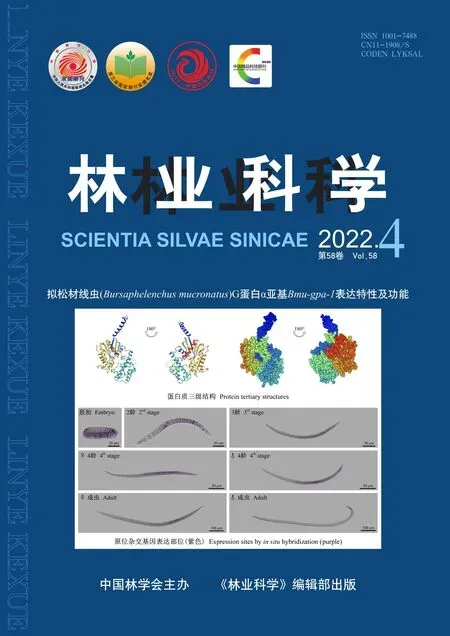
拟松材线虫G蛋白α亚基Bmu-gpa-1表达特性及功能*
2022-07-20 06:07刘文义余红仕周立峰胡加付
林业科学 2022年4期
马 芮 刘文义 余红仕 周立峰 胡加付
(浙江农林大学 杭州 311300)
拟松材线虫(Bursaphelenchusmucronatus)是一种松树寄生线虫,与外来物种松材线虫(B.xylophilus)同属于松材线虫病害系统,已在国内外造成严重的生态损失及经济损失(汤勤等, 1989; 张治宇等, 2004; 汤坚等, 2008; Sultanaetal., 2013)。过去对拟松材线虫病害没有引起足够的重视,但近年来越来越多的研究报告显示拟松材线虫也具有较高的致病性(陈凤毛等, 2010; 周立峰, 2017; 薛美静等, 2019)。高效频繁的交配行为是拟松材线虫和松材线虫快速繁殖及病虫害发生的基础,研究其交配繁殖机制有着深远的意义(徐红梅等, 2017; Liuetal., 2019)。迄今为止,国内外关于拟松材线虫的研究多侧重于与松材线虫的比较鉴定,而关于拟松材线虫交配繁殖学的研究较少,特别是关于拟松材线虫交配繁殖分子机制的研究。本课题组前期对拟松材线虫进行转录组测序,在拟松材线虫转录组数据中挖掘到了21个与线虫交配繁殖有关的G蛋白α亚基(Gα)家族成员,转录组结果显示Bmu-gpa-1基因在拟松材线虫交配前后表达差异显著,且表达模式具有雌、雄虫差异性,这为笔者探究拟松材线虫的交配繁殖分子机制提供了依据。
G蛋白,即鸟苷酸结合蛋白,由α、β和γ这3个亚基构成,是一类重要的信号转导蛋白,普遍存在于真核生物细胞中(周宝宏,1989)。G蛋白是G蛋白偶联受体(GPCRs)联系胞内信号通路的关键蛋白,在线虫的生长发育、运动取食、交配繁殖等方面发挥着重要作用( Tall, 2002; Reynoldsetal., 2004)。模式生物秀丽隐杆线虫(Caenorhabditiselegans)中共有21个Gα、2个Gβ及2个Gγ(Cuppenetal., 2003; Jansenetal., 1999),其中Gα是激活G蛋白信号传导的关键亚基,是主要的受体接触位点及GTP结合位点。当化学信号激活GPCRs后,GPCRs会诱导G蛋白的α亚基与β、γ亚基,导致GDP解离,最终结合GTP并激活下游效应蛋白,参与线虫的各种生理活动(Mendeletal., 1996; Yoshidaetal., 2012)。
在秀丽隐杆线虫中,整个gpa基因家族均参与线虫的化学感应系统,除gpa-12外其余的gpa基因都在化学感应神经元上集中表达。研究发现,多个gpa基因可以在同一个神经元中共表达,比如gpa-2、gpa-3、gpa-5、gpa-6、gpa-13及odr-3基因在AWC神经元中表达,可以负向调控线虫的感知系统。另外,同一个基因也可在多个神经元上表达,比如gpa-3基因可以在ASE、AWA神经元和ADL神经元上表达;gpa-1基因可以在ADL、ASH、ASI、ASJ、PHA、PHB神经元及雄虫尾部的特定p.c.s.神经元中表达,调节线虫对于氯化钠、异戊醇和乙醇等化学物质的趋向躲避行为以及雄性线虫的交配行为(Afsharetal., 2004)。研究发现过量表达雄性秀丽隐杆线虫的gpa-1基因,其性功能急剧下降,出现尾部翻转异常、交合刺插入困难等现象,但干扰沉默该基因却未发现异常(Jansenetal., 1999; Lansetal., 2004)。
