
培养温度、光照及贮藏时间对黑木耳菌包中两种酶活性的影响*
2022-08-04 09:43高智涛王洪刚范东茹李金禹李占君董傲宇
中国食用菌 2022年7期
高智涛,王洪刚,范东茹,李金禹,李占君,张 巍,郭 兴,董傲宇
(黑龙江省林业科学院伊春分院,黑龙江 伊春 153000)
黑木耳(Auricularia auricula) 富含蛋白质、粗纤维,是中国广泛人工栽培的食用菌品种,黑木耳栽培也成为一些地区的支柱产业之一。近年来,随着人们生活水平的提高,对黑木耳的消费需求也日渐也增长,黑木耳总产量随之快速增长。但近年来,因菌株退化、老化和不利变异导致的栽培性状难以预测等问题,造成了黑木耳栽培特性、栽培产量的不稳定[1],从而严重影响了黑木耳产业的发展。
食用菌生长发育所需的条件主要包括营养条件和环境条件。营养条件主要包括碳源、氮源、碳氮比、水分、酸碱度和矿质元素含量等;环境条件主要包括光照、温度、湿度、风速、CO2和O2等,其中光照与温度是关键的环境因素[2]。若食用菌菌丝在生长发育过程中受到不利因素影响,后续的栽培同样会受到影响,最终导致菇农在生产中遭受损失。有研究表明,侧耳(Pleurotus sp.) 和金针菇(Flammulina velutipes) 的菌丝中羧甲基纤维素酶的活力与其子实体产量成正相关[3]。同样,在黑木耳菌种培养过程中可以通过测定酶活性研究菌种的酶活性变化规律,检测菌种分解纤维素、木质素等养分的能力,可在一定程度上检测出菌种老化、退化以及不利变异等情况[4-5]。如此可杜绝劣质菌种投入生产,为黑木耳栽培生产提供科学、系统的理论依据。因此,研究黑木耳胞外酶在栽培过程中的活性变化规律对判断优劣十分重要。
此次通过对黑木耳菌包培养期间温度、光照及满袋后冷冻时间进行干预,测定各因素对菌种多酚氧化酶(Polyphenol oxidase,PPO) 和羧甲基纤维素酶(Carboxymethyl cellulase,CMCase) 活性的影响,探讨黑木耳菌丝生长过程中酶活性变化的规律性,为判断黑木耳菌包品质是否优良提供参考。
1 材料与方法
1.1 供试菌包
黑木耳三级菌包(菌种为“黑尊二号”,已接种培养15 d),伊春黑尊公司。
1.2 温度处理
将黑木耳菌包分别置于20 ℃、25 ℃、30 ℃、35 ℃、40 ℃的恒温培养箱中,培养至菌丝满袋,进行酶活测定。
1.3 光照处理
将黑木耳菌包置于25 ℃恒温培养箱中,培养箱内搭设红光、蓝光、紫光、黄光4 种LED 光源,分别在不同光源照射下培养至菌丝满袋;黑暗培养作为对照(CK1);进行酶活测定。
1.4 低温处理
将25 ℃培养至菌丝满袋,且经后熟培养的菌包置于-20 ℃进行冷冻,分别在冷冻10 d、20 d、30 d、40 d、50 d 时取出菌包,自然解冻;未经冷冻的菌包作为对照(CK2);进行酶活测定。
1.5 酶活测定
1.5.1 粗酶液提取
取长满黑木耳菌丝的培养料基质5 g,于20 ℃~25 ℃高速匀浆3 min,定容至250 mL;然后转移至500 mL 三角烧瓶中,25 ℃、120 r·min-1摇床培养3 h;培养结束后,4 ℃、4 000 r·min-1离心5 min;获得的上清液为粗酶液。
1.5.2 酶活性的测定及计算
参照王玉江等[6]的方法测定粗酶液中多PPO 和CMCase 的活性。PPO 单位定义为:以每毫升粗酶液与底物作用30 min 内改变0.1 个光度值为1 活力单位(U)。CMCase 单位定义为:以每毫升粗酶液中的酶量与底物作用30 min 释放出1 mg 葡萄糖为1 活力单位(U)。
