
基质栽培与土壤栽培对越心草莓蔗糖和柠檬酸积累的影响
2022-07-28 06:47:40杨肖芳李云端孙云帆李绍佳苗立祥张豫超蒋桂华
浙江农业学报 2022年7期
杨肖芳,李云端,孙云帆,李绍佳,苗立祥,张豫超,蒋桂华,*
(1.浙江省农业科学院 园艺研究所,浙江 杭州 310021;2.浙江大学 农业与生物技术学院,浙江 杭州 310058)
栽培草莓(×)因其果实酸甜可口、香味浓郁且营养价值高,在世界各地已成为了重要的经济水果,深受消费者欢迎。果实的酸甜度是草莓品质的重要指标,是影响其经济价值的主要因素之一。果实中可溶性糖类物质主要有3种,即蔗糖、葡萄糖和果糖,不同种类成熟果实中积累的可溶性糖类各不相同,成熟草莓果实中蔗糖含量最高。有机酸是决定果实酸度的重要因子,而影响草莓果实酸度的有机酸主要为柠檬酸。因此,探索生产上调控草莓果实蔗糖和柠檬酸积累的有效手段对于调控其酸甜度至关重要。
参与蔗糖代谢相关的酶主要包括蔗糖磷酸合酶(SPS)、蔗糖合酶(SUS)和转化酶(IVR)。SPS主要与蔗糖合成相关,大量研究表明,SPS与果实蔗糖含量呈正相关,在苹果中过表达基因可显著提高果实的蔗糖含量。SUS既可催化蔗糖分解,也可参与蔗糖合成。研究表明,在梨等果实发育过程中产生了2种不同形式的SUS,即SS1和SS2;SS1在未成熟果实中主要起分解蔗糖的作用,在坐果时分解活性最高,SS2则在成熟果实中起积累蔗糖的作用。IVR则将蔗糖不可逆催化水解成葡萄糖和果糖,IVR主要包括细胞壁转化酶(CWIN)、液泡转化酶(VIN)和细胞质(CIN)3类;IVR与蔗糖含量关系密切,在过表达CWIN抑制剂基因-番茄植株中,CWIN活性明显受到抑制,果实中蔗糖含量则显著上升,达到野生型果实2倍以上。