为了探究拟松材线虫种内交配繁殖机制,本文对拟松材线虫Bmu-gpa-1基因进行了功能分析。通过生物信息学分析,概括其与其他种属线虫的亲缘关系,明确进化地位; 利用原位杂交、转录组测序分析及实时荧光定量PCR技术研究该基因的表达特性,确定其在线虫交配前后及不同性别上的表达差异性; 采用RNA干扰技术对Bmu-gpa-1的生物学功能进行研究,明确其在线虫交配行为及繁殖力上的作用,这即可为进一步探究G蛋白信号转导在线虫中的功能提供参考,也可为揭示2种线虫的种间竞争机理提供分子基础。
1 材料与方法
1.1 供试线虫培养
供试拟松材线虫由安徽省南陵市罹病马尾松中分离获得,25 ℃避光条件下在灰葡萄孢(Botrytiscinerea)菌苔上培养。用贝尔曼漏斗法收集大量怀卵雌虫,使其避光产卵1 h,可获得同步胚胎,孵育29 h后可获得同步2龄(J2)幼虫。将同步J2幼虫接入灰葡萄孢中,25 ℃培养35、62、82 h,即可获得同步生长的3龄(J3)幼虫、4龄(J4)幼虫及成虫。收集同步J4虫,根据性别形态差异挑选雌、雄虫,分别饲养至成虫,即可获得未交配的雌、雄成虫。
1.2 Bmu-gpa-1基因克隆与序列分析
使用TRIZOL法提取拟松材线虫总RNA,使用PrimeScript RT reagent Kit with gDNA Eraser试剂盒(Takara)将拟松材线虫总RNA反转录为cDNA。PCR扩增引物为Bmu-gpa-1-NF: 5′-GGCGTATCA ATGACTCTA-3′,Bmu-gpa-1-NR:5′-TTGTATCCATG ACCGAAT-3′,扩增体系及条件参照Zhou等(2020)前期报道。将PCR 扩增产物克隆在pGEM®-TEasy载体上,送往北京擎科生物有限公司测序。根据测序结果利用NCBI保守域数据库确定其保守结构域,使用DNAMAN8软件进行氨基酸序列比对,通过 MEGA 7.0邻接法进行系统发育分析。在SWISS MODEL(https:∥swissmodel.expasy.org)网站上预测编码蛋白的三级结构,并利用ExPASy(https:∥web.expasy.org/protparam/)预测蛋白质分子量、亲水性及稳定性。
1.3 Bmu-gpa-1表达模式研究
利用荧光定量PCR(RT-qPCR)技术分析Bmu-gpa-1基因在拟松材线虫不同发育时期的表达水平,利用原位杂交技术在mRNA水平上研究该基因在线虫不同发育时期的表达部位,明确其时空表达特性。RT-qPCR引物为Bmu-gpa-1-qF:5′-TAAAAC GGCCAAAAAGCGCA-3′,Bmu-gpa-1-qR:5′-TCTCTT TATTGCGACGGGCA-3′,内参基因选择18S及TUB(周立峰, 2017),具体步骤参照Zhou等(2020)前期报道。
1.4 Bmu-gpa-1生物学功能分析
1.4.1 dsRNA合成及RNAi浸泡试验 使用MEGAscript ®试剂盒进行体外转录反应合成dsRNA。引物为Bmu-gpa-1-iF: 5′-TAATACGACTC ACTATAGGGAGAGGCGTATCAATGACTCTA-3′,Bmu-gpa-1-iR: 5′-TAATACGACTCACTATAGGGAGATTGT ATCCATGACCGAAT-3′。将5 μL的拟松材线虫混悬液放入15 μL的Bmu-gpa-1-dsRNA干扰液中黑暗浸泡处理30 h,再将2批等量的线虫分别转移至15 μL的gfp-dsRNA干扰液和15 μL不含有dsRNA的缓冲液中黑暗浸泡处理30 h,作为对照组。每组均含有3 μL的M9 Buffer,干扰液中的dsRNA终浓度为0.5 μg·μL-1,最后用ddH20定容(陈莎妮等, 2021)。
1.4.2Bmu-gpa-1干扰效率检测 提取对照组、处理组线虫的RNA,反转录为cDNA,应用RT-qPCR技术检测各个发育阶段拟松材线虫的RNAi效率,每次重复4组,共3次。
