雅长保护区老龄林树种空间优势度二阶特征分析*
2022-07-20 08:34黄绍娴王宏翔王耀仪李远发叶绍明
林业科学 2022年4期
黄绍娴 王宏翔 彭 辉 王耀仪 李远发 叶绍明
(1.广西大学林学院 南宁 530001; 2.凤阳山-百山祖国家级自然保护区凤阳山管理处 龙泉 323700)
近年来,生态系统退化和物种多样性丧失,使得全球生物多样性面临巨大威胁和挑战,维持和保护生物多样性成为国内外研究热点之一(马克平等, 1998; Pimmetal., 2014; 李爱农等, 2018)。结构多样性是森林生态系统多样性的重要内容,其综合反映了森林的发展过程,如起源和更新、林木生长、死亡以及外部干扰活动等(雷相东等, 2002; Gadowetal., 2012; 赵中华等, 2020),同时也决定了森林未来发展方向及其所发挥的功能(如生产力)(Zhangetal., 2017),通常采用物种多样性、林木位置分布格局多样性和林木大小多样性等来刻画和描述(Leietal., 2009; 惠刚盈, 2013; Lietal., 2020)。以往关于森林群落生态学的研究大多侧重于对树种多样性及其形成机制的探讨(Loreauetal., 2001; Cardinaleetal., 2006; Gamfeldtetal., 2011; Isbelletal., 2011),忽略了林木大小空间异质性特征及其对树种多样性维持的作用。在森林生态系统中,由于光照和养分等生存资源的局限性,林木大小特征是其个体及种群适应生长环境的结果,不同大小林木在空间中组织在一起形成层次复杂的群落结构,也被称为大小分化或大小多样性 (Weineretal., 1984; 1986; Wangetal., 2020)。研究发现,林木大小多样性对维持鸟类、昆虫以及土壤微生物等多样性具有重要意义(雷相东等, 2002),然而与其重要性相反,大小多样性在群落生态学研究中很少被关注。
同一群落中,林木个体之间大小存在差异,基于特定树种计算的林木个体大小优势度很大程度上可反映该树种在群落中的竞争态势(惠刚盈等, 1999; 赵中华等, 2014)。关于树种优势度表达方法,现主要有树种重要值法(Curtisetal., 1951; 邵方丽等, 2012; 蔡军奇等, 2019)和基于最近相邻木关系构建的指数(惠刚盈, 2013; 惠刚盈等, 2019),因这些方法具有数据获取相对容易、计算过程相对简单、结果容易解释等优势,常被应用于森林经营实践中(Szmyt, 2014; Lietal., 2014; 2020); 但也由于这些方法未考虑林木大小优势度的空间尺度效应,在详细分析树种空间优势度及其动态变化等方面存在不足。在空间点过程统计中,标记二阶特征方法弥补了这一缺陷,其可分析不同空间尺度上林木位置和属性的分布特征,并能够检验林木及其属性在空间中的相关关系(Getzinetal., 2006; Wangetal., 2017; 2018),因此,很有必要将树种优势度分析方法与标记二阶特征方法相结合,以此来刻画不同空间尺度上的树种优势度,为保护关键树种及监测其竞争态势和空间优势度动态变化提供有效途径。
此外,群落内树种空间优势度与树种混交和树种空间分布格局可能存在一定联系,但该联系如何解释物种共存鲜见报道。生态学家为了解释众多物种如何实现共存提出了许多理论和假说,如生态位和中性理论(牛克昌等, 2009)、种库理论(Zobel,1997)、Janzen-Connell假说(Janzen,1970; Connell,1971)、空间隔离假说(Pacalaetal., 1996; Pacala, 2009; Chesson, 2000a)等,其中,空间隔离假说被认为是解释物种共存的重要机制之一(Wangetal., 2010)。