柠檬酸作为果实中常见有机酸之一,广泛存在于各种果实中。线粒体是柠檬酸合成的主要部位。首先,糖在果实细胞内转化为磷酸烯醇式丙酮酸(PEP),形成丙酮酸,脱羧形成乙酰辅酶A(Ac-CoA),CO在磷酸烯醇式丙酮酸羧化酶(PEPC)的催化下与磷酸烯醇式丙酮酸反应生成草酰乙酸(OAA);然后,OAA与Ac-CoA在柠檬酸合酶(CS)的作用下缩合为柠檬酸。已有研究表明,CS为柠檬酸合成的限速酶,过表达基因可促进柠檬酸的合成。在柠檬酸降解过程中,细胞质顺乌头酸酶(ACO)被认为是其降解的关键酶,过表达的果实柠檬酸含量显著降低,而缺失则导致果实柠檬酸含量升高。
目前,尽管蔗糖和柠檬酸的合成通路和分子调控机制已经得到了较为深入的研究,但是生产实践中不同栽培条件对草莓果实蔗糖和柠檬酸合成的影响未有报道。本文以栽培草莓越心(בYuexin’)为研究对象,比较基质栽培和土壤栽培草莓果实蔗糖和柠檬酸积累差异,利用实时荧光定量PCR(qRT-PCR)技术分析蔗糖、柠檬酸代谢相关基因表达量差异,为探究差异形成的分子机制奠定基础,并为实际生产中通过不同栽培条件调控草莓品质提供理论依据。
1 材料与方法
1.1 材料
八倍体栽培草莓越心由浙江省农业科学院海宁杨渡科技创新基地提供。基质栽培的配方为40%泥炭、20%有机介质、10%蛭石、20%植生基盘材、10%珍珠岩、缓释肥料(2 kg·m)、基质杀菌剂、稳定剂,生产厂家均为杭州锦海农业科技有限公司。基质的有机质含量为38.7%、全氮27.6 g·kg、有效磷237 mg·kg、有效钾8 317 mg·kg、电导率(EC)3.27 mS·cm、pH值5.98。土壤的有机质1.62%、全氮1.09 g·kg、有效磷123.23 mg·kg、有效钾194.57 mg·kg、EC 34 mS·cm、pH值7.08。试验周期内用营养液(EC 0.8 mS·cm,pH值5.8)对草莓进行滴灌。
