
2020 年我国部分地区猪繁殖与呼吸综合征分子流行病学调查
2022-08-06 10:16梁俊超刘红祥郝尧光王宗升王辉之杜元钊
中国动物检疫 2022年8期
梁俊超,赵 翔,刘红祥,袁 飞,李 坤,郝尧光,王宗升,王辉之,刘 东,杜元钊
(1.吉林大学动物医学学院人兽共患病研究所,人兽共患病研究教育部重点实验室,吉林长春 130062;2.青岛易邦生物工程有限公司,动物基因工程疫苗国家重点实验室,山东青岛 266000;3.青岛市市北区疾病预防控制中心,山东青岛 266000)
猪繁殖与呼吸综合征病毒(porcine reproductive and respiratory syndrome virus,PRRSV)可引发猪繁殖与呼吸综合征(porcine reproductive and respiratory syndrome,PRRS),俗称“蓝耳病”。PRRS 是一种高度接触性传染病,导致猪群出现繁殖障碍、免疫抑制及呼吸道疾病等问题,引发猪只死淘率增高,对猪场生产造成严重影响。PRRSV 是一种有囊膜的单股正链RNA 病毒,基因组大小15 kb 左右,含开放阅读框(open reading frame,ORF)至少11 个,编码结构蛋白8 种,非结构蛋白至少16 种。
PRRSV 可分为两大亚群,分别为PRRSV1(欧洲株)和PRRSV2(美洲株)[1]。据ORF5 基因序列分析,PRRSV2 可分为9 个谱系,其中在我国流行的主要分布在4 个谱系,分别为谱系1(代表毒株为NADC30 及MN184B)、谱系3(代表毒株为GM2)、谱系5(代表毒株为VR2332)及谱系8(代表毒株为CH-1a、BJ0706 及JXA1)[2]。2012年前,在我国流行的PRRSV 毒株为经典美洲株(代表毒株CH-1a)和高致病性毒株(HP-PRRSV,代表毒株JXA1),其中2006 年以后HP-PRRSV 为主要流行毒株,其致病力比经典PRRSV 明显增强[3-4]。2012 年NADC30-like PRRSV 被报道。该类毒株易重组,彼此间同源性低,且临床致病性具有明显差异。2017 年后,我国多省发现NADC34-like PRRSV[5-6]。当前,我国主要流行毒株已逐渐 由HP-PRRSV 转 为NADC30-like PRRSV[7]。NADC30-like PRRSV 毒株与美国1-7-4 分支毒株相似,而1-7-4 分支毒株已对美国及秘鲁的养猪业造成重大影响。2020 年美国报道一种美洲型HPPRRSV 毒株,被命名为ORF5 1-4-4 Lineage 1C 变异株(简写为PRRSV 1-4-4)[8]。该毒株传播力强,致死率高,已严重影响美国养猪业。NADC30-like PRRSV、NADC34-like PRRSV 及PRRSV 1-4-4 均为PRRSV2(美洲株),且属于谱系1[9]。
本试验收集2020 年我国部分地区PRRSV 感染临床可疑样品,通过ORF5 基因测序比对分析,了解国内PRRS 的分子流行病学变化情况,旨为PRRS 防控提供依据。
1 材料与方法
1.1 病料收集处理
2020 年从山东、河北、江苏、安徽等7 个省,采集570 份疑似PRRSV 感染的病料(胎衣和组织)、血液(全血和脐带血)及精液,经实验室处理后,按照文献[10]的RT-PCR 方法进行抗原检测。
检测用ORF5 基因引物如下:
上 游:CATTTCATGACACCTGAGACCA;下游:AGAGCATATATCATCACTGGCG。
RT-PCR 扩增条件如下:
94 ℃预变性2 min;94 ℃变性15 s,54 ℃退火30 s,68 ℃延伸1 min,32 个循环;68 ℃延伸10 min。
