
绒山羊角蛋白关联蛋白基因家族研究进展
2022-08-04 06:46何兆华罗玉柱王继卿李少斌
家畜生态学报 2022年6期
何兆华,罗玉柱,王继卿,胡 江,刘 秀,李少斌
(甘肃农业大学 动物科学技术学院,甘肃省草食动物生物技术重点实验室,甘肃 兰州 730070)
羊绒是一种具有独特特性的天然纤维,具有轻、软、暖等特点,常被用作高档纺织原料,提高绒品质和产量是绒山羊生产的首要目标。角蛋白关联蛋白(keratin-associated proteins,KAPs)由角蛋白关联蛋白基因家族()编码,其作为羊绒纤维的主要结构蛋白,对羊绒纤维的物理性质起着决定性作用。KAPs通常含有丰富的半胱氨酸或甘氨酸和酪氨酸,可以通过广泛的二硫键交联形成一个半刚性基质,进而嵌入到位于角蛋白中间丝蛋白的半胱氨酸残基中,形成羊毛纤维的主体结构,最后通过将角蛋白中间丝(keratin intermediate filaments, KIFs)组装成更大的纤维束来决定羊毛特性。毛囊分化的进程以一系列不同的和特异性角蛋白基因的连续激活为特征,这两类基因的结构完整性和严格调控对于毛发的正常生长至关重要。
通过比较不同哺乳动物的角蛋白(Keratin)和KAPs,发现角蛋白在结构和功能上相对保守,而KAPs则表现出高度分化的特征,这与哺乳动物毛发表型的多样性一致,充分说明KAPs与毛发表型紧密相关。随着分子生物学的发展,的研究不断深入。本文综述了绒山羊基因的种类、染色体定位、核苷酸多态性以及组织表达等方面相关研究进展,以期为后续研究提供参考。
1 山羊KAPs的种类和结构特性
根据氨基酸成分,KAPs可被分为3大类,即高硫KAPs(HS,半胱氨酸≤30%),超高硫KAPs(UHS,半胱氨酸>30%)和高甘氨酸/酪氨酸KAPs(HGT,甘氨酸和酪氨酸35%~60%)。由于基因变异属于非致死性突变,只会引起毛发蛋白的变异,最终造就了庞大的KAPs家族。
目前,在人类和绵羊中发现至少有25个亚家族的80个KAPs和19个亚家族的36个KAPs。然而,在山羊中只发现了来自15个亚家族的21个KAPs(表1)。可以预见,随着研究的不断深入,将会有更多的山羊KAPs被鉴别。
1.1 山羊高硫KAP的种类与特征
目前,在山羊中已知的高硫KAP包括9个亚家族的12个成员,除KAP1有3个成员(KAP1-1、KAP1-3和KAP1-4)及KAP13有2个成员(KAP13-1和KAP13-3)外,其余7个亚家族(KAP3、KAP11、KAP15、KAP24、KAP26、KAP27、KAP28)均只有1个成员(表1)。在12个成员中,5个为酸性蛋白,7个为碱性蛋白。半胱氨酸含量8.2%~30%,与其他HS-KAPs相比,KAP24-1和KAP26-1的半胱氨酸含量异常低,分别为8.9%和8.2%,这意味着目前认为的角蛋白关联蛋白都有较高含量的半胱氨酸或者富含甘氨酸/络氨酸的观点可能需要修正。另外,KAP11-1、KAP13-3、KAP15-1、KAP26-1、KAP27-1及KAP28-1均含有较高的丝氨酸,由于丝氨酸残基可以翻译后磷酸化,可能与蛋白质的磷酸化有关,而磷酸化会影响角蛋白的组成,因而高水平丝氨酸的生物学意义值得进一步研究。