该假说认为,因种子扩散限制和生境异质性的影响,林木主要聚集在同种个体周围,从而降低了种间竞争相较于种内竞争的重要性(Stoll, 2001; Wangetal., 2010),这种效应可改变林木种内种间相遇的概率,降低树种混交程度,使个体更倾向于干扰同种植物(Pacalaetal., 1996; Dettoetal., 2016)。根据树种空间隔离假说,本研究认为树种空间优势度主要受同种林木个体影响,处于竞争优势的物种种群往往会受到抑制,降低其空间优势度,同时又会阻止或减缓其对群落中其他劣势树种的竞争排除作用,从而使物种打破竞争转向共存,促进物种多样性(Stoll, 2001; Fibichetal., 2011; 王进等, 2020)。因此,分析树种空间优势度与树种空间分布格局和树种混交之间的关系可为探究树种共存提供理论依据,对保护和维持森林群落树种多样性具有指导性意义。
处于演替后期的森林群落与其他类型林分相比,物种多样性更丰富、结构更复杂、生态系统更稳定(马姜明等, 2007),分析其树种空间优势度可为探究森林结构多样性的形成提供重要参考。鉴于此,为了更加详细刻画和表达树种空间优势度,反映树种空间隔离对物种共存的贡献,本研究以雅长自然保护区常绿阔叶林为研究对象,将树种优势度分析方法与标记二阶特征方法相结合,旨在检验基于大小比数和双曲正切指数构造的标记二阶特征函数的有效性,探究随着空间尺度范围增加不同树种空间优势度的变化特征,分析树种空间分布格局和树种混交对树种空间优势度的影响,以期为保护关键树种及监测其空间优势度动态变化提供有效途径,为维持森林群落物种多样性提供理论依据。
1 研究区概况
研究区位于广西壮族自治区百色市乐业县雅长兰科植物国家级自然保护区(106°11′31″—106°27′04″E,24°44′16″—24°53′58″N),区内有北盘江、南盘江和红水河等河流,所在江河段属龙滩水电站上游库区,地理位置独特,生态区位重要性突出。属南亚热带季风气候,热量丰富,干湿季节明显,年均气温16.3 ℃,冬季有霜雪,区内因焚风效应影响降水量较少,年降雨量和年蒸发量分别为1 051.7和1 484.7 mm,年均相对湿度82%(李述万, 2017)。海拔500 m以下的红水河河谷地带主要分布褐红土,500~1 000 m主要分布山地红壤,1 000 m以上发育成山地黄壤,部分山体顶部则发育成草甸土; 受成土母岩影响,在石灰岩地区发育有黑色石灰土和棕色石灰土(和太平等, 2007)。保护区植被类型属西部南亚热带常绿阔叶林,野生动植物资源种类繁多,生物多样性丰富。
试验样地位于广西国有雅长林场九龙分场,平均海拔1 750 m。样地保存完好,无明显人为和牲畜干扰,无伐桩,植物种类十分丰富,其中分布着大量胸径较大、树高较高的植株,如枫香树(Liquidambarformosana)、银木荷(Schimaargentea)、贵州桤叶树(Clethrakaipoensis)、红楠(Machilusthunbergii)等; 同时,样地内还有较多数目的自然枯死木,属典型老龄林群落。
2 研究方法
2.1 样地设置
2.1.1 实测样地 2020年7月,在查阅雅长自然保护区森林植被分布概况的基础上,设置1块面积1.6 hm2(200 m × 80 m)的固定样地。使用全站仪(NTS-372R10)将样地划分为40个20 m × 20 m小样方,在每个小样方4个顶点处埋设PVC管,并用绳子将格子边界框起来作为标记。用全站仪对样地内每木进行定位,获取位置坐标(x,y,z); 同时,调查样地内所有胸径(DBH)≥1 cm的木本植物,对每株植株挂上带有编号的不锈钢牌并进行物种鉴定,记录其胸径、树高、枝下高等。

具体算法如下: 首先,随机选择模拟样地中具有不同物种属性的一对点,并对二者的树种和大小属性进行临时交换; 接着,根据能量函数C(式1)确定属性交换后的树种空间配置结构是否接近于期望状态,若Cnew
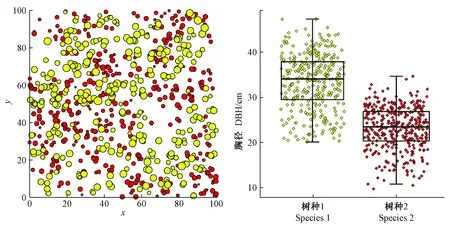