分别在白果期(white,W)、转色期(turning,T)、半红期(intermediate red,IR)和全红期(full red,R)4个发育阶段挑选大小一致且无明显缺陷的果实进行取样。1.5 h内运输到实验室,除去花萼后将果实平分成尖部和基部,迅速切成小块,立即在液氮中冷冻,然后在-80 ℃保存备用。每6个果实为1组生物学重复,每个果实发育阶段取4个生物学重复。
1.2 果实糖酸含量测定
参考Li等的方法进行草莓果实糖酸提取与含量测定。取0.1 g冻于-80 ℃冰箱的草莓果实粉末于1.5 mL离心管中,加入提前放置于-20 ℃预冷的色谱甲醇1.4 mL,涡旋3次使之充分混匀。将离心管置于70 ℃金属浴中950 r·min加热15 min,4 ℃、10 000×离心10 min,将上清液倒入10 mL离心管中,加入750 μL三氯甲烷(-20 ℃)和1.5 mL mili-Q水(4 ℃),涡旋3次,常温下2 200×离心10 min。离心后,吸取上清液于1.5 mL离心管中,置于-40 ℃备用。
吸取100 μL上清液加入20 μL内标核糖醇(20 mg·mL),置于常温下真空干燥。干燥后加入60 μL现配的20 mg·mL甲氧胺盐酸盐(吡啶溶),充分涡旋后37 ℃、950 r·min加热1.5 h,随后加入40 μL N,O-双(三甲基硅)三氟乙酰胺(含1%三甲基氯硅烷),同样条件下金属浴加热30 min。