1.2 山羊超高硫KAP的种类与特征
目前,在山羊中发现的超高硫KAP只有KAP9亚家族中KAP9-2,编码191个氨基酸,等电点为8.03,为弱碱性蛋白,该蛋白质不含任何Trp残基。KAP9-2可能在连接中间丝角蛋白中起重要作用。
1.3 山羊高甘氨酸/酪氨酸KAP的种类与特征
目前,山羊已知的高甘氨酸/酪氨酸KAP包括5个亚家族的8个成员,其中KAP6、KAP8和KAP20亚家族分别包括2个成员(KAP6-1、KAP6-3、KAP8-1、KAP8-2、KAP20-1和KAP20-2),其余2个亚家族(KAP7和KAP22)分别只包含1个成员(表1)。8个成员中,2个为酸性蛋白,6个为碱性蛋白,甘氨酸和酪氨酸含量35%~60%。其中,KAP6蛋白由甘氨酸酪氨酸和甘氨酸酪氨酸甘氨酸的重复单位组成,该蛋白质不含任何Trp残基,序列的N末端是M(Met)。相比其他HGT-KAP,KAP8-2含有高含量的天冬氨酸(3.2%)和谷氨酸(1.6%),山羊KAP20-2序列的甘氨酸和酪氨酸含量(53.23~54.84%)低于人KAP20-2(61.5%)和小鼠KAP20-2(63.3%),人和小鼠的KAP20-2包含M(I/C)YY(R/S)(G/N)YY和RY(W/-)(S/-)(Y/C)GFY的保守氨基末端和羧基末端序列,而山羊KAP20-2并不精确地包含这些保守序列。
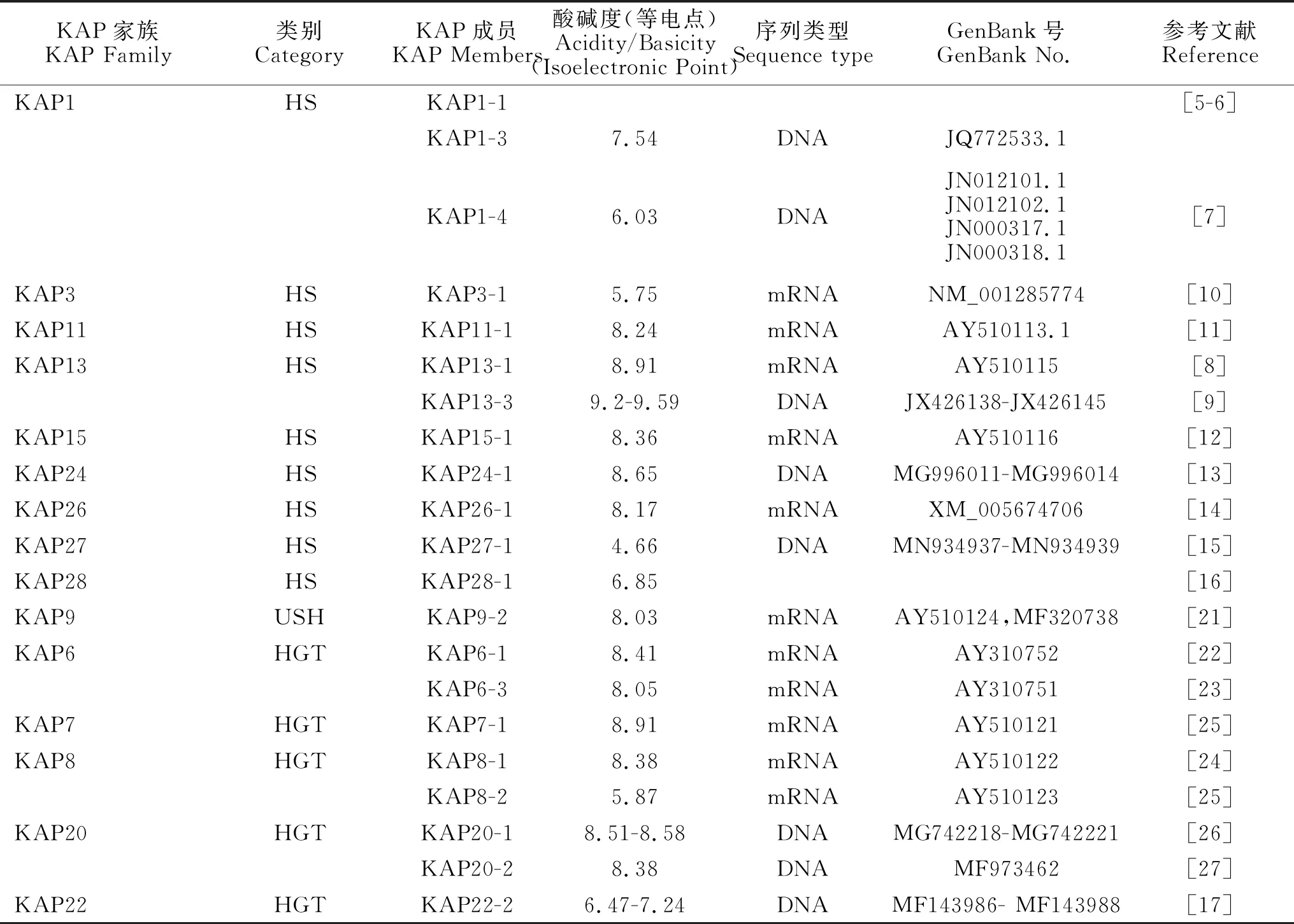
表1 山羊角蛋白相关蛋白(KAP)及其序列信息
2 山羊KRTAPs家族基因的染色体定位
迄今为止,已发现的山羊基因大部分位于1号染色体,包括11-1、7-1、8-1、8-2、20-2、22-2、6-3、6-1、20-1、15-1、13-1和13-3、27-1、26-1、28-1 和24-1;位于19号染色体的有3-1、1-4、1-3和9-2。山羊基因染色体上的相对位置和转录方向与绵羊所报告的情况基本一致(图1)。
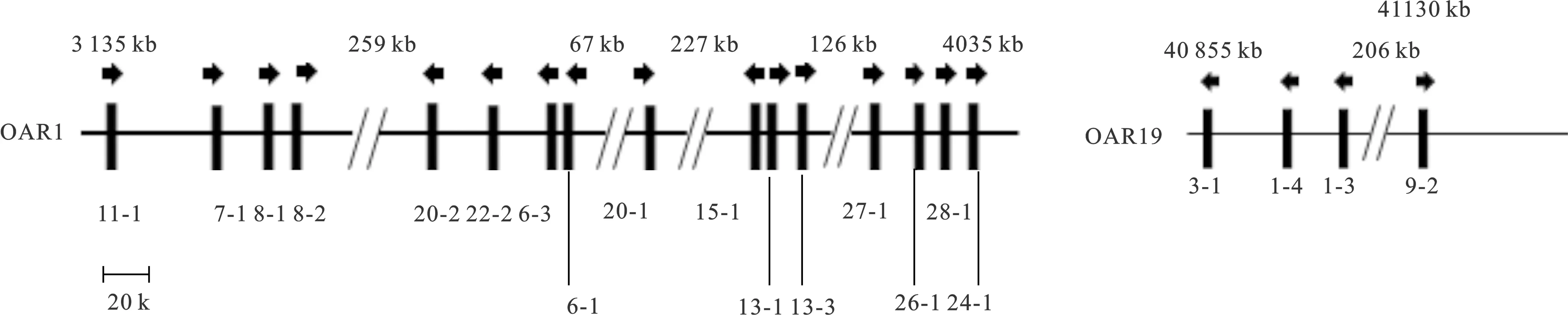
图1 已知山羊KRTAPs在染色体上的位置
3 山羊KRTAPs的核苷酸多态性及其对羊绒性状的影响
3.1 山羊KRTAPs的核苷酸多态性
在KAP1家族中,山羊1-1和1-4多态性均较为丰富。Zhang等和Andrews等在1-1中共检测到6个SNPs和1个30 bp的片段缺失。Shah等在山羊1-4的编码区共观察到7个SNPs,其中5个是同义突变,另外2个是非同义突变,2个非同义突变分别位于编码序列224和357位,分别导致半胱氨酸(C)和丙氨酸(A)转为丝氨酸(S)和苏氨酸(T)。
在KAP6家族中,山羊6-2和6-3多态性已有研究。周群等对随机选取的127只西藏绒山羊的6-2基因进行多态性分析,检测到两种基因型,共发现4处变异,多态信息含量PIC为0.196,属于低度多态位点,说明西藏绒山羊群体6-2遗传性稳定。吴玉江等对西藏绒山羊6-3基因的编码区和部分3'-UTR区(199 bp)进行多态性研究,共发现7个SNPs位点,表现为5种基因型,与刘海英等在内蒙古绒山羊上检测到的SNPs有4处相同。Zhou等和Li等在绵羊6-3中均检测到片段长度多态性,而山羊该基因未检测到类似的突变。