图1 模拟林分树种空间分布(左)和树种胸径分布(右)Fig. 1 Spatial distribution of trees of two species in simulated stand (left) and DBH distributions of species (right)
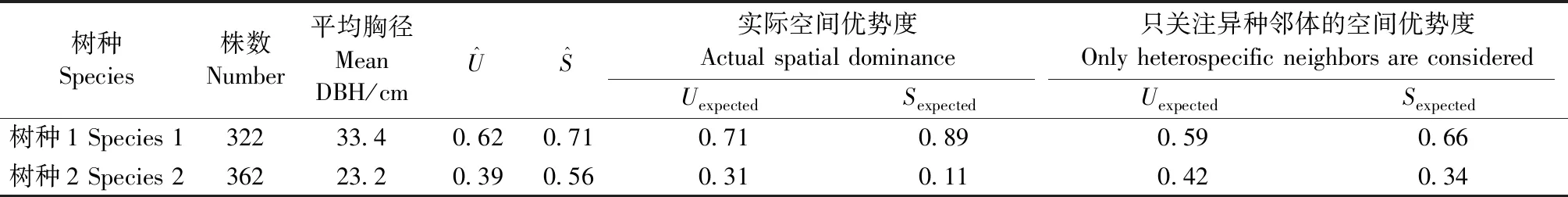
表1 模拟样地概况Tab.1 Characteristics of the simulated plot
在模拟退火算法中,能量函数选择对树种空间结构配置非常重要,其决定着模拟过程。能量函数通常选择最小二乘误差函数(Tschescheletal., 2006),其表达式为:
(1)

本算法基于物种分离指数(Pommereningetal., 2019)模拟种群聚集分布模式,其表达式为:
(2)
式中:M为混交度,即与参照树不属同种的相邻木个数占全部最近相邻木的比例;Mexpected为与相邻木位置和个数无关的期望混交度。
综上,产生的模拟林分中,树种1为较大径级树种,处于较优势地位,树种2为较小径级树种,处于较劣势地位,且树种之间存在空间隔离。本研究通过模拟数据检验标记大小比数和标记双曲正切指数二阶特征函数对空间优势度的表达能力以及树种空间分布格局和树种混交对空间优势度的影响。
2.2 研究方法
2.2.1 林木空间优势度指数 惠刚盈等(1999)提出描述林木个体大小优势度的大小比数(Ui,dominance),在林分空间结构分析中,基于特定树种计算的大小比数是表达该树种生长优势度的重要指标。大小比数定义为小于参照树的相邻木个数占全部最近相邻木的比例,其表达式为:
(3)
式中:mi和mj为植物i和j的大小变量,0≤Ui≤1。Ui=0,表示n株邻近木均大于参照木i,参照木处于劣势状态;Ui=1,表示n株相邻木均小于或等于参照木i,参照木处于绝对优势。
树种期望大小比数(树种林木大小在空间中随机分布时树种的大小比数)与选取邻体数量无关,其表达式为:
(4)
式中:N代表所有林木个体数;n代表关注树种林木个体数; DOi代表将所有林木个体大小变量按升序排列后(m1≤m2≤ … ≤mN)比mi小的个体数量。当只考虑异种邻体计算大小比数时,期望值公式中(式4)N代表与关注树种异种的林木个体数; DOi代表与关注树种异种林木个体大小变量按升序排序后(m1≤m2≤ … ≤mN)比mi小的个体数量,此时分母为N×n。
另外,Pommerening等(2020a)提出表达林木空间大小分化程度的最近相邻木指数——双曲正切指数 (Si, hyperbolic tangent index)。与其他空间结构多样性指数相似,双曲正切指数及其概率密度函数可对树种种群的邻体大小分化程度进行估计,其表达式为:
(5)
式中:mi和mj为植物i和j的大小变量;k为相邻木个数;α为表示邻体间相互作用对称 (=0)或不对称 (→∞)的参数,一般取值为1。Si取值范围[0,1],值越大,植物i越占优势。
双曲正切指数的期望值表达式为:
(6)
式中:N代表所有林木个体数;n代表关注树种林木个体数;α=1。若只考虑异种邻体计算双曲正切指数时,期望值公式中(式6)N代表与关注树种异种的林木个体数,此时分母为N×n。
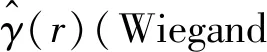
1(‖xi-xj‖,r)×ωi,j。
(7)

(8)
2.2.3 零模型 在点过程统计中,通过分析实际结构特征与零模型模拟期望值之间的关系,可检验林木大小在空间中的分布特征是否偏离零模型的随机过程及其偏离程度。由于不同树种可能具有特定的大小(如胸径)范围(Wangetal., 2020; Pommereningetal., 2020b),因此零模型需要保持林木大小和树种属性不变,为移除林木大小属性的空间相关性,将所有个体位置完全随机。通过999次Monte Carlo模拟得到95%置信区间,将实测值与模拟值相比,分析树种空间优势度。当δ(r)在上包迹线之上,则该树种比期望值更占优势,具有吸引小树、排斥大树的趋势; 当δ(r)在下包迹线之下,则该树种较期望值更为劣势,具有吸引大树、排斥小树的趋势(图2)。