样品制备完成后,使用气相色谱仪(7890N-5975C,安捷伦公司,美国)测定草莓果实糖酸含量。色谱柱为HP-5MS(30 M×0.25 mm×0.25 μm,J & WScientific,Folsom,CA)。用氮气作为载体,进样口温度250 ℃。进样量为1 μL,分流比10:1,流量为1 mL·min。升温程序:100 ℃保持1 min,3 ℃·min升温至185 ℃,15 ℃·min升至230 ℃,保持1 min。
1.3 果实RNA提取和cDNA合成
参考改良的CTAB法提取草莓果实总RNA。使用PrimeScriptRT(TaKaRa,日本)去除RNA样品中的基因组DNA,随后用1 μg的RNA按试剂盒操作说明(PrimeScript1st Strand cDNA Synthesis Kit,TaKaRa,日本)进行cDNA第一链合成,合成体积20 μL。
1.4 qRT-PCR分析
利用草莓基因组数据库获取蔗糖合酶基因1(gene12940)、2(gene11077),蔗糖磷酸合酶基因1(gene11606)、2(gene22863)、3(gene31122),柠檬酸合酶基因1(gene00583)、2(gene05118)、4(gene26778),顺乌头酸酶基因1(gene14541)、3(gene23775)的CDS序列,利用PrimerPremier5.0设计引物。对cDNA以1∶10的比例进行稀释,2 μL作为模板进行qRT-PCR。反应总体积20 μL,包含10 μL Ssofast EvaGreen Supermix(Bio-Rad,美国),上下游引物(10 μmol·L)各1 μL,6 μL DEPC水和2 μL稀释后的cDNA模板。qRT-PCR在CFX96仪器(Bio-Rad)进行。qRT-PCR扩增程序:95 ℃ 3 min;95 ℃ 10 s,60 ℃ 30 s,45个循环。