在KAP7家族中,山羊7-1显示出较高的序列保守性。已有研究表明,7-1在辽宁绒山羊、黎城大青羊、辽-岢高代杂种山羊、西藏绒山羊中都没有多态性。彭艺艺等研究发现,在陕北白绒山羊、太行黑山羊和南疆绒山羊中,7-1基因的CDS区也未检测到突变,仅在5'调控区检测到1个SNP。这与绵羊7-1的相关研究结果一致,均没有检测到编码区的突变。
在山羊KAP8家族中,8-1和8-2的多态性均有研究。Zhao等对内蒙古白绒山羊和陕北白绒山羊进行8-1基因多态性检测,发现存在两个同义突变:c.63 T>G和c.66 C>G。Andrews等通过对3个山羊群体(南非安哥拉山羊、波尔山羊和安哥拉×波尔山羊杂交群体)的8-1多态性检测,发现的等位基因与Zhao等前期研究结果相一致。但赵俊星等在辽宁绒山羊、黎城大青羊和辽-岢高代杂种山羊群体中未检测到8-1突变,这种差别有必要进行进一步研究。索朗达等在西藏绒山羊8-2基因编码区序列进行多态性分析,发现8-2基因扩增产物序列中存在5个SNPs位点,分别为C115T、A207G、A214G、G216A和A278T,除了A278T外,其余SNPs位点均位于编码区。
在KAP9家族中,山羊9-2多态性较为丰富。Wang等对3个山羊品种(内蒙古白绒山羊、陕北白绒山羊和关中奶山羊)进行9-2多态性分析,检测到9-2基因在586位存在C/T突变,此突变可能与该基因mRNA翻译或稳定性相关。3个山羊品种该突变位点均处于哈代-温伯格不平衡状态,T和C等位基因的频率在绒山羊和奶山羊中差异显著,并且C等位基因为优势基因。张丹等通过对西藏绒山羊9-2基因CDS区序列克隆测序,检测到25个SNPs 位点及1个30 bp的缺失突变,其中12个SNPs为错义突变,其它13处为同义突变。
在KAP13家族中,山羊13-1和13-3均有研究。吴玉江等对西藏绒山羊13-1基因编码区进行多态性分析,发现5个SNPs位点,分别为C73A、T115C、C124T、C213G 和 G447 A,SNPs位点都位于编码区,其中包括3个非同义突变和2个同义突变。李明娜等在4个山羊群体(河西绒山羊、内蒙古绒山羊、辽宁绒山羊和陇东绒山羊)中进行13-1多态性检测,共发现6种基因型,4个等位基因中存在5个SNPs,分别为G51A、T99C、C108T 、T170G和C197G,且均位于编码区,其中170和197位点的核苷酸突变为非同义突变,分别使氨基酸Leu(CTG)变为Arg(CGG),Thr(ACC)变为Ser(AGC)。Li等首次鉴定山羊13-3基因并分析其多态性,从214只辽宁绒山羊中鉴定出8个等位基因变异,在编码区内发现9个SNPs,均为错义突变。之后,Andrews等对3个山羊群体(南非安哥拉山羊、波尔山羊和安哥拉×波尔山羊杂交群体)进行13-3多态性检测,新发现8个SNPs位点和10个等位基因,说明山羊13-3具有丰富的多态性。