1.4.3 交配行为测定 收集未交配的同步雌、雄虫,按照对照组(1条正常雌虫+1条正常雄虫)、雌虫组(1条干扰后雌虫+1条正常雄虫)、雄虫组(1条正常雌虫+1条干扰后雄虫)设置不同试验组,每次试验重复8组,共3次。观察记录每组线虫在交配高峰期(8 h)内的交配时长及交配次数,24 h后进行胚胎计数。计算每组线虫的交配率(交配组数/试验总组数)、平均交配次数(交配总次数/交配组数)、平均交配时长(交配总时长/交配组数)、有效交配率(产卵组数/交配组数)、平均产卵量(产卵总量/有效交配组数)等参数,用以评估每组线虫的交配能力及繁殖能力(赵双修, 2016; 徐红梅等, 2017)。本研究中,正确交配行为定义为: 雄虫的交合刺准确插入雌虫的阴户,持续时长不低于1 min。有效交配行为定义为: 拟松材线虫正确交配,并产下后代(赵双修, 2016)。
2 结果与分析
2.1 Bmu-gpa-1基因的克隆与序列
拟松材线虫Bmu-gpa-1全长1 780 bp,其中CDS为1 065 bp,编码354个氨基酸。ExPASy-ProtParam tool预测基因编码蛋白的分子式为C1822H2918N492O536S17,具有亲水性,不稳定指数(instability index,Ⅱ)为47.4,是一类不稳定蛋白。NCBI Conserved Domains预测其主要功能区为G1-5box、SwitchⅠ-Ⅱregions,它们是GTP结合位点、Gβγ和下游效应器的结合位点,在各种生理活动中起调节的作用,如图1A所示。在SWISS MODEL上预测其编码蛋白的三级结构,结构图如图1中D1、D2所示。结果显示该基因编码蛋白结构与典型的异三聚体G蛋白晶体结构1got.1最为相似,GMQE为0.74,QMEAN为0.70±0.05,蛋白序列一致性为44.06%,这表明预测结果可信度较高,蛋白模型匹配度较好,该基因编码蛋白属于G蛋白家族(Lambrightetal., 1996)。利用 DNAMAN软件将Bmu-gpa-1基因所编码的氨基酸序列与Bursaphelenchusxylophilus(CAD5232120.1)、Caenorhabditiselegans(NP_505840.1)、Caenorhabditisbriggsae(XP_002636462.1)、Strongyloidesstercoralis(AAK54043.1)、Strongyloidesratti(XP_024503395.1)、Loaloa(XP_003135770.1)、Brugiamalayi(CDP98783.1)及Brugiatimori(VDO30356.1)的氨基酸序列进行多重序列比对。结果如图1B所示,Bmu-gpa-1基因的氨基酸序列与不同物种的gpa-1基因的氨基酸序列具有较高的保守性。利用MEGA软件,采用邻接法构建系统发育树。该进化树包含Bursaphelenchus、Strongyloides、Brugia等属,涉及了13个物种。结果如图1C所示: 拟松材线虫Bmu-gpa-1与其他线虫的gpa-1氨基酸序列聚类在一起,与另一个Gα基因egl-30氨基酸序列区别开来,表明Bmu-gpa-1基因与其他物种的gpa-1基因归属于同一个基因家族,区别于其他Gα基因。其中,Bmu-gpa-1基因与松材线虫gpa-1基因、秀丽隐杆线虫gpa-1基因亲缘关系较近,这为Bmu-gpa-1基因的生物学研究提供了参考。
2.2 Bmu-gpa-1基因的时空表达特性
拟松材线虫转录组测序分析及RT-qPCR结果如图2A、2B所示: 2组结果相一致,均表明Bmu-gpa-1基因在拟松材线虫J2及J3的表达量最高,与其他龄期相比差异显著(P<0.05),J4及成虫期表达量最低,且雄虫的基因表达水平显著高于雌虫,基因表达模式具有雌、雄虫差异性。这说明Bmu-gpa-1基因可能在拟松材线虫J2、J3时期发挥着重要的作用; 在时间上,随着线虫的发育,基因表达水平由低到高再到低变化。