以413为内参基因进行表达量的校正,并将土壤栽培和基质栽培中全红期基因的表达量设为1,用2法计算果实成熟过程中各基因的相对表达量。
1.5 数据处理
采用SPSS软件进行相关性分析和统计分析,使用Graphpad Prism 8软件作图。

表1 实时荧光定量PCR引物序列
2 结果与分析
2.1 不同栽培条件下草莓果实的蔗糖含量
草莓果实尖部的成熟早于基部,蔗糖的积累是果实成熟的重要性状之一。因此,把果实分为尖部和基部两部分进行分析。蔗糖含量测定的结果显示,蔗糖含量随草莓果实的发育不断增加,在果实完全成熟时其蔗糖含量达到最大值。土壤栽培的果实尖部蔗糖积累在白果期、转色期、半红期和全红期均显著高于基质栽培。在果实基部,除白果期土壤栽培比基质栽培高外,其他3个时期2种栽培条件下蔗糖含量均无显著差异(图1)。
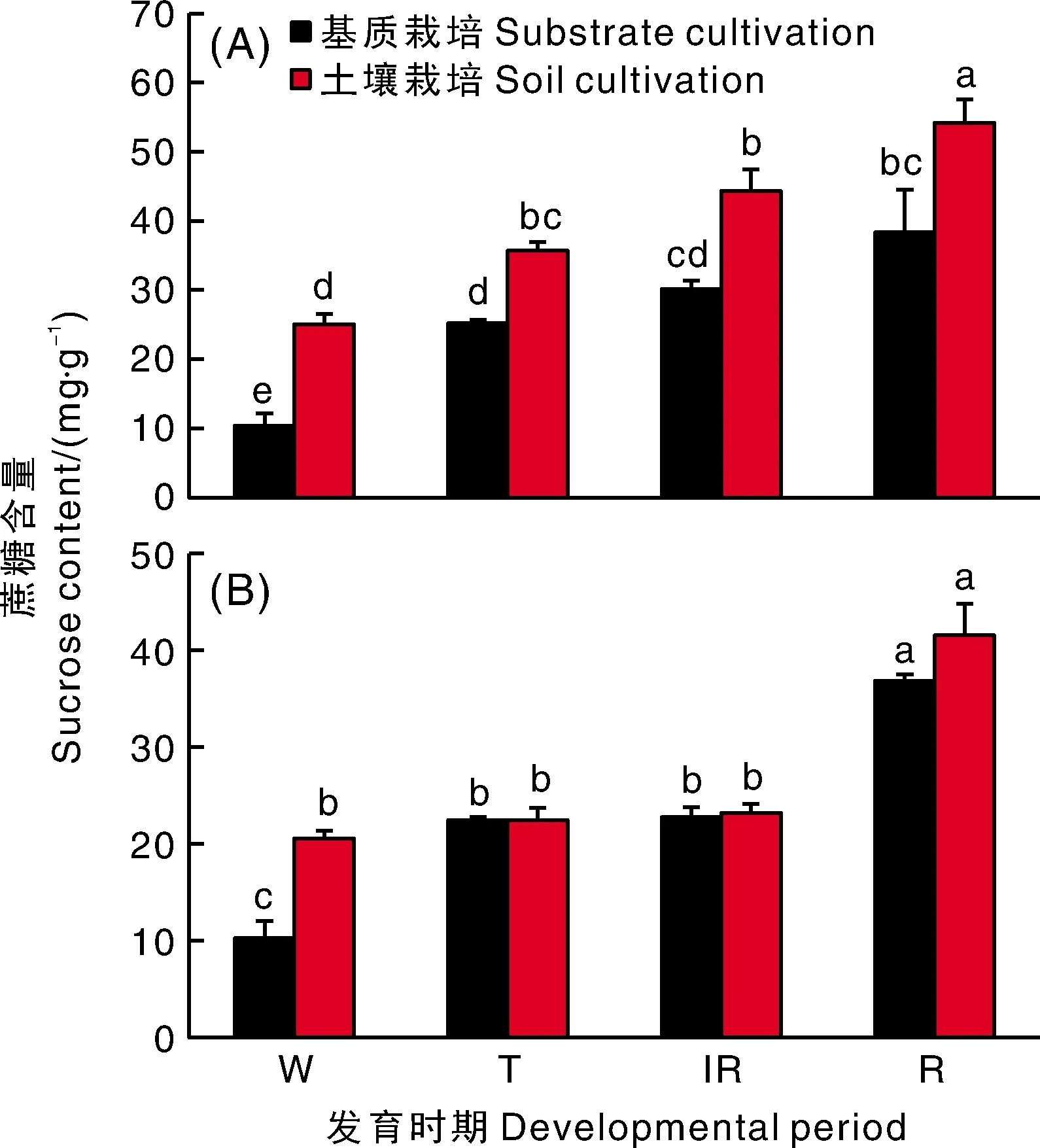
A,果实尖部;B,果实基部。W,白果期;T,转色期;IR,半红期;R,全红期。下同。
2.2 不同栽培条件下草莓果实的柠檬酸含量
在草莓果实发育过程中,基质栽培果实尖部和基部的柠檬酸含量先上升后下降,而土壤栽培的则随果实的发育逐步下降(图2)。土壤栽培果实尖部和基部的柠檬酸含量均显著高于同一时期基质栽培的果实,表明土壤栽培可能较基质栽培更有利于柠檬酸的积累。
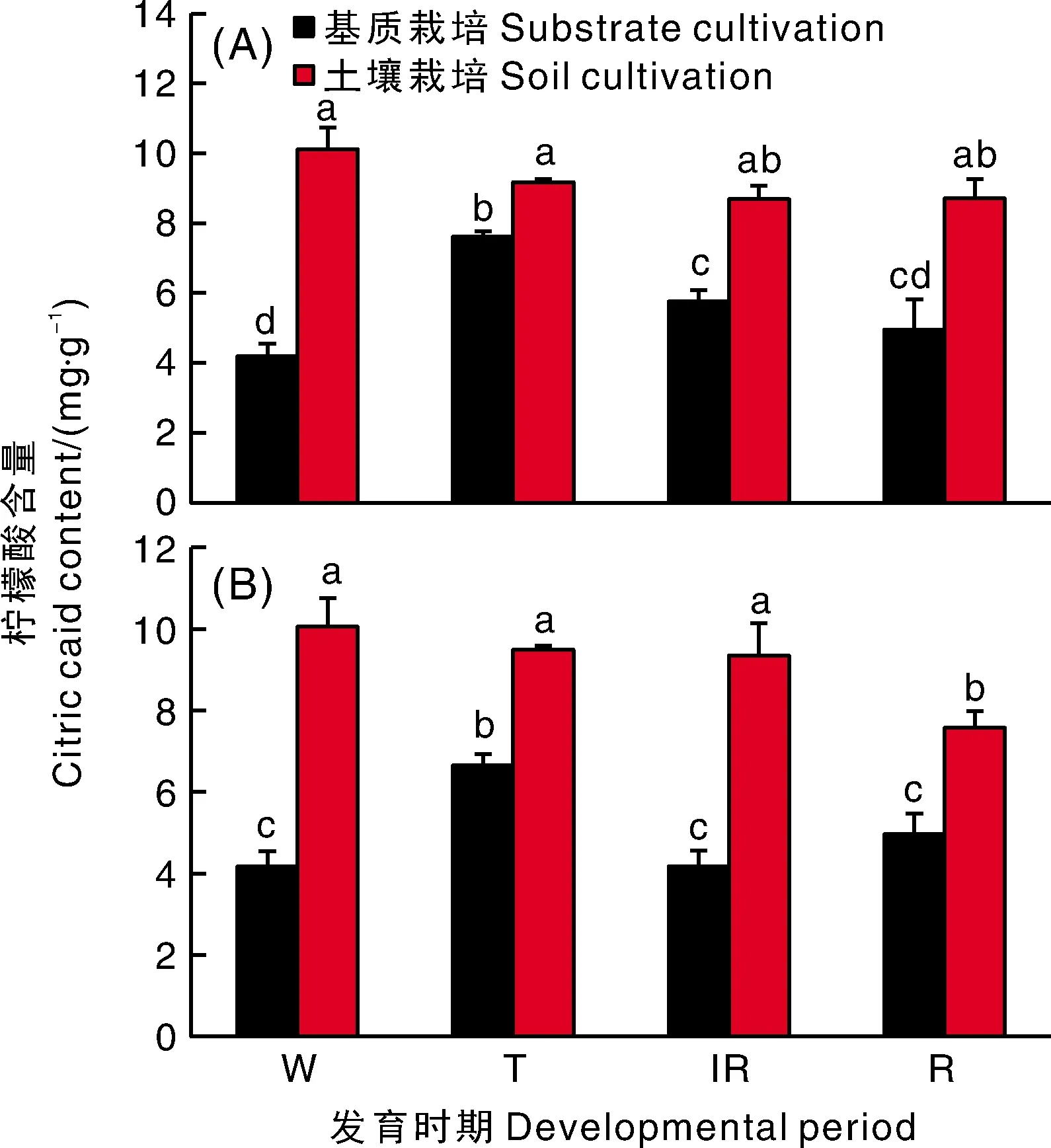
A,尖部;B,基部。
2.3 不同栽培条件下草莓果实蔗糖合成相关基因的表达模式