KAP15家族,山羊15-1已有研究。Zhao等在山羊15-1中鉴定出8个SNPs位点形成6个等位基因,其中5个SNPs为非同义突变,引起氨基酸的变化。
山羊KAP20家族中,两个成员20-1和20-2均有研究报道。Wang等以人类20-1序列为模型,在山羊1号染色体上寻找并鉴定了山羊20-1基因,鉴定出4种不同基因型,检测到6个SNPs位点(c.18C>T、c.52G>T、c.105C>T、c.181G>A、c.*11A>G和c.*24C>T),其中c.52G>T和c.181G>A属于非同义突变。经过序列比对发现,山羊20-1与人类20-1序列具有很高的相似性。对山羊20-2多态性检测发现,山羊20-2中存在4个SNPs(c.27C>T、c.37C>T、c.125T>C和c.126G>A),其中3个属于非同义突变,c.37C>T使His变为Tyr,而c.125T>C和c.126G>A可使Met变为Thr。山羊、人和小鼠20-2序列具有较高的同源性。
KAP22家族中,山羊22-2已有研究。山羊22-2存在3个SNPs和1个6 bp的长度突变,其中C.25G>C为非同义突变。有3个等位基因(A、B、C),其中等位基因C存在6个碱基的插入,插入精氨酸和半胱氨酸2个氨基酸。
在山羊KAP24家族中,24-1已有研究。Wang等在山羊24-1中鉴定出4个等位基因,9个SNPs位点,包括8个位于编码区(其中4个非同义突变)和1个接近ATG起始密码子的SNPs。
山羊KAP26家族中,存在1个成员26-1。多态性检测发现,在5个山羊群体(辽宁绒山羊、中卫山羊、柴达木山羊、河西绒山羊和陇东绒山羊)中,山羊26-1存在4个等位基因(A~D),共发现了6个SNPs位点,其中3个属于非同义突变。
KAP27家族,山羊27-1已有研究。Zhao等通过多态性检测发现,山羊27-1存在3个等位基因(A~C)和两个SNPs位点,包括一个同义突变(c.495C>T)和一个非同义突变(c.413C>T;p.Ala138Val)。
山羊KAP28家族的成员,目前有28-1被鉴定研究。Wang等通过多态性检测发现,山羊28-1存在5个等位基因(A~E),存在2或4 bp碱基缺失和11个SNPs位点,其中1个(c.-8A>G)位于5'-UTR,6个(c.*6A>G,c.*39C>T,c.*71C>T,c.*108C>T,c.*112C>T和c.*113G>A)位于3'-UTR,其余变异(c.17G>A,c.129T>A,c.166C>T和c.190A>G)位于编码区内,编码区内的所有SNPs都是非同义突变。
3.2 山羊KRTAPs的核苷酸多态性对羊绒性状的影响
通过分析不同多态位点与产绒性状的相关性,可找到与该性状相关的遗传标记位点,进而为后期分子遗传育种工作提供参考依据。部分KRTAPs家族基因变异与羊绒性状如羊绒细度、产绒量等存在相关性。