1.2 主要试剂
RNA 提取试剂盒,购于TAKARA 公司;高保真反转录试剂盒,购于罗氏公司;LATaqDNA聚合酶,购于宝生物工程(大连)有限公司。
1.3 PRRSV GP5 测序
将PRRSV RT-PCR 阳性产物送至生工生物工程(上海)股份有限公司进行测序。
1.4 核苷酸进化分析
通过软件DNAstar Lasergene,对送检测序PRRSV ORF5 基因进行序列同源性比对分析;利用MEGA 6.0 软件,对ORF5 基因序列进行遗传进化分析。
1.5 氨基酸进化分析
通过软件DNAstar Lasergene,对送检测序PRRSV GP5 蛋白氨基酸位点进行分析,并基于ORF5 基因编码序列,通过NetNGlyc l.0 Sever 在线软件(http://www.cbs.dtu.dk/services/NetNGlyc/),预测并分析相应氨基酸序列的糖基化位点。
2 结果与分析
2.1 RT-PCR 检测
将2020 年采集的570 份疑似PRRSV 感染病料进行RT-PCR 检测,目的条带大小为603 bp。结果(图1、表1)显示:从送检的疑似PRRSV 感染病料中检出PRRSV 病原阳性135 份,阳性检出率为23.68%,其中保育猪病料的阳性检出率最高,为32.24%,哺乳猪次之,为19.09%。

表1 2020 年不同猪群送检病料的PRRSV 阳性检出结果统计
2.2 ORF5 基因同源性比对分析
随机送检60 份PRRSV 阳性样品的PCR 产物进行ORF5 基因测序,经比对后确认得到60个603 bp 的ORF5基因序列。将 这60 个ORF5基因序列按照“送检年月-样品编号-送检省份缩写”进行编号。基因序列对应省份及数量分布见表2。60 个ORF5 基因序列与不同谱系标准毒株序列通过DNAstar 软件的比对结果(表3、图2)显示,60 个ORF5 基因序列彼此间同源性为79.9%~100%,与国内流行的PRRSV2 4 个谱系标准毒株同源性为81.3~99.7%,与PRRSV1 标准毒株同源性仅为60.7%~63.8%,表明这60 个PRRSV毒株均为PRRSV2。
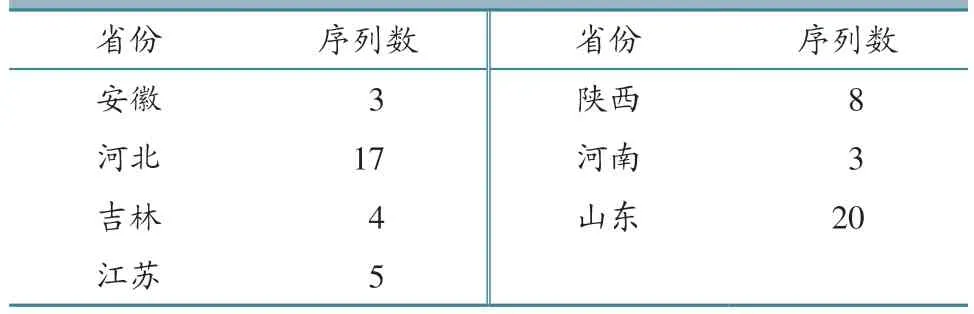
表2 60 个PRRSV 基因序列区域分布及数量 单位:个

表3 60 个PRRSV ORF5 基因序列与标准毒株同源性对比结果 %
2.3 ORF5 基因遗传进化分析
60 个PRRSV 的ORF5 基因序列通过MEGA 6.0 软件绘制遗传进化树。结果(图3)显示,31个PRRSV 属于谱系1,占比达51.67%,26 个属于谱系8,占比达43.33%。此外,1 个PRRSV 属于谱系3,1 个属于谱系5,而剩余1 个(20200624-262-JS)介于谱系5 和谱系8(可能为疑似PRRSV重组毒株,有待进一步研究),占比均为1.67%。
2.4 GP5 蛋白氨基酸位点变异分析
将60 个PRRSV GP5 蛋白氨基酸序列通过DNAstar 软件与相应谱系的标准毒株进行比对。结果(图4)显示,60 个PRRSV GP5 蛋白氨基酸以点突变为主,其中10 个PRRSV 分别在33 aa 或34 aa 处存在缺失突变;信号肽区域(1~25 aa)以及27~39、57~61、120~125、167~170 及189~196 aa等6 个区域的氨基酸点突变集中程度较高。