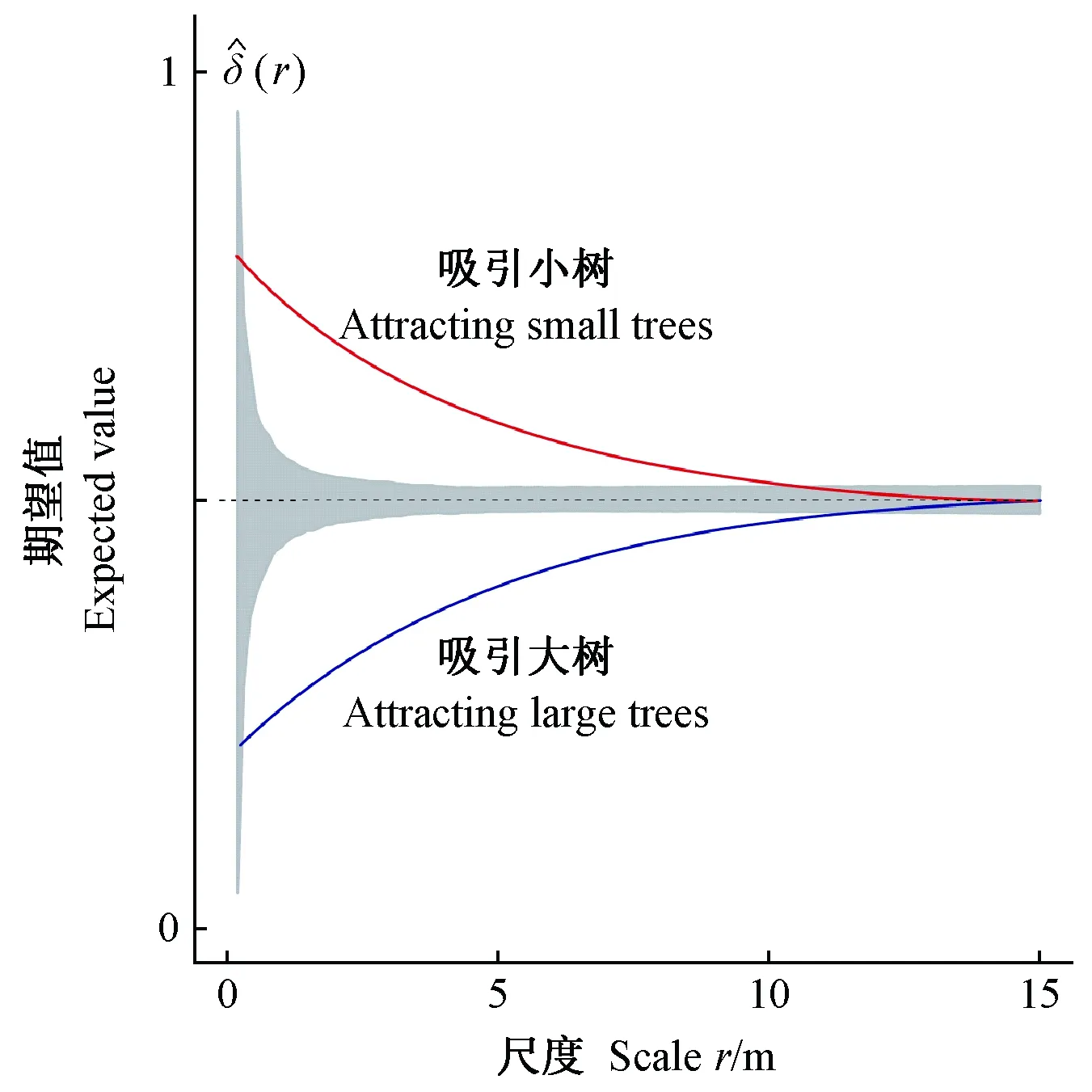
图2 标记二阶特征函数示意Fig. 2 Schematic diagram of mark second order functionsδ(r)
2.3 数据分析与处理
为探究树种混交和树种空间分布格局对空间优势度的影响,本研究分别计算主要树种的实际空间优势度(关注所有的同种和异种邻体)以及只关注异种邻体的空间优势度。
以上所有计算分析均使用 R 统计软件进行。标记大小比数和双曲正切函数的计算以及模拟林分数据通过编写的R和C ++代码实现; R函数包Rcpp(Eddelbuettel, 2011; Eddelbuetteletal., 2013)、spatstat(Baddeleyetal., 2016)用于计算中。
3 结果与分析
3.1 样地内种群结构特征
研究样地内记录木本植物86种,共10 082株,隶属于35科61属,主要树种组成见表2。样地内所有树种的总胸高断面积为49.81 m2·hm-2,其中胸高断面积大于1.0 m2·hm-2的物种有7个,分别为多花杜鹃 (Rhododendroncavaleriei)、枫香树、凹脉柃(Euryaimpressinervis)、银木荷、贵州桤叶树、红楠、榕叶冬青(Ilexficoidea),前5个树种的重要值也较高,为该林分的主要优势树种; 细齿柃和榕叶冬青的平均胸径虽然很小,但其株数分别排名第一和第二,在林分中占有一定地位(图3左)。因此,本研究将样地内重要值排名前5位的主要优势树种(多花杜鹃、枫香树、凹脉柃、银木荷、贵州桤叶树)和典型小径级树种(细齿柃、榕叶冬青)作为主要关注树种。
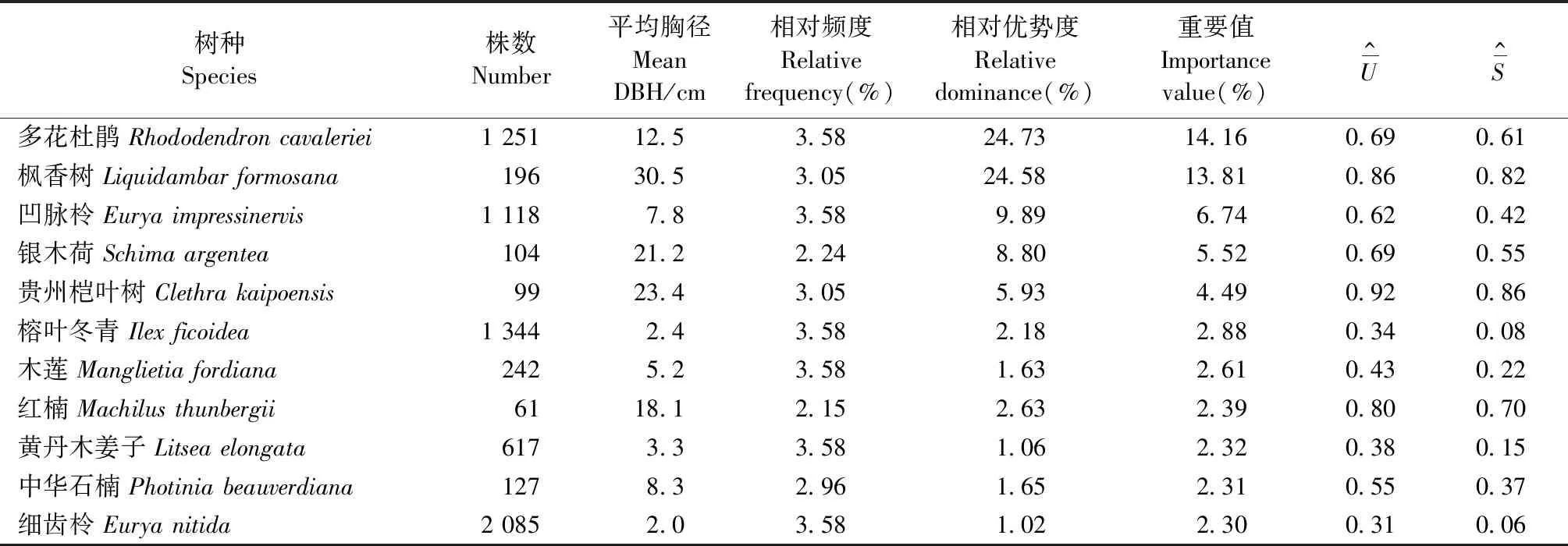
表2 实测样地内种群组成的数量特征Tab.2 Quantitative characteristics of species composition in the experimental plot
主要树种的径级结构见图4,多花杜鹃的径级结构为偏常态分布,以DBH = 15 cm为中心向两侧递减; 枫香树在DBH = 35 cm时出现高峰,其最大胸径达117 cm,为样地内较大径级树种; 凹脉柃表现为倒J形分布,但在DBH >15 cm时株数显著下降; 银木荷种群的小径级和中径级木个体数较多; 贵州桤叶树不存在DBH < 8 cm的林木个体,胸径集中分布于较大径级; 细齿柃和榕叶冬青胸径分布均呈倒J形,具有充足、大量的幼树,2个树种胸径集中分布于1~8 cm的小树径级。因此,群落中不同树种具有不同胸径分布范围(图3左、图4)和优势度(图3右)。同时,基于4株相邻木计算的主要树种大小比数和双曲正切指数(表2)与树种平均胸径之间存在显著正相关(R2= 0.79、R2= 0.81)(图3右)。
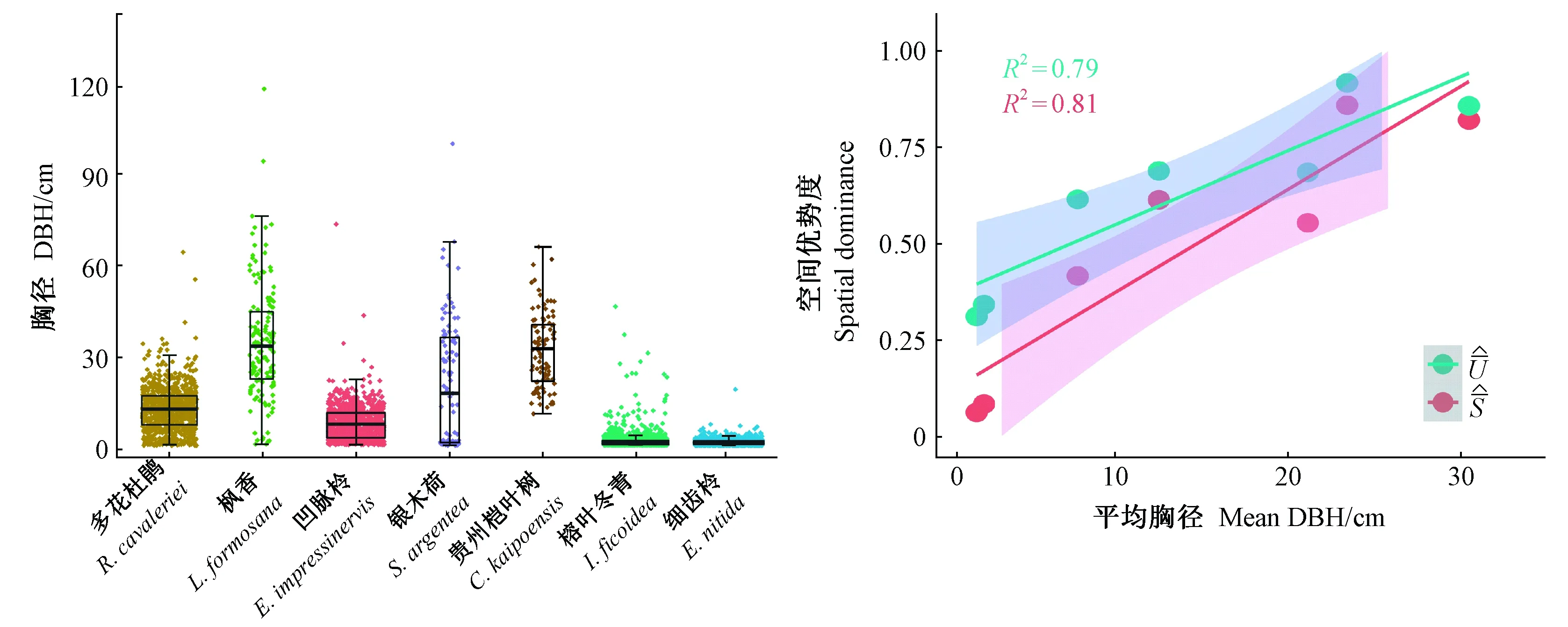
图3 主要树种DBH箱线图(左)及其平均胸径与大小比数、双曲正切指数回归(右)Fig. 3 Boxplot of DBH for the main species (left) and relationships between tree species mean DBH and dominance/hyperbolic tangent index values (right)箱体宽度表示样本量The width of the box indicates sample sizes of each species.