目前相关研究表明,山羊28-1、27-1、24-1、11-1、15-1和8-1基因变异与羊绒纤维直径存在相关性,具有作为改良羊绒纤维直径的分子标记价值。6-1、1-3和1-1基因变异与产绒量具有相关性,具有作为产绒量分子标记辅助选择的潜力。高原型藏山羊相关研究表明,6-1和6-2的不同基因型与产绒量、绒细度以及绒长相关,具有分子标记价值。西藏绒山羊相关研究表明,13-1变异与公羊胸围、产绒量和绒长均存在相关性;8-2基因变异与胸深和产绒量有关;6-3基因变异与绒长和产绒量有关,均可作为西藏绒山羊生产性状的辅助选择标记。陇东绒山羊20-1和20-2基因变异与产绒量和绒长存在相关性。
4 山羊KAPs的表达及其对羊绒性状的影响
在人类中,除了KAP16、KAP22、KAP25和KAP27家族外,迄今为止已鉴定的所有基因都在毛囊中表达。羊绒纤维品质的差异与毛发中角蛋白的含量变化有关,含量变化越明显,羊毛纤维品质的差异越大。不同的KAPs可存在于同一细胞中,与不同中间纤维组织状态相联系。不同的KAPs 在毛囊中的表达顺序有所不同。Powell等研究表明,微丝角蛋白(IFs)基因最先表达,其次是HGT-KAPs,HGT-KAPs在不同物种中的卵泡表达差异很大,最后表达的是HS-KAPs。HS-KAPs与角蛋白中的赖氨酸残基形成二硫键,它们之间的疏水作用对于毛干的形成是有重要作用。USH-KAPs富含甘氨酸和半胱氨酸,在毛囊周期性分化过程中表达,且在毛发形成过程中发挥重要作用。人类UHS-KAP中所含有的高水平的半胱氨酸,可能在连接中间微丝角蛋白(KIF)中起到重要作用。
通过qRT-PCR验证,多数山羊基因均只在毛囊中表达,且在次级毛囊中的表达量高于在初级毛囊中的表达量。但存在少数的一些山羊基因,不仅在毛囊中表达,在其它组织中也有表达。如11-1基因在毛囊、心脏、皮肤、肝脏均有表达,而在脾、肺、肾中无表达。在次级毛囊中,11-1在生长期的表达明显高于在其它时期中的表达。此外,11-1基因在内根鞘、毛发基质中表达较强,在毛球中表达较低。
在不同物种之间,基因在毛囊中表达的具体位置和表达量不同。例在人类毛发中,1-4、6 和7基因在毛发的所有皮质层均表达,但在羊毛中6和7基因只在正皮质层表达,4基因只在副皮质层表达。6基因在人类毛发中微弱表达,但在羊毛中高水平表达,山羊6-1在不同时期的初、次级毛囊的皮质层均有强烈的表达信号,且其表达存在品种和性别差异。9-2基因在奶山羊皮肤组织中不表达,而在绒山羊皮肤组织中特异性表达。8-1基因在不同品种绵山羊皮肤组织中的表达量不同,且绒细度越细,该基因相对表达量越高,说明该基因可能对绒山羊绒细度具有正向调控作用。
在同一品种的不同时期,基因的表达同样也存在差异。Jin等发现山羊11-1在初级毛囊中退行期的表达量明显高于生长期,然而在次级毛囊中,11-1在生长期的表达量明显高于在退行期的表达量。赵濛等研究发现,3-1基因在毛囊生长期表达量明显高于退行期。王艳杰等以辽宁绒山羊为试验对象,发现在毛囊生长期,6-2基因在初级毛囊中表达量极显著高于次级毛囊,7-1、8-2和26-1基因在次级毛囊的表达量极显著高于初级毛囊;毛囊退行期,7-1、8-2和26-1基因在初级毛囊的表达水平较高,说明7-1、8-2和26-1基因可能对绒毛细度具有正向调控作用。赵志东等研究发现,在毛囊生长期,高产绒山羊9-2基因的表达量极显著低于低产绒山羊,休止期的表达量极显著高于低产绒山羊,说明该基因可能具有抑制绒毛生长的作用。
