
黑素瘤小鼠脾脏淋巴细胞抗肿瘤效应及机制的初步探究
2022-07-13 13:59邹锐涛蔡桂月陈晓旋樊志金陈嵘祎
中国麻风皮肤病杂志 2022年9期
邹锐涛 蔡桂月 陈晓旋 肖 铿 樊志金 陈嵘祎
1广东医科大学,广东广州, 523024;2南方医科大学皮肤病医院,广东广州,510091
恶性黑素瘤(malignant melanoma,MM)是起源于黑素细胞的一种恶性肿瘤,具有侵袭性强,如不及时治疗预后差等特点[1]。在现阶段,MM常规治疗的疗效不佳,其中手术仅对早期治疗有效。近年来,免疫治疗在MM中发挥着越来越重要的角色。目前,应用在MM中的免疫治疗主要有:免疫检查点抑制剂(PD-1和CTLA-4等单克隆抗体);细胞因子(IL-2,IFN-α-2b等);溶瘤病毒(溶瘤病毒T-VEC);肿瘤疫苗(肽疫苗、DC疫苗等);过继性细胞疗法(肿瘤浸润淋巴细胞疗法、工程T细胞受体治疗、嵌合抗原受体T细胞疗法、NK细胞疗法等)[2]。其中,过继性细胞疗法是近几年来在肿瘤免疫治疗中的一大热点。
过继性细胞疗法(adoptive cell therapy,ACT)是通过收集患者体内的免疫细胞,经过体外激活或基因修饰,扩增至足够数量时并选择具有肿瘤靶向的免疫细胞,再回输到机体从而起到抗肿瘤的一种新型的免疫治疗方法[3,4]。这些免疫细胞可来源患者外周血淋巴细胞、肿瘤浸润淋巴细胞(tumor infiltrating lymphocytes, TIL)或淋巴因子激活的杀伤细胞(Lymphokine activated killer cell, LAK)。嵌合抗原受体T细胞免疫疗法(CAR-T)是对外周血中淋巴细胞进行磁珠分选T细胞后再进行基因编辑成CAR-T,对于肿瘤治疗具有精准、快速、高效,但对于实体瘤的治疗效果欠佳,在MM治疗中的研究较少;TIL是从肿瘤组织中提取出来的,这类淋巴细胞具有精准识别肿瘤细胞的能力,能够有效地杀伤肿瘤细胞,对于转移性黑素瘤具有较好的治疗效果[5-7]。但是TILs量少,分离困难,很多情况下是功能耗竭的,无法发挥正常的抗肿瘤作用,发展新的淋巴细胞来源具有重要意义。
本研究通过构建黑素瘤小鼠模型,提取荷瘤小鼠脾脏淋巴细胞,与黑素瘤细胞共培养,发现与正常脾脏淋巴细胞相比,来自黑素瘤小鼠的脾