从图3可以看出,基质栽培条件下,果实尖部1基因在全红期的表达量显著高于土壤栽培。果实基部,土壤栽培条件下2基因在半红期和全红期的表达量显著高于基质栽培。果实尖部和基部,基质栽培和土壤栽培条件下3基因在相同发育时期没有显著性差异。在果实尖部,土壤栽培条件下1基因在白果期的表达量显著高于基质栽培,而在全红期土壤栽培条件下1基因的表达量显著低于基质栽培。在果实基部和尖部,土壤栽培条件下果实中2基因的表达量在白果期均显著高于基质栽培,其他时期均无显著性差异。进一步分析表明,只有2的表达量在两种栽培条件的果实中随发育呈上升趋势,且与蔗糖的积累呈正相关(相关系数=0.567)(图4)。这与土壤栽培条件下果实蔗糖含量高于基质栽培条件下的果实相一致(图1)。
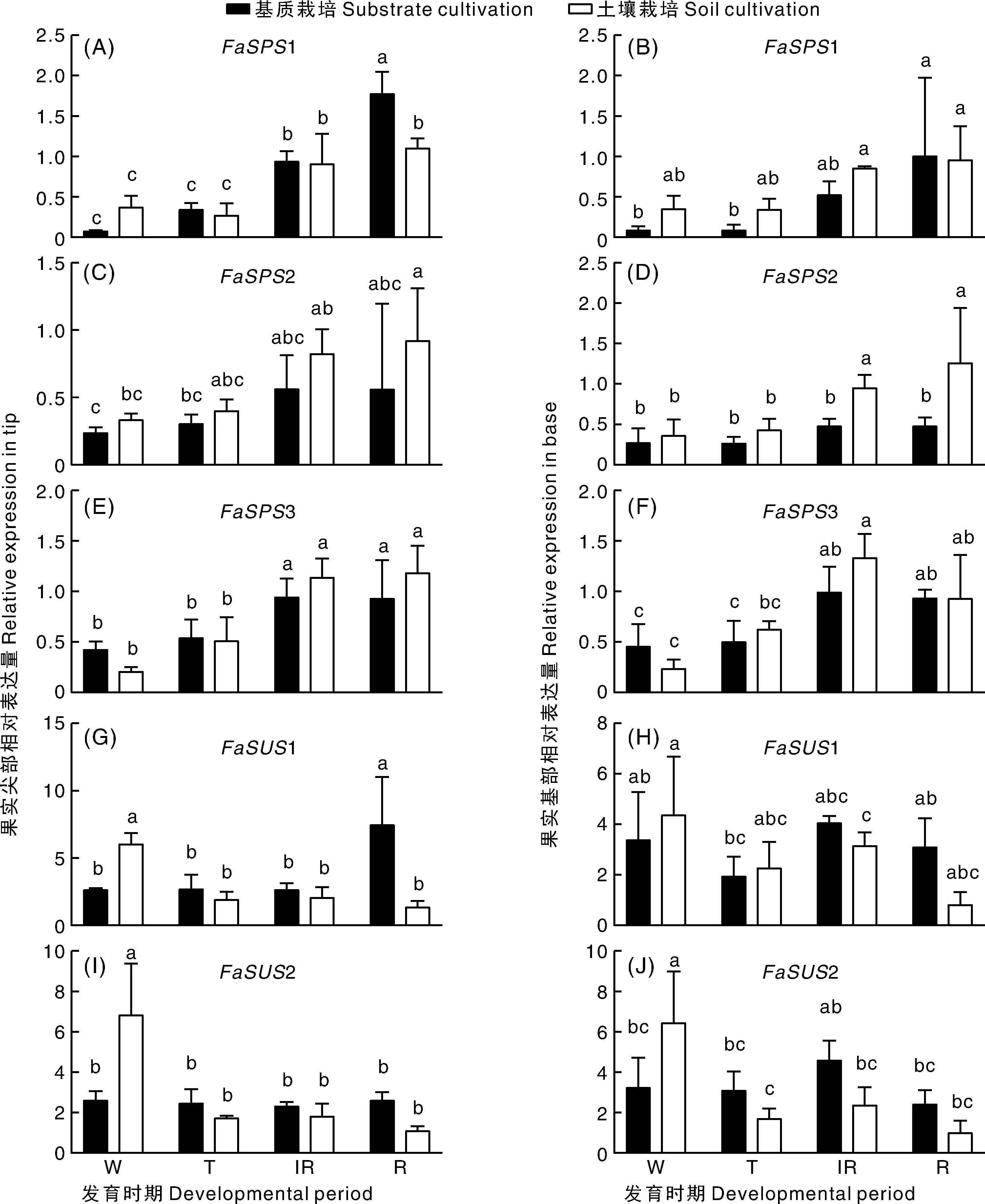
图3 基质栽培和土壤栽培条件下蔗糖合成代谢相关基因的表达模式
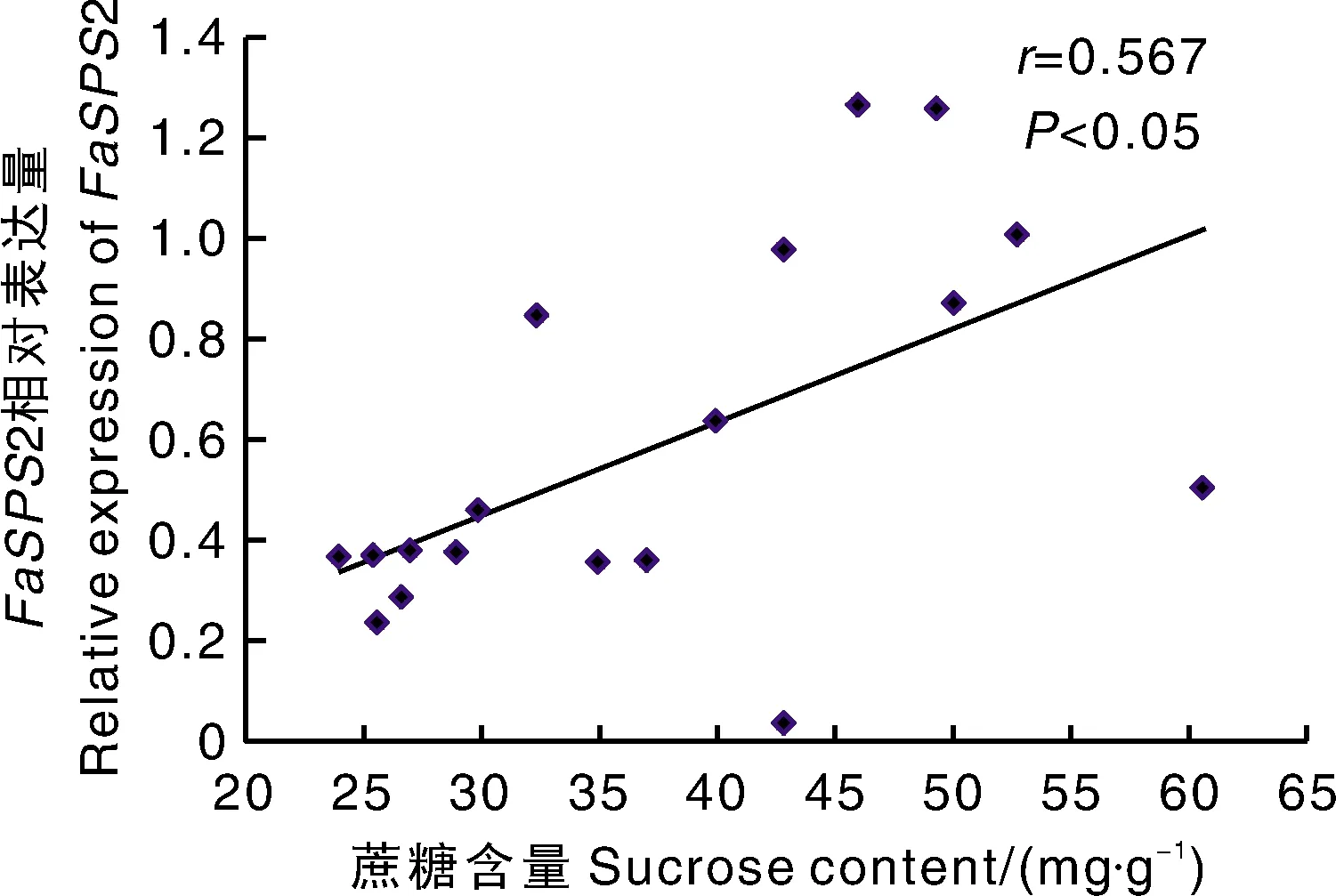
图4 蔗糖含量与FaSPS2基因相对表达量的相关性分析
2.4 不同栽培条件下柠檬酸降解相关基因的表达模式
从图5可以看出,随着果实发育进程的推进,与柠檬酸代谢相关的基因表达量有不同程度的差异,其中,1和3基因的表达量在土壤和基质栽培条件下有显著差异。在半红期,果实基部基质栽培的1基因比土壤栽培的表达量低,其他时期基质栽培条件下1、3基因的表达量都高于土壤栽培。相关性分析结果表明,1、3基因的表达量与柠檬酸的积累呈负相关(图6),这与基质栽培条件下果实柠檬酸含量低于土壤栽培条件下的果实相符合(图2)。基质栽培和土壤栽培条件下果实基部1基因表达量仅在半红期有显著差异,其他均无显著差异。土壤栽培栽培条件下果实尖部2基因表达量仅在全红期显著高于基质栽培,果实基部2基因表达量仅在半红期显著高于基质栽培。土壤栽培条件下果实基部4基因的表达量仅在半红期显著高于基质栽培。