原位杂交结果如图3所示,具体表达特性为: 随着胚胎发育,表达部位由散点至全胚胎变化(图A-D),J2时期全身表达(图E)、在J3时期的肠道、体壁肌肉及生殖原基处表达(图F)、在J4雌、雄虫的性腺表达(图G、H)。在成虫期,雌、雄差异表达: 雌虫只在阴户部位表达(图I),雄虫在交合刺及尾部表达(图J)。这说明Bmu-gpa-1在拟松材线虫各个发育阶段均有表达,且随着线虫的发育,基因表达部位从局部到全身再到局部变化,在成虫期雌、雄差异表达。由此笔者推测,在线虫的发育初期多部位表达的Bmu-gpa-1可能与线虫早期的生长发育有关,而在线虫的发育后期集中于性腺及尾部表达的Bmu-gpa-1可能与线虫后期的生殖发育有关,参与调控拟松材线虫的交配繁殖行为。

图1 Bmu-gpa-1生物信息学分析Fig. 1 Bioinformatics analysis of Bmu-gpa-1A: Bmu-gpa-1保守结构域; B: Bmu-gpa-1基因序列比对; C: Bmu-gpa-1系统发育分析。采用邻接法构建系统发育树,自检次数为1 000次,分支数字表示支持率; D1、D2: Bmu-gpa-1蛋白质空间结构。A: The conservative domain of Bmu-gpa-1 B: The sequence alignment of Bmu-gpa-1; C: Phylogenetic analysis of Bmu-gpa-1. The phylogenetic tree was constructed by neighbor-joining method with 1 000 self-checks, and the branch number indicated the support rate; D1,D2: The protein structure of Bmu-gpa-1.

图2 Bmu-gpa-1基因在拟松材线虫不同发育阶段的表达水平Fig. 2 Expression level of Bmu-gpa-1 in different stages of B. mucronatusA: 转录组分析; B: 实时荧光定量PCR测量; Embryo: 胚胎期; J2: 2龄时期; J3: 3龄时期; F-J4: 雌虫4龄时期; M-J4: 雄虫4龄时期; V-F-A: 未交配雌虫成虫期; V-M-A: 未交配雄虫成虫期; M-F-A: 已交配雌虫成虫期; M-M-A: 已交配雄虫成虫期。不同小写字母表示差异显著(P<0.05)。下同。A: Transcriptome analysis; B: RT-qPCR; Embryo: Embryonic stage; J2: The second stage of juvenile; J3: The third stage of juvenile; F-J4: The fourth stage of female juvenile; M-J4: The fourth stage of male juvenile; V-F-A: The adult stage of virgin female; V-M-A: The adult stage of virgin male; M-F-A: The adult stage of mated female; M-M-A: The adult stage of mated male. Different lowercase letters mean significant difference at 0.05 level. The same below.