通过皮肤毛囊转录组测序(RNA-seq)可筛选出影响毛用性状的差异表达基因,其中包括一些已鉴定的山羊基因。品种不同,绒山羊绒纤维的品质也有明显差异,这可能与不同品种间基因的表达差异有关。刘红亮等通过对常年长绒型与季节长绒型辽宁绒山羊皮肤差异表达基因分析,在常年长绒型绒山羊皮肤组织中共筛选出9个差异表达的基因,其中15-1为上调基因(上调8.76倍),其余均为下调基因。 Ahlawat等通过对印度帕什米纳绒山羊与Barbari肉用型山羊皮肤差异表达基因分析发现,与Barbari羊相比,在帕什米尔绒山羊皮肤组织中筛选出的差异表达基因中,上调的基因有7-1、3-1、11-1(上调5倍),说明基因在不同品种之间的较大表达差异与毛绒生长密切相关。
5 小 结
当前,对于羊毛品质相关基因的研究主要集中在KAPs和Keratin两大类,这两大类蛋白共同决定羊毛的品质。其中毛纤维的90%由Keratin和KAPs组成,两者的性质共同决定羊绒品质,对于羊绒品质相关基因的研究当前也主要集中于这两类基因。
随着绵山羊基因组的功能研究,蛋白分离、测序等生物技术的不断发展,研究者们对羊毛的组分与结构有了更进一步的了解,进而可以对多种主要成分蛋白及编码基因进行更深层次的研究。一些已在绵羊和人类中鉴定的基因,在山羊中有待进一步验证,且可行性较高。研究表明,多数基因对羊毛发育有重要作用,在挖掘绵山羊角蛋白新基因的同时,也应对其功能特性、表达调控和作用机制等进行深入研究。
部分学者认为,由于毛绒生产性状属于数量性状,且数量性状由若干微效多基因协同控制,如果只研究单个基因,可能无法系统的对参与毛绒生长及决定其品质的功能基因网络进行分析,所以有必要借助功能基因组相关研究方法,从全基因组水平揭示有关毛绒生长的分子机制。比如,利用皮肤毛囊转录组测序(RNA-seq)的方法筛选出影响毛绒用性状的差异表达基因,进而为绵山羊毛用性状分子育种提供理论基础。虽然RNA-seq技术已广泛应用于分析动物复杂和重要经济性状的遗传基础,但是该方法需要前期对大量相关基因进行鉴定和验证,只有这样,测序数据才可以反映到特定物种的参考基因组上,如果参考基因组的注释不完整(即序列没有被识别),那么该方法的价值就会受限。与人类和绵羊相比,山羊还有许多基因未被鉴定出来。在所有的角蛋白关联蛋白和角蛋白被鉴定出来之前,RNA-seq技术在研究毛囊活动和羊绒纤维功能基因挖掘方面的应用仍会受限。
猜你喜欢
纺织报告(2022年5期)2022-11-22
中国畜牧杂志(2022年1期)2022-11-06
今日农业(2022年14期)2022-09-15
考试与评价·七年级版(2021年2期)2021-08-14
初中生学习指导·中考版(2020年7期)2020-09-10
新农业(2020年5期)2020-06-01
今日财富(2020年7期)2020-04-07
科技风(2019年15期)2019-10-21
新农村(2018年35期)2018-04-02
中学英语之友·中(2008年12期)2008-12-08