3.2 树种空间优势度分析
模拟样地中包括2个树种,树种1为大径级树种,基于4株相邻木计算的大小比数和双曲正切指数分别为0.62、0.71,在模拟林分中较占优势; 树种2径级较小,其大小比数和双曲正切指数分别为0.39和0.56,在模拟林分中处于较劣势地位(表1)。同时,对模拟数据的分析表明,标记大小比数和双曲正切指数二阶特征函数能够有效表达树种空间优势度(图5)。树种1期望大小比数和双曲正切指数分别为0.71、0.59(表1),但在小尺度范围内实际空间优势度(同时考虑同种和异种邻体)却低于随机区间(图5Aa、Ca),表明树种1周围分布着较大邻体,导致其局部空间优势度降低; 若只关注异种邻体,期望大小比数和双曲正切指数分别为0.89、0.66 (表1),此时,其空间优势度处于随机区间内,且较实际空间优势度高 (图5Ba、Da),说明种内聚集效应降低了树种1的空间优势度。树种2的期望大小比数和双曲正切指数分别为0.31、0.42(表1),但在小尺度范围内实际空间优势度却高于期望值(图5Ab、Cb),表明树种2周围分布邻体较小,提高了其邻域范围的空间优势度; 若只关注异种邻体,期望大小比数和双曲正切指数分别为0.11、0.34(表1),空间优势度位于随机区间内,且较实际空间优势度低(图5Bb、Db),说明种内聚集效应提高了树种2的空间优势度。
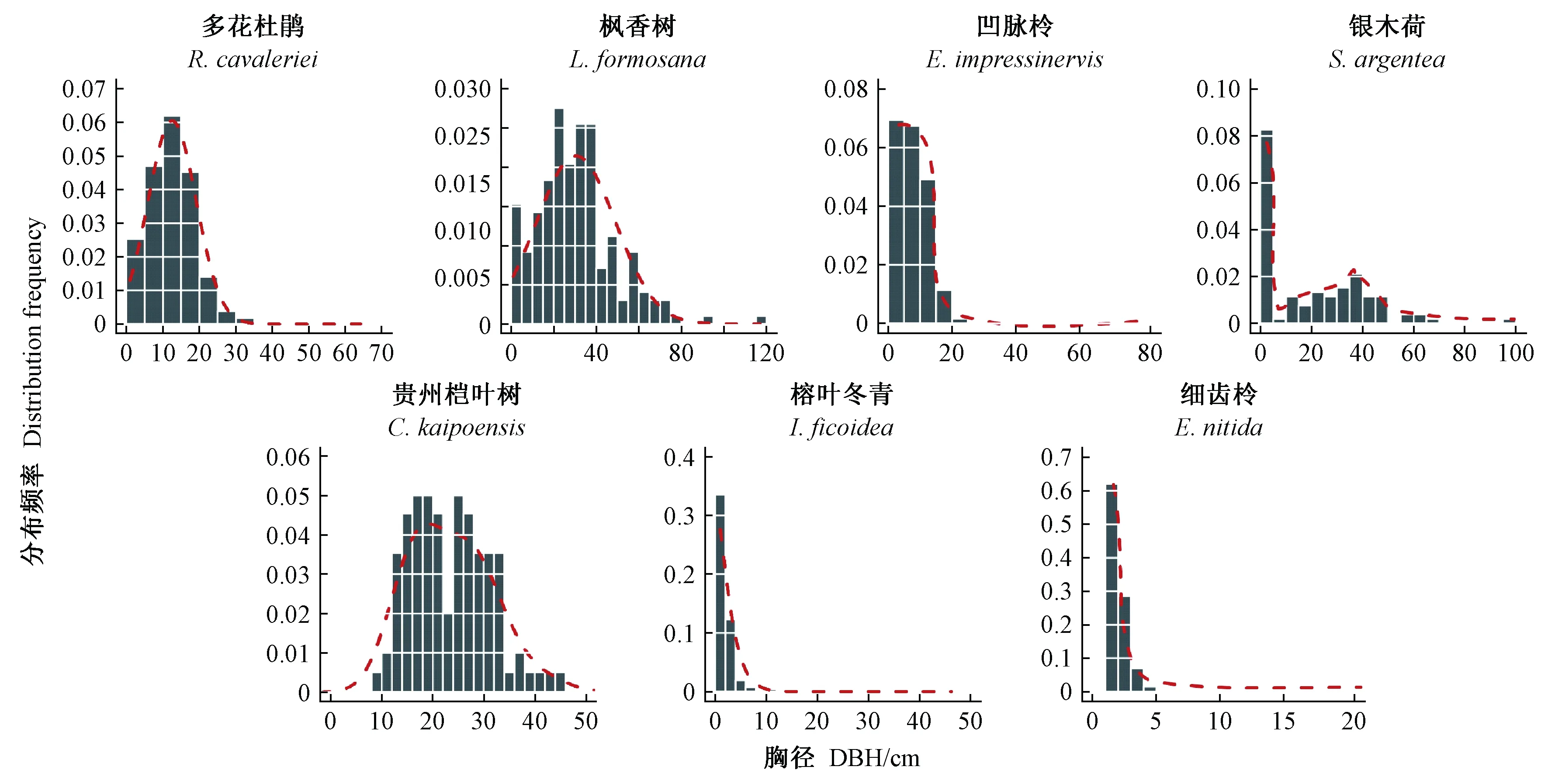
图4 样地内主要树种径级分布Fig. 4 DBH size-class distributions of the main tree species in the experimental plot
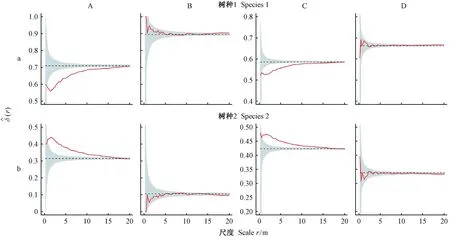
图5 模拟数据: 树种1和树种2的空间优势度Fig. 5 Simulation data: spatial dominance of species 1 and 2红线代表实际标记二阶特征函数值; 灰色区域表示零模型模拟的95%置信区间; 零模型将林木位置完全随机,但保持林木大小和树种属性不变; A列和B列为标记大小比数函数; C列和D列为标记双曲正切函数; A列和C列为实际空间优势度 (同时关注同种和异种邻体); B列和D列为只关注异种邻体计算的空间优势度。若观测值在上包迹线以上,表示周围分布较小的林木; 观测值在下包迹线以下,表示周围分布较大的林木,下同。Red lines represent the empirical marked second-order characteristic functions; grey areas represent 95% confidence intervals simulated by the null model; tree positions are generated following complete spatial randomness(CSR) but retaining tree size and species attributes unchanged in the null model; column A and B represent marked dominance functions; column C and D represent marked hyperbolic tangent functions; column A and C indicate the actual tree spatial dominance (all con- and heterospecific neighbors are considered) while column B and D indicate that only heterospecific neighbors are considered in the calculations. Nearest neighbors are often of smaller trees if the observed values are above the simulated envelopes, while nearest neighbors are of larger sizes if the observed values are below the simulated envelopes.