2.3 Bmu-gpa-1生物学功能
2.3.1Bmu-gpa-1基因的RNA干扰效率分析 与对照组拟松材线虫(试验体系不含有dsRNA)相比,另一对照组(试验体系含有gfpdsRNA)线虫的Bmu-gpa-1基因表达略有下降,但差异不显著,而处理组(试验体系含有Bmu-gpa-1dsRNA)的拟松材线虫各龄期Bmu-gpa-1基因表达量均显著降低,表达水平均减少50%以上(图4),表明经RNAi处理后,拟松材线虫各龄期的Bmu-gpa-1基因表达水平受到显著抑制。

图3 Bmu-gpa-1原位杂交Fig. 3 In-situ hybridization of Bmu-gpa-1处理组: A. 囊胚期胚胎; B. 原肠期胚胎; C. 蠕虫期胚胎; D. 1龄幼虫; E. 2龄幼虫; F. 3龄幼虫; G. 4龄雌虫; H. 4龄雄虫; I. 雌性成虫; J. 雄性成虫; K. 雄性成虫交合部位。对照组. L. 原肠期胚胎; M. 雄性成虫; m. 食道球; v. 阴户; sp. 交合刺; an. 肛门。Treatment: A. Blastosphere; B. Gastrulae; C. Worm embryo; D. The first stage of juvenile; E. The second stage of juvenile; F. The third stage of juvenile; G. The fourth stage of female juvenile; H. The fourth stage of male juvenile; I. The female adult; J. The male adult; K. The copulative site of male adult. Control. L. Gastrulae; M. The male adult; m. Metacorpus; v. Vulva; sp. Spicules; an. Anus.

图4 利用荧光定量PCR分析RNAi效率Fig. 4 The RNAi efficiency quantified by RT-qPCREmbryo: 胚胎期; J2: 2龄时期; J3: 3龄时期; J4: 4龄时期; Adult: 成虫期。不同小写字母表示差异显著(P<0.05)。Embryo: Embryo; J2: The second stage of juvenile; J3: The third stage of juvenile; J4: The fourth stage of juvenile; Adult: The adult stage. Different lowercase letters mean significant difference at 0. 05 level.
2.3.2Bmu-gpa-1基因对拟松材线虫交配繁殖行为的影响 按照方法1.4.3进行试验,交配行为测定结果如表1所示: 对照组中,68.75%的拟松材线虫在8 h内完成交配,每组平均交配1.05次,平均交配时长为22.14 min,其中83.45%的交配行为是有效交配行为,平均每组雌虫产14.28颗卵。干扰雄虫组中,82.29%的线虫存在交配行为,每组平均交配1.47次,平均交配时长持续约15.19 min,但只有59.54%的交配行为属于有效交配行为,且平均每组雌虫产卵8.07颗。与对照组相比,干扰雄虫组的线虫交配次数显著增加,交配时长显著缩短,且繁殖力明显下降,但干扰雌虫组的线虫却无显著变化。推测Bmu-gpa-1基因可以调控拟松材线虫雄虫的交配行为,干扰该基因可使雄虫兴奋程度提高,表现出频繁快速低效的交配行为模式。
3 讨论
通过对拟松材线虫的转录组数据分析,筛选出线虫交配前后差异表达的Bmu-gpa-1基因,且该基因表达具有雌、雄虫差异性。通过生物信息学分析,笔者明确了Bmu-gpa-1基因属于G蛋白α亚基家族,与其他线虫的Gα具有较高的同源性,且拥有相同的代表性结构位点。构建的系统发育树显示拟松材线虫gpa-1基因与松材线虫gpa-1基因位于同一分支,且与秀丽隐杆线虫gpa-1基因关系较为紧密,这为笔者研究拟松材线虫gpa-1基因提供了参考,也为后续研究拟松材线虫和松材线虫种间竞争分子机制提供了理论支持。