2 结果与分析
2.1 不同温度对酶活性的影响
酶活性的变化可以反映出菌丝分解利用木质素的能力,当对极致营养物质的利用率高时可促进菌丝生长;一定范围内升高培养温度虽然会提高菌丝生长速度,但也会增大菌包内水分蒸腾速率,导致培养环境的湿度增大,而在温度和湿度较高的培养条件下极易感染霉菌等杂菌,会增大菌包被污染的风险[7]。因此,目前黑木耳常用的栽培管理方法中,在刚接种三级种时设置的培养温度略高,约为25 ℃~30 ℃,而后续的培养保持在25 ℃左右[8]。不同培养温度对黑木耳菌包中酶活性的影响结果见图1。
如图1 所示,黑木耳菌包PPO 酶活性在15 ℃~40 ℃范围内随温度升高呈上升趋势;其中,15 ℃~35 ℃时,酶活性升高趋势明显、差异性较大;35 ℃~40 ℃时,酶活性变化趋于平缓,变化差异性较小。CMCase 酶活性在15 ℃~40 ℃范围内随温度升高呈上升趋势;其中,20 ℃~35 ℃时,酶活性升高趋势明显,差异性较大;15 ℃~20 ℃及35 ℃~40 ℃的温度区间内,酶活性变化趋于平缓,差异性较小。因此,按照黑木耳栽培温度管理方法,可保证PPO 和CMCase 酶活性处于较高水平。
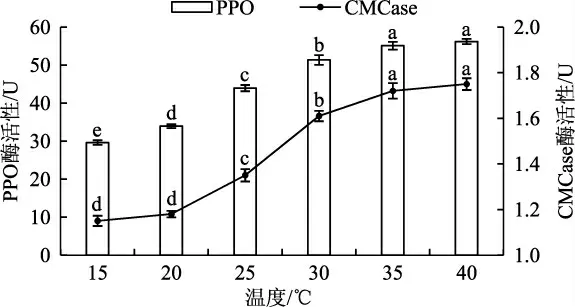
图1 不同培养温度对黑木耳菌包中PPO、CMCase变化情况影响Fig.1 Effects of different culture temperatures on the changes of PPO and CMCase in the Auricularia auricula bags
2.2 不同光源对酶活性的影响
不同颜色的光源对黑木耳菌包中酶活性的影响结果见图2。

图2 不同光源对黑木耳菌包中PPO、CMCase 变化情况影响Fig.2 Effects of different light sources on the changes of PPO and CMCase in the Auricularia auricula bags
如图2 所示,不同光源处理对黑木耳菌包中PPO、CMCase 的酶活性均有一定影响。蓝色光源下黑木耳菌包中的PPO 酶活性较高,黄色、红色光源下的PPO 酶活性与对照组(CK1) 黑暗培养相比,变化不显著,紫色光培养下PPO 酶活性最低。紫色光源下黑木耳菌包CMCase 酶活性较高,但与蓝色光源下培养的黑木耳菌包的CMCase 酶活力变化的差异性不显著,红色、黄色光源与黑暗条件下培养的CMCase 酶活性变化的差异性不显著。由此可知,光照对食用菌菌丝生长具有一定的影响,蓝光、紫光均可提升2 种酶的活性;但在光照培养过程中也发现有少部分菌包形成耳芽,这会对后续栽培产生影响。
2.3 不同冷冻时长对酶活性的影响
菌丝满袋且经后熟培养后,不同的冷冻时间对黑木耳菌包中酶活性的影响结果见图3。
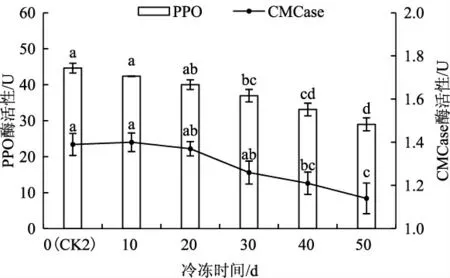
图3 不同冷冻时间对黑木耳菌包中PPO、CMCase变化情况影响Fig.3 Effects of different freezing times on the changes of PPO and CMCase in the of Auricularia auricula bags