在实测样地中,对重要值排名前5位的较大径级树种(多花杜鹃、枫香树、凹脉柃、银木荷、贵州桤叶树)和典型小径级树种(榕叶冬青、细齿柃)的空间优势度进行分析(图6、7)。较大径级树种的空间优势度较高,在林分中占据优势地位(图6),且只考虑异种邻体计算的空间优势度高于实际值; 小径级树种则相反(图7)。同时,对关注树种的点状分布(图8)分析表明,7个树种均呈现出不同强度的聚集分布,说明种群的空间分布格局和种内种间关系对树种空间优势度具有显著影响。
较大径级树种(多花杜鹃、枫香树、凹脉柃、贵州桤叶树)的空间优势度均大于0.5(图6),在林分中较占优势,但其实际空间优势度在小尺度范围内显著低于随机区间,随着尺度范围增大接近期望值(图6A、C),说明树种周围分布着较大邻体降低了其邻域范围内的空间优势度; 若只关注异种邻体,四者空间优势度均趋近于随机区间,且高于实际空间优势度(图6B、D),说明种内聚集效应降低了大径级树种空间优势度。银木荷的空间优势度在小尺度范围处于随机区间内,而在大尺度范围高于随机区间(图6d),可能是由于生境异质性的影响,在大尺度上被较小树包围。
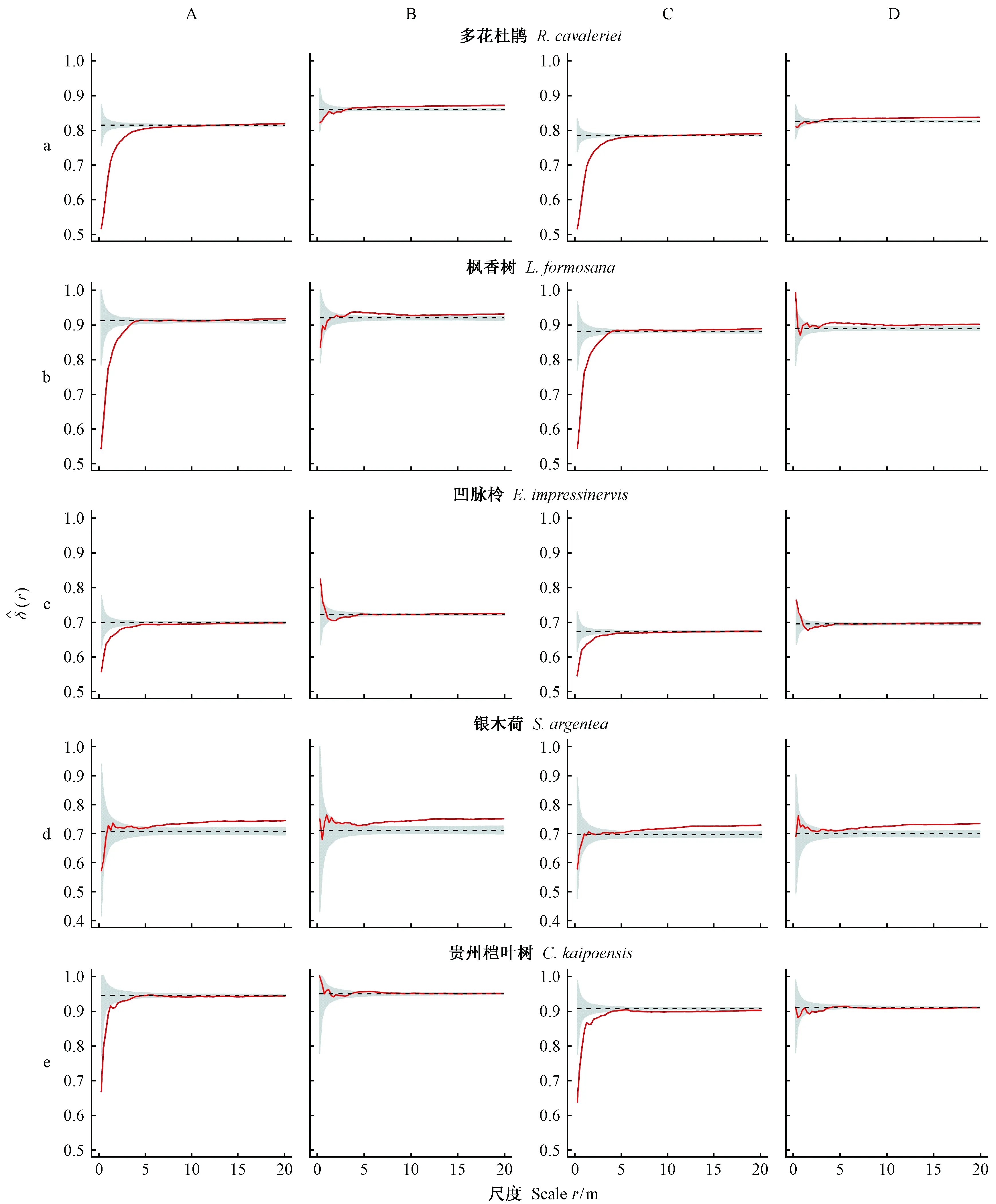
图6 较大径级树种的空间优势度二阶特征Fig. 6 Spatial dominance of trees of large-sized species
榕叶冬青和细齿柃等小径级树种的空间优势度较小,仅为0.2~0.5(图7),然而二者在不同距离尺度上的实际空间优势度均显著高于随机区间(图7A、C),说明树种周围分布邻体较小,增加了局部空间优势,随着尺度范围增大实际空间优势度接近于期望值。若只考虑异种邻体计算空间优势度,细齿柃在小尺度范围内观测值低于随机区间(图7Bb、Db),表明周围分布较大异种邻体,随着尺度范围增大空间优势度逐渐接近随机区间; 榕叶冬青在小尺度范围空间优势度高于随机区间,随尺度范围增大空间优势度接近期望值(图7Ba、Da)。同时,二者只关注异种邻体时的空间优势度低于实际值,表明同种聚集分布提高了小径级树种的空间优势度。