本研究中,拟松材线虫Bmu-gpa-1基因的原位杂交结果与实时荧光定量PCR、转录组结果相一致。结果显示该基因在拟松材线虫各发育阶段均有表达,发育初期线虫全身表达,表达量较高,发育后期则主要在雌虫阴户、雄虫交合刺及尾部特异性表达,表达水平较低。笔者推测该基因可能与拟松材线虫的生长、生殖发育有关: J2、J3是线虫发育增长最快、形成生殖原基的时期(叶为民等, 1993),线虫大量取食以满足生长发育需求,此阶段的Bmu-gpa-1基因可能更多地在生长发育方面发挥作用,基因表达较高较广; 而线虫到了J4及成虫时期,线虫体内含有较多颗粒营养物质,足以维持正常的生长发育,雌、雄差异表达的Bmu-gpa-1基因则更多地在线虫的生殖发育方面发挥作用,基因表达相对较少。研究发现,秀丽隐杆线虫的gpa-1基因的表达水平也具有相似的特性,即各龄期广泛表达,表达水平大致随着线虫的发育而下降,且雄虫的基因表达水平显著高于雌雄同体线虫。在表达部位上,秀丽隐杆线虫也主要在头感器及尾感器上的ASI、ASJ、ADL、ASH、 PHA、PHB神经元及雄虫交配必需神经元p.c.s.上表达。Jansen(1999)也发现,秀丽隐杆线虫gpa-1、gpa-7、gpa-13、gpa-15基因也都在雄虫尾部特定神经元特异表达,这与笔者在拟松材线虫上取得的研究结果极为相似。

表1 交配行为测定结果①Tab.1 Results of mating test
借助于RNAi技术研究该基因的功能,发现拟松材线虫干扰后的雄性成虫表现出频繁交配,交配时长缩短及后代数量减少等交配质量下降特征,但对雌虫影响较弱。这说明gpa-1基因影响雄虫的交配行为及繁殖能力。在秀丽隐杆线虫中,过量表达gpa-1基因可以导致雄虫性功能急剧下降,雄虫无法准确定位雌虫阴户,交合刺插入困难,但干扰沉默该基因却未发现异常的交配行为(Jansenetal., 1999; Lansetal., 2004)。研究发现,干扰沉默秀丽隐杆线虫的另一个Gα基因goa-1出现类似情况,goa-1基因被沉默后,雄虫的交配效率显著下降,出现运动亢奋、性功能下降、繁殖力降低等现象,Mendel等(1996)认为这可能与该基因所表达的神经元、肌肉有关,这些特定神经元、肌肉可以影响线虫的交配繁殖行为。结合该基因表达模式,推测在拟松材线虫雄虫交合刺及尾部特异表达的Bmu-gpa-1基因可以调节雄虫的交配行为系统,维持正常的交配繁殖机制,保持种群稳定。
目前,国内外关于线虫交配繁殖学的研究多集中于交配行为的研究,在其遗传控制机制方面的研究鲜有报道。本研究通过对拟松材线虫Bmu-gpa-1基因的表达特性及生物学功能研究,为线虫Gα基因的研究提供方法参照及理论参考,也对揭示拟松材线虫种内交配机制具有重要意义。同时,国内外关于拟松材线虫与松材线虫的种间竞争研究也多集中于繁殖差异性上,进一步与姊妹种松材线虫的比较,可为两者种间竞争机制的研究提供依据,也可为阐明松材线虫病害机理及调控基础提供重要突破口。
4 结论
拟松材线虫Bmu-gpa-1基因是G蛋白α亚基家族中的一员,在拟松材线虫各发育阶段均有表达,发育初期被广泛表达,表达水平较高,发育后期出现雌、雄虫差异表达,表达水平较低。同时,Bmu-gpa-1基因在功能上也具有雌、雄虫差异性,对雌虫影响较弱,但可以影响其交配繁殖行为,这为笔者研究线虫的种内交配分子机制提供了基础,也为拟松材线虫的病害防治提供了新思路,后续将继续探究Bmu-gpa-1与Bxy-gpa-1的异同及其在种间竞争中发挥的作用,为进一步揭示拟松材线虫生态位丧失及松材线虫入侵机制奠定基础。
猜你喜欢
广西林业科学(2022年1期)2022-03-03
作物研究(2021年6期)2021-12-25
今日农业(2021年16期)2021-11-26
甘肃农业科技(2021年6期)2021-07-25
新农民(2020年5期)2020-12-10
绿色中国(2019年13期)2019-11-26
江西农业学报(2019年10期)2019-10-29
科学导报(2019年35期)2019-09-03
浙江林业(2015年10期)2015-12-11
江苏农业科学(2014年3期)2014-07-16