60个PRRSV GP5蛋白的2个非中和表位(27~30和180~197 aa)、1 个中和表位(37~45 aa)及2 个潜在毒力位点(R13和R151)氨基酸变化及相应谱系占比详见表4。从表4 可知:60 个PRRSV 在非中和表位(180~197 aa)突变比例最高,平均为96.67%;其次为R13潜在毒力位点,突变比例平均为53.33%;非中和表位(27~30 aa)突变比例最低,平均为35.00%;而中和表位及R151潜在毒力位点也发生不同程度突变,平均分别为51.67%和48.33%。

表4 60 个PRRSV 的GP5 蛋白重要氨基酸位点变化及相应谱系占比 %
2.5 GP5 蛋白潜在N-糖基化位点变异分析
通过NetNGlyc l.0 Sever 在线软件,对60 个PRRSV GP5 蛋白氨基酸序列的N-糖基化位点进行预测分析。结果(表5)显示,谱系1 参考毒株NADC30、PRRSV 1-4-4 和NADC34 潜在糖基化位点数量分别为3、4 和5 个,而潜在糖基化位点数量为4 个的样品占比最高,为48.39%(15/31);谱系3、谱系5 参考毒株与其相应样品PRRSV 潜在糖基化位点数量一致;20200624-262-JS 潜在糖基化位点数量为3 个;谱系8 参考毒株CH-1a和JXA1 潜在糖基化位点数量分别为3 和4 个,潜在糖基化位点数量为4 个的样品占比最高,为73.08%(19/26)。60 个PRRSV 的潜在N-糖基化位点位置主要集中在34~35、43~44 和50~51 aa,占比分别为65.00%(39/60)、96.67%(58/60)和100%(60/60)。

表5 60 个PRRSV 的GP5 蛋白潜在N-糖基化位点数量及位置分布 单位:个
3 讨论
PRRS 是猪场防控的重要猪病。我国猪群PRRSV 感染较普遍,田间流行毒株复杂多样[11]。从PRRSV 的RT-PCR 抗原检测结果可知,疑似发病场的PRRSV 阳性检出率为23.68%,其中保育猪群最高(32.24%),这与猪场临床发病情况相符,主要与保育阶段猪只免疫力低、应激因素多以及环境因素等直接相关[12]。
通过60 个PRRSV 的ORF5 基因同源性比对结果可知,不同PRRSV 毒株间同源性差异较大,谱系1 中PRRSV 与NADC30、NADC34及PRRSV 1-4-4 的同源性为87.1%~97.7%,与其他谱系参考毒株同源性<88.2%;谱系8 中PRRSV 与JXA1、HuN4 及TJ PRRSV 的同源性为97.2%~99.7%,与其他谱系参考毒株同源性<89.1%。由此可知,不同谱系PRRSV 的ORF5 基因同源性较低,而PRRSV 同源性高低对PRRSV疫苗保护效果产生影响[13]。
通过进化树结果分析发现:59 个PRRSV分别属于谱系1、谱系3、谱系5 和谱系8,剩余1 个介于谱系5 与谱系8;谱系1 占比最高,为51.67%,其中30 个为NADC30-like PRRSV,仅1 个河北样品为NADC34-like PRRSV,未发现PRRSV 1-4-4 及MN184B PRRSV。结果表明,国内流行的PRRSV 毒株复杂多样,但以NADC30-like PRRSV 为主流流行毒株。该结果与张洪亮等[5]、杨汉春等[14]的报道相符。