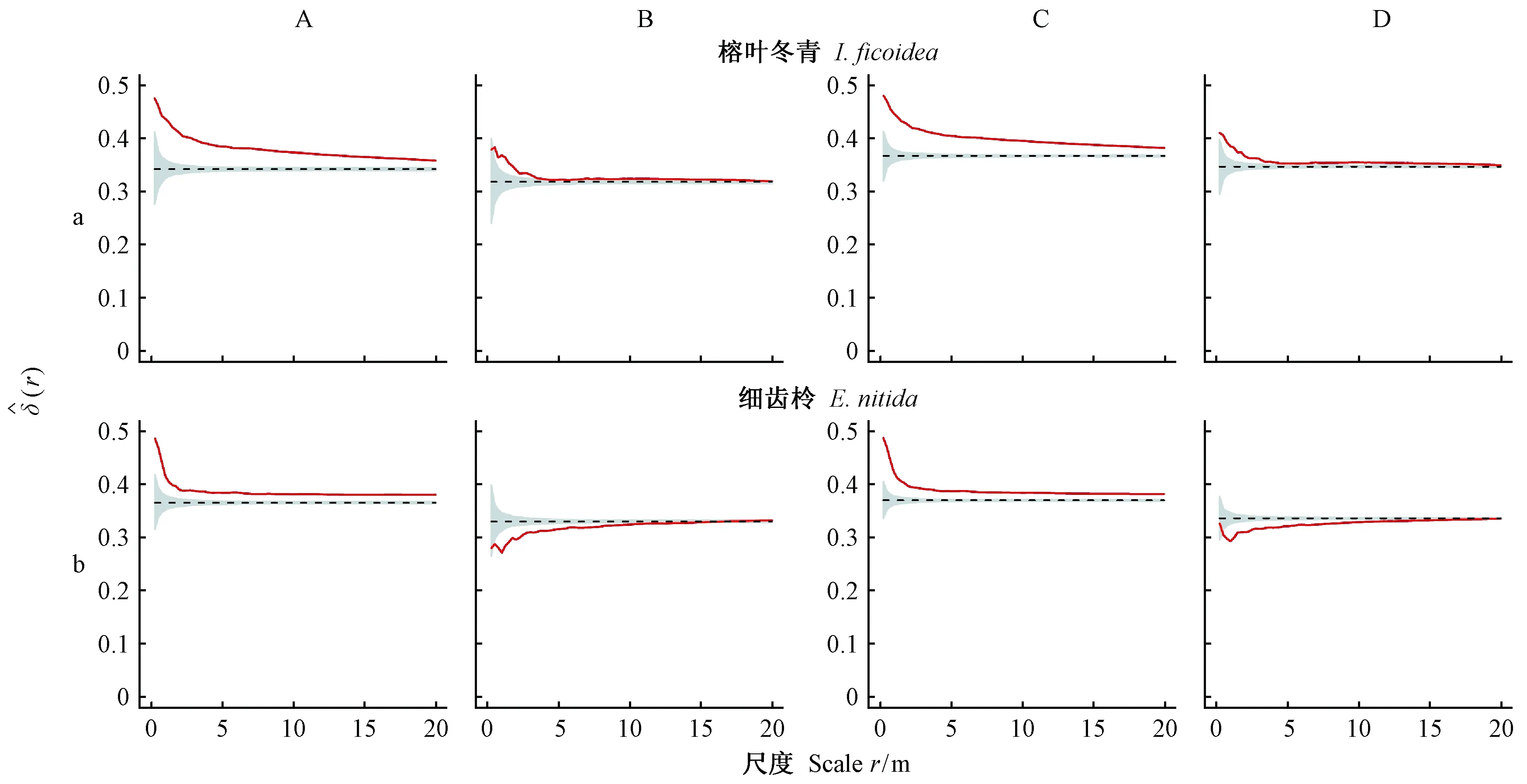
图7 较小径级树种的空间优势度二阶特征Fig. 7 Spatial dominance of trees of small-sized species
4 讨论
以往衡量树种优势度的指标多为重要值和基于最近相邻木关系构建的指数(刘宪钊等, 2009; 惠刚盈, 2013; 惠刚盈等, 2019),其中,重要值法计算简单,广泛用于评估植物物种在群落中的相对重要性(邵方丽等, 2012; 蔡军奇等, 2019); 基于最近相邻木关系构建的指数能够灵敏反映出树种空间优势度短期内的细微变化,在实际森林经营管理中易于实现(Szmyt, 2014; Lietal., 2014; 2020)。然而,重要值反映的是树种在群落中的数量对比关系,忽略了林木空间信息(惠刚盈, 2013; 惠刚盈等, 2019); 基于最近相邻木关系构建的指数虽体现了关注树种全部个体空间状态,但不能详细表达树种优势度随空间尺度的变化特征。本研究构造的2个标记二阶特征函数(标记大小比数和双曲正切二阶特征函数)可详细刻画某一树种在不同空间尺度下优势度的变化特征以及树种空间优势度偏离随机分布状态的程度,能够为长期监测林分中主要树种以及稀有树种的空间优势度提供更为有效的途径。
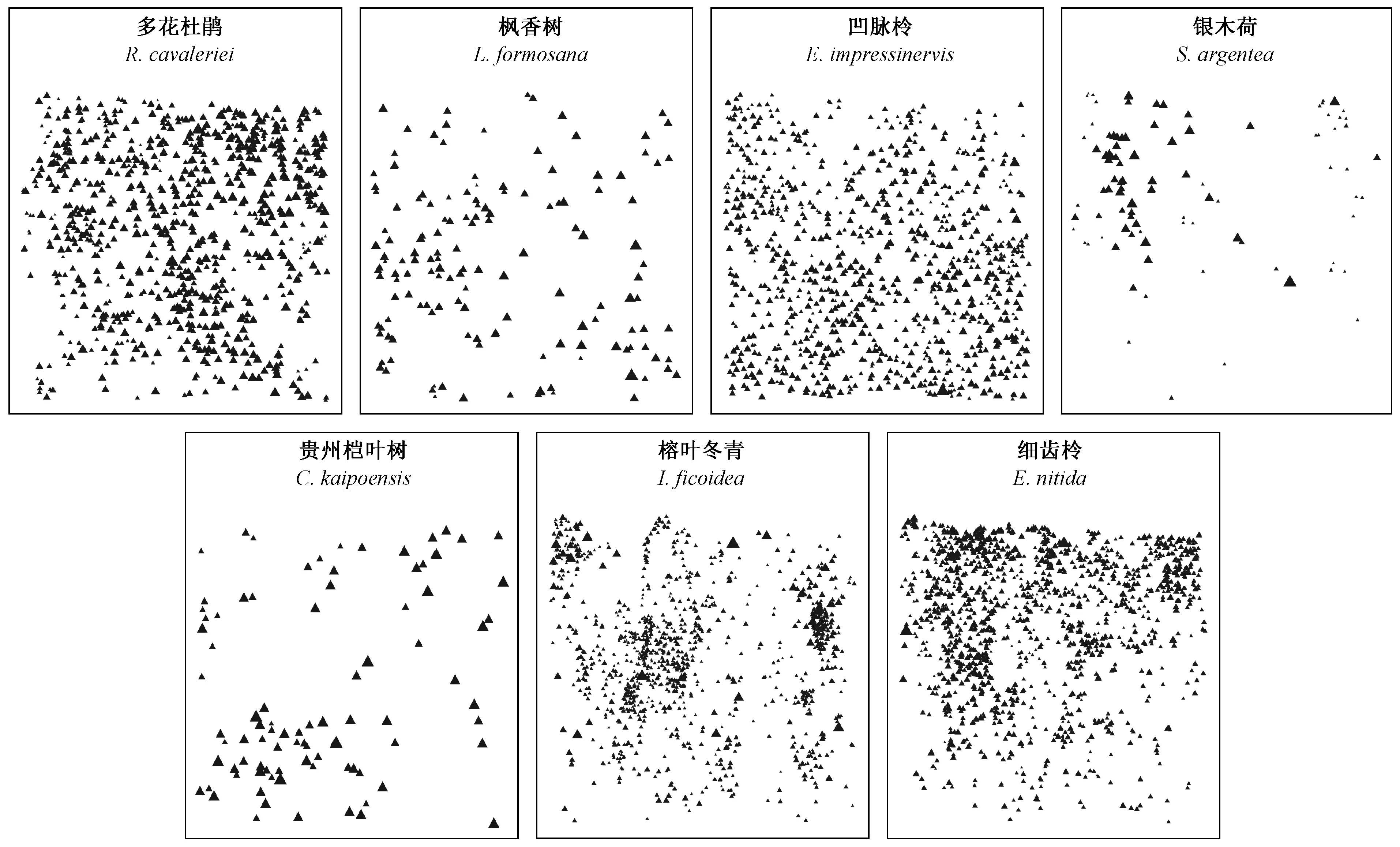
图8 样地内主要种群点状分布Fig. 8 Tree spatial distributions of the main species populations in the experimental plot三角形表示树木空间相对位置,三角形的大小表示林木个体直径相对大小。Triangle represents tree spatial positions, and the sizes of triangles indicate the relative diameter of the individual trees.