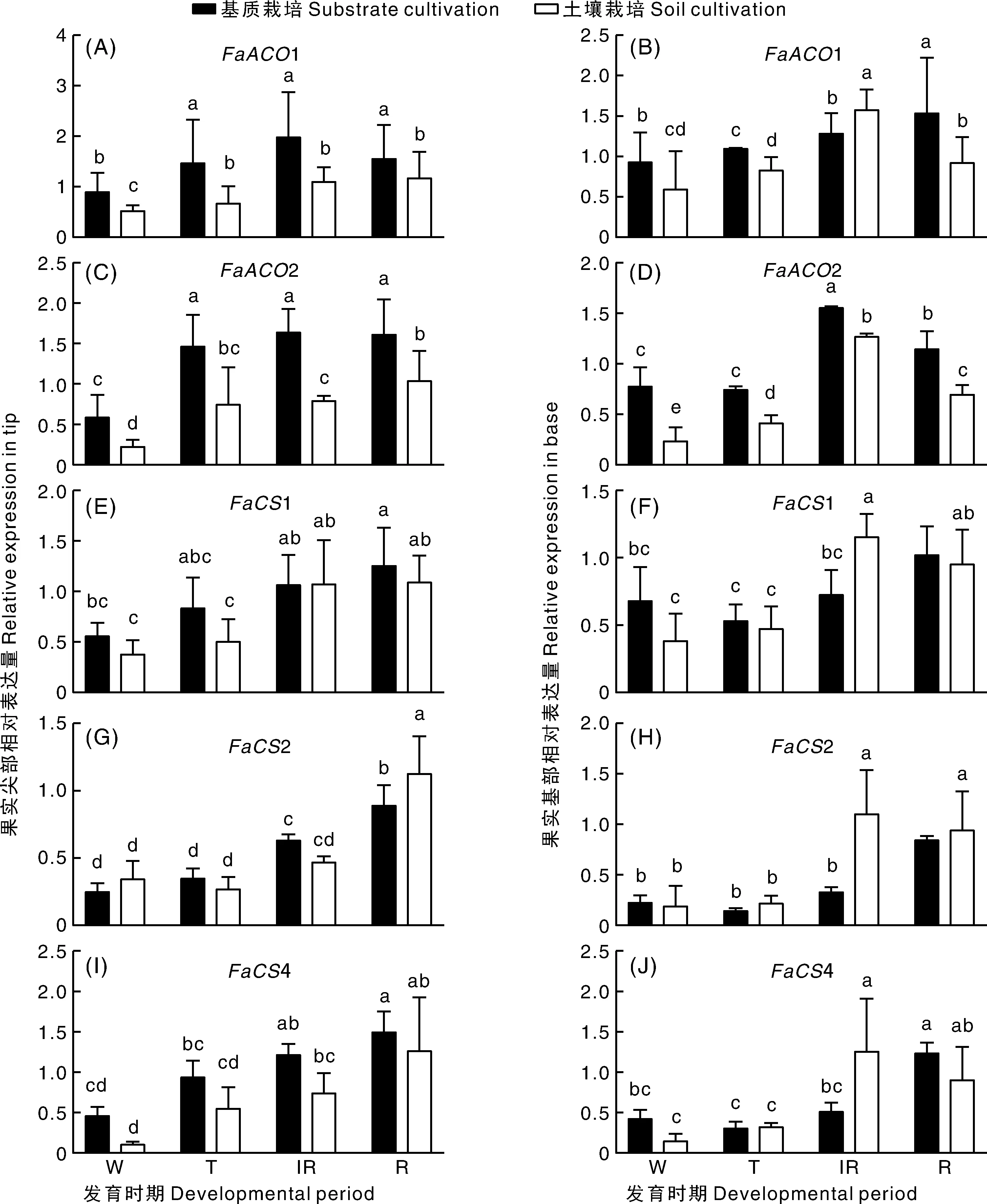
图5 基质栽培和土壤栽培条件下柠檬酸降解相关基因的表达模式
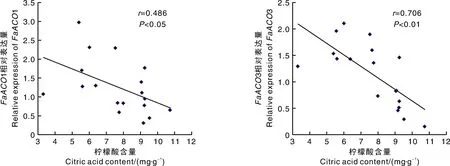
图6 柠檬酸含量与FaACO1基因和FaACO3基因相对表达量的相关性分析
3 结论与讨论
本研究表明,参与蔗糖合成的关键基因2的表达量与蔗糖的积累呈正相关,与柠檬酸降解相关的关键基因1、3表达量与柠檬酸的积累呈负相关,土壤栽培较基质栽培更利于越心草莓积累蔗糖和柠檬酸。
栽培环境直接影响植株生长状况、产量和品质。已有研究表明,土壤栽培比基质栽培更有利于促进草莓植株的生长,果实的品质、口感风味也较好。分别用土壤和基质进行罗马花椰菜栽培时,与基质栽培相比,土壤栽培不仅可以提高单果重和产量,还能提高可溶性糖含量。本实验也得出了类似的结论,土壤栽培的越心草莓可以比基质栽培积累更多的蔗糖和柠檬酸。也有研究表明,基质栽培比土壤栽培在品质改善方面更有效果。当用海岛土壤进行草莓栽培时,其糖度会低于基质栽培。基质栽培的番茄能显著提高净光合速率,有利于培育壮苗,提高可溶性糖含量,改善糖酸比。导致这些研究结果不一致的原因,除了作物种类不同外,还有可能与品种特性、栽培环境的理化特性,以及所含的有效营养元素含量有关。
草莓果实中蔗糖分布具有一定的梯度,果实尖部的含量大于基部含量。本研究发现,不同栽培环境不会改变这一特性。草莓土壤栽培较基质栽培显著提高了蔗糖含量,主要是提高了尖部的蔗糖含量,但两种栽培方式下果实基部蔗糖含量差异不显著。1、2、1、2、3等基因在草莓果实蔗糖合成与积累中扮演了重要的角色,然而本研究显示,这些基因的表达量并没有一定的规律。进一步将基因表达量与糖含量进行相关分析发现,只有2的表达量在两种栽培条件的果实中随发育呈上升趋势,且与蔗糖的积累呈正相关。