N 端胞外域27~41 aa 区域和C 末端180~197 aa 区域是PRRSV GP5 蛋白2 个重要抗原表位相关区域[15]。相关研究[16]显示,GP5 蛋白的C端抗原表位在维持其蛋白构象方面具有重要作用。GP5 蛋白具有3 个重要抗原表位,分别为中和抗原表位37~45 aa、非中和抗原表位(又称诱饵表位)27~30 aa 和非中和抗原表位180~197 aa。非中和抗原表位27~30 aa 和中和抗原表位37~45 aa 在中和抗体产生过程具有关键作用,其中38 和42~44 aa是病毒中和表位的识别位点,39~41 aa 是病毒中和表位的结合位点[17]。通过氨基酸比对结果分析发现,谱系1 中PRRSV 的中和表位及2 个非中和表位均有突变,其中中和表位突变率低,为9.68%,2 个非中和表位27~30 和180~197 aa 突变比例相比中和表位更高,180~197 aa 表位突变比例高达96.77%,这可能对中和抗体产生及GP5 蛋白构象产生影响,导致PRRSV 免疫失败[18-19]。
谱系3 中PRRSV 的3 个表位均发生突变;谱系5 中PRRSV 及20200624-262-JS 各有2 个 表位发生突变,其中180~197 aa 表位均发生突变;谱系8 中的PRRSV 只有中和表位及非中和表位180~197 aa 发生突变,且突变比例均为100%,其中中和表位突变位点主要为39 aa,而该位点是病毒中和表位的结合位点,这或对GP5 蛋白构象及中和抗体对病毒的结合能力产生影响,也可能对蛋白免疫原性产生影响,更利于病毒免疫逃避,影响现有疫苗免疫保护效果[20]。
据相关研究[21],GP5 蛋白R13和R151与PRRSV 毒力相关,是潜在毒力位点。60 个PRRSV仅20200624-262-JS 的2 个位点均未发生突变,其余59 个PRRSV 均发生不同程度突变,尤其谱系1中PRRSV 的R13位点突变比例高达96.77%,这对NADC30-like PRRSV 的毒力影响程度有待进一步研究。GP5 蛋白具有多个糖基化位点,这些糖基化位点严重影响病毒蛋白的正确折叠及生物学特性,可形成糖基屏蔽效应有利于病毒免疫逃避及持续性感染[21-22]。
通过N-糖基化位点预测发现,60 个PRRSV糖基化位点数量及分布相比各自谱系参考毒株均存在不同程度差异,这也表明不同PRRSV 免疫逃避能力存在差异,因而加重了PRRS 防控的复杂程度。
疫苗免疫是PRRS 防控的重要手段。我国目前商品化的PRRSV 疫苗均为PRRSV2 且属于谱系8,与当下田间流行的NADC30-like PRRSV 野毒株同源性较低。PRRSV 弱毒活苗对不同野毒株具有交叉保护性,其与野毒株同源性高低可能与保护率无关。Opriessnig 等[23]在猪群免疫PRRSV弱毒活苗后,用与疫苗毒同源性仅为76%的高毒力PRRSV 攻毒,发现免疫组比未免疫组肺脏损伤率低37.8%,仅为4.2%。据报道[24],TJM-F92 等PRRSV 弱毒活苗与NADC-like PRRSV 同源性虽大多小于89%,但对NADC-like PRRSV 仍可提供有效临床保护,减轻临床症状,缩短发病时间,改善平均日增重。由此可知,PRRSV 弱毒活疫苗对同源性低的野毒株也可提供一定的交叉保护,对免疫猪群可提供有效的临床保护,改善猪群体况。总之,PRRS 需要综合防控,猪场除可选用安全有效的PRRSV 弱毒活疫苗外,猪场生物安全及饲养管理也具有重要作用。
猜你喜欢
传染病信息(2022年4期)2022-09-17
艺术品鉴(2022年16期)2022-07-09
中国高原医学与生物学杂志(2022年3期)2022-06-22
复旦学报(医学版)(2021年5期)2021-10-13
北方论丛(2021年2期)2021-05-22
景德镇陶瓷(2021年1期)2021-03-24
智慧健康(2021年2期)2021-03-15
三农资讯半月报(2020年8期)2020-05-13
当代陕西(2019年19期)2019-11-23
温州医科大学学报(2019年4期)2019-04-28