群落中树种空间优势度与树种空间分布格局和树种混交之间存在复杂的内部联系。树种空间分布格局影响种内种间林木个体相互作用的强度,聚集性强的树种种内竞争作用大于种间,聚集程度弱的树种更容易受到种间竞争的影响(Stoll, 2001; Semchenkoetal., 2013)。在天然林中,因种子扩散限制和生境异质性的影响,自然群落种群通常为聚集分布,同种个体易于紧邻在彼此周围(Conditetal., 2000; Volkovetal., 2009; Wangetal., 2010; Linetal., 2011; Lara-Romeroetal., 2016),从而导致树种间的空间隔离,降低了树种混交程度。一般来说,异种邻体之间相互作用强度较弱,竞争排斥作用较小,而同种邻体之间竞争排斥作用明显(祝燕等, 2011),因此,是否关注同种邻体计算的空间优势度反映了树种“空间隔离”效应 (Dettoetal., 2016)对物种多样性维持的贡献。在模拟林分和实测林分中,树种“空间隔离”效应导致的低混交度使得种内竞争强度大于种间,此时消除或减缓优势树种对劣势树种的排除作用,有利于维持物种共存,提高森林群落物种多样性(Murrelletal., 2001)。
较大径级树种在占据空间能力上存在明显优势,具有很强竞争能力。若只考虑异种邻体计算大径级树种空间优势度,由于树种竞争能力强且空间优势度较高,偏离随机区间并不明显。同时关注同种和异种邻体时,树种聚集分布造成种内竞争作用加强,大径级树种整体空间优势度会明显降低,且在小尺度上的局部空间优势度显著下降。由此说明,聚集分布格局使得大径级树种与其他树种(尤其是小径级树种和稀有树种)发生相互作用的机会减少,导致树种“空间隔离”效应(Chesson, 2000b),该效应一定程度上限制了大径级树种在群落中的绝对优势,从而减轻了对较劣势树种和稀有树种的竞争排除作用,达到维持物种共存的目的。对小径级树种来说,当只关注异种邻体时,细齿柃在小尺度上空间优势度低于随机区间,表明该树种周围分布着较大异种树,且在林分中受到较大竞争压力; 然而,小径级树种实际空间优势度较只关注异种邻体空间优势度明显提高。图8显示较小径级树种的分布格局为聚集分布,由于其胸径较小且处于林冠下层,故林木个体间为了争夺一定的生长必需空间和资源彼此会产生激烈的竞争。虽然小径级树种空间优势度较低,个体在群落中处于竞争劣势,但由于其往往呈现较强聚集性,从而减小了种间竞争排斥作用,相应地提高了较小径级种群空间优势度(毛子昆等, 2020),促进了森林中树种的多样性(Clark, 2010)。
林分空间结构之间是相互联系、相互制约的有机整体(惠刚盈等, 2018)。例如,一些研究从林分空间结构参数之间的联合概率分布等探讨林分结构的不同方面,分析林木空间分布格局、树种混交以及林木大小分化之间的关系(Lietal., 2012; 毛沂新等, 2019),然而,过去多关注的是基于群落水平的林木空间分布格局。本研究认为种群空间分布格局是影响其空间优势度和混交程度的重要原因,同时,决定种群空间分布的过程(如种子扩散限制、生境异质性等)对其空间优势度存在显著影响。需要说明的是,本研究仅基于大小比数和双曲正切指数表达树种空间优势度,探讨其与空间分布格局以及隔离程度之间的关系,尚有待于从结构的不同方面(如林木大小分化、物种分离指数等)全面探究林分空间结构不同维度之间的关系。
5 结论
标记大小比数函数和标记双曲正切指数函数能精细刻画不同空间尺度上所关注树种的空间优势度,且通过分析不同树种空间优势度发现,种群的空间分布格局、混交程度、空间优势度之间存在一定关联性,即同种聚集分布导致的树种空间隔离可影响树种空间优势度。对较大径级树种,聚集分布导致其空间优势度降低,而小径级树种则相反,该结论表明树种“空间隔离”效应导致的种内种间关系对森林群落中树种多样性维持具有重要作用。在今后的森林经营实践中,标记二阶特征方法可为分析群落中主要树种和关键树种的空间优势程度及监测其动态变化提供有效途径。
猜你喜欢
广西林业科学(2022年4期)2022-11-07
林业调查规划(2022年3期)2022-06-13
军事文摘(2021年22期)2022-01-18
昆明医科大学学报(2021年10期)2021-12-02
建材发展导向(2021年20期)2021-11-20
检察风云(2021年16期)2021-10-29
园艺与种苗(2021年3期)2021-05-26
上海理工大学学报(2020年2期)2020-05-30
华东师范大学学报(自然科学版)(2020年1期)2020-03-16
种子(2019年6期)2019-07-13