如图3 所示,PPO、CMCase 酶活性随着冷冻天数的增加而逐渐降低。分析于-20 ℃下冷冻时黑木耳菌包中的PPO 酶活性,冷冻10 d 与未经冷冻(CK2) 二者之间无差异性;冷冻20 d 与CK2 及冷冻10 d 相比,差异性较小;冷冻30 d 及以上差异性较显著,且酶活性呈下降趋势。分析CMCase 酶活性发现,冷冻10 d 与CK2 二者之间也无差异性;冷冻20 d、30 d 与CK2、冷冻10 d 的相比,差异性较小;冷冻40 d、50 d 时差异性极为显著。冷冻10 d时CMCase 活性略微增长,这与CMCase 特性有关:CMCase 是一种诱导酶,随着菌丝体生长发育,菌丝细胞分泌的胞外CMCase 会逐渐增多,因此,CMCase 活性随着培养时间逐渐增加[9]。由此可知,黑木耳菌包经短时间的低温储藏并不会对PPO、CMCase产生较大影响,但长时间的低温储藏会降低PPO、CMCase 的酶活性。
3 结论与讨论
黑木耳栽培的整个过程中,菌丝生长主要靠分解栽培基质中的纤维素、半纤维素、木质素等获取营养,虽然不同品种菌株的分解能力不同,但均需在酶的作用下才能进行。PPO、CMCase 是与木质素等大分子物质降解有关的酶,是伴随黑木耳生长整个过程中的关键指标之一,酶活性会影响菌丝长势及产量[5]。
试验结果显示,菌包在培养过程中PPO、CMCase 活性会随着温度变化而发生变化,这与大多数酶活性变化规律相同[10]。黑木耳菌包培养过程中,在25 ℃~35 ℃温度条件下PPO、CMCase 酶活性升高趋势明显,酶活性的升高说明菌种对木质素的分解活性较高。现阶段进行黑木耳菌包培养时要求温度控制在25 ℃左右,但若通风不良、菌包堆放密集,呼吸产生的热量致使局部温度达到35 ℃以上;虽然一定程度上升高温度可提高酶活性,但同时也会对黑木耳菌丝造成伤害,导致菌丝细胞组织结构损伤和生理代谢紊乱[11]。
培养阶段使用不同光源照射会影响菌包中PPO、CMCase 酶活性,试验结果与红光、蓝光适合黑木耳菌丝的生长的结论相符[12]。虽然光照会增加2 种酶的活性,但也会导致不规则耳芽的提前出现,影响栽培出菇效益。因此,不建议在菌包培养阶段使用光源刺激,而应在出菇期间进行合理光源利用。
黑木耳菌包在满袋后熟后,若因培养室空间不足、需要转移或运输等情况发生时,有时会遇到极端天气,菌包受冻也经常发生。通过试验发现,随着冷冻时间延长,PPO、CMCase 酶活性降低十分明显,这与黑木耳菌丝生理特性相吻合[13]。当冷冻时长不超过20 d,并不会对这2 种酶产生较大影响,但时间过长则会对黑木耳菌丝造成损害。因此建议生产黑木耳三级种时,菌包成熟后若需储存于0 ℃以下的环境中,放置时间不可超过20 d。
食用菌生产中通常以菌包外观判断菌包品质,但因栽培过程中同品种食用菌的生产条件相同,有时无法仅从观察菌包外观判断其在培养过程中是否受到其他因素损害。因此,通过检测菌包中酶活性的变化,可以反映菌包在培养过程中菌丝生长的活性状况,从而为判断黑木耳菌包品质提供更准确的参考。
猜你喜欢
农业与技术(2022年19期)2022-10-17
河南农业科学(2022年3期)2022-05-27
陕西农业科学(2022年2期)2022-03-28
阅读(中年级)(2021年5期)2021-08-23
农民致富之友(2019年10期)2019-05-22
新农村(2018年12期)2018-08-24
农村农业农民·B版(2017年11期)2017-12-06
农家科技(2017年3期)2017-06-26
读写算·小学低年级(2009年12期)2009-12-28
农业工程技术·温室园艺(2009年11期)2009-05-17