柠檬酸是草莓果实中最重要的有机酸,与可溶性糖一起决定了果实的主要口感。无论是土壤栽培还是基质栽培,果实基部和尖部的柠檬酸含量相差不大。白果后期,随着果实的发育柠檬酸逐渐被降解,起作用的基因有1、2、4、1、3等。本研究显示,白果期草莓的柠檬酸含量最高,随着果实不断成熟,柠檬酸含量逐渐降低。在全红期,土壤栽培的果实柠檬酸含量显著高于基质栽培。由于1、3基因在果实成熟过程中主要负责柠檬酸的降解,且与柠檬酸的积累呈负相关。因此推测,1、3的表达量差异可能是引起土壤栽培与基质栽培草莓果实柠檬酸含量差异的主要原因。
猜你喜欢
江苏调味副食品(2021年1期)2021-04-01 12:51:12
广西糖业(2020年3期)2020-09-25 08:32:44
浙江工业大学学报(2017年5期)2018-01-22 02:03:33
热带农业科学(2017年9期)2017-10-23 22:56:24
中国糖料(2016年1期)2016-12-01 06:49:06
Acta Mathematica Scientia(English Series)(2016年5期)2016-11-24 11:59:31
农村农业农民·B版(2016年7期)2016-10-21 10:44:23
西南农业学报(2016年4期)2016-05-17 05:41:45
应用化工(2015年2期)2015-07-13 03:12:26
中国果业信息(2015年11期)2015-01-23 19:19:58
