔形态和数目,降低蒸腾作用,以最低水平的水分散失获取最大光合能力18]。Farquhar 等[19]的研究指出,当净光合速率和胞间CO2浓度变化趋势相同时,净光合速率下降是由气孔因素引起的,因此,As 胁迫下棉花叶片气孔趋于关闭,成为光合速率降低的主要因素,王碧霞等[20]的研究也得出了类似的结果。 本研究发现,与对照处理相比,AMF 接种显著提升了同一As 浓度下的光合参数值, 促进了气孔发育、改变了气孔形态,证明AMF 能够调节棉花叶片的气孔开放程度,进而减小CO2进入细胞的阻力,但同时蒸腾速率也随气孔开放程度的增大而增大,最终导致光合速率升高、光合能力增强, 在一定程度上减轻了As 胁迫对棉花光合作用造成的不利影响。
叶绿素荧光参数与光合作用有着密切的联系,能够反映光合作用受环境变化的影响[21]。最小荧光参数可反映逆境胁迫对植物叶片PSⅡ造成不可逆损伤的程度,最大荧光参数则可反映PSⅡ中的电子传递情况,最大光化学量子产量揭示了植物体对生长环境长期适应的机制,可代表植物光合器官的生理状况[22-23]。 本研究中,随着As 浓度的增加, 最小荧光参数逐渐提高, 在200 mg·kg-1As 处理下达到最大值;最大荧光的变化趋势与之相反;最大光化学量子产量和实际光合效率均随As 浓度增加显著降低。说明As 污染阻碍了棉花PSⅡ中电子传递过程, 使棉花PSⅡ反应中心受损,光合产能下降,这与李荟星[24]的研究结果一致。 此外,光化学淬灭系数在一定程度上反映了PSⅡ的开放程度, 光化学淬灭系数与PSⅡ电子传递活性呈正相关;非光化学淬灭系数表示以热能等形式耗散的光能,二者可反映出光能在植物体内的分配和利用效率[24]。本研究发现随着As浓度的增加,光化学淬灭系数不断降低而非光化学淬灭系数不断升高,说明PSⅡ电子传递活性减小,而以热能形式散失的光能增加。吴敏兰等[25]的研究表明可能是由于As 胁迫使植物体内活性氧和自由基浓度升高而损害了光合系统。 而李荟星[24]研究表明,As 会竞争结合PSⅡ反应中心的D1蛋白,从而阻断从QA到QB的电子传递,造成QA无法进行下一步的电子传递,进而抑制光合效能。 与对照处理相比,AMF 接种后,棉花光合系统受损程度得到有效缓解,最大光化学量子产量与实际光合效率均显著升高,光化学淬灭系数升高的同时非光化学淬灭系数降低。 其原因可能是AMF 提高了PSⅡ反应中心的电子传递活性,优化了As 胁迫下光能的分配, 使非光化学能量淬灭减少,降低热耗散,提高了PSII 反应中心的电子传递能力和叶绿体性能, 从而提高了As 胁迫下棉花光化学效率。此外,研究表明AMF 也可激活棉花的抗氧化系统并调节其渗透系统,抗氧化酶活性的升高也可降低过氧化物对棉花叶片的损伤,同时渗透调节系统加强了叶片对水分的利用效率,这也间接地促进了棉花光合效率的提升[13]。
光合作用是绿色植物生存的基础,也是植物对环境胁迫最为敏感的生理过程。 在植物生长发育过程中,砷离子会在植物体内进行迁移,可影响叶绿体、 线粒体等多个细胞器的组织代谢活动,干扰细胞内相关蛋白合成、呼吸作用和光合作用,造成直接的毒害效应[26]。 线粒体膜上的ATP 酶活性被抑制、钾离子的渗透等因素都会引起线粒体嵴结构改变,导致线粒体内的氧化磷酸化循环被破坏,甚至解体死亡。 本研究中,As 胁迫导致棉花叶肉细胞内叶绿体肿胀变形,打乱了类囊体片层的排列,同时也使线粒体嵴结构改变甚至溶解消失,在细胞质中形成许多电子致密颗粒物质。 由此推测,As 严重影响了光合电子传递链的正常功能,抑制了光能的有效转化。 与对照处理相比,AMF 接种使得细胞内致密电子颗粒密度降低,细胞器损伤程度减轻,200 mg·kg-1As浓度下,FM 处理可在叶肉细胞中观察到线粒体,这也是As 胁迫对棉花叶肉细胞的毒性得到缓解的表现。
4 结论
AMF 接种可增加棉花叶片气孔数量并提高气孔开放程度,减轻叶绿体、线粒体等细胞器的损伤,促进叶绿体内电子链传递,提高其净光合速率,增强As 胁迫下的光合能力,有效减轻As对棉花光合作用的毒害作用。 根内根孢囊霉和摩西斗管囊霉这2 种AMF 对棉花耐As 性均表现出良好且一致的促进作用,但摩西斗管囊霉表现出更强的效应。 本研究为AMF 可能缓解棉花的光合系统受As 毒害机制提供了一定的理论支撑。
猜你喜欢
生态学报(2024年1期)2024-01-25 07:05:16
干旱地区农业研究(2022年1期)2022-01-28 04:51:30
蔬菜(2022年12期)2022-01-01 11:32:10
云南化工(2021年9期)2021-12-21 07:44:10
装备制造技术(2019年12期)2019-12-25 03:06:52
科技创新与品牌(2019年12期)2019-02-06 04:03:29
农村百事通(2019年1期)2019-01-30 13:36:34
中国铸造装备与技术(2015年5期)2015-12-10 10:23:33
农家顾问(2014年11期)2014-12-01 13:29:34
影像科学与光化学(2014年5期)2014-03-11 16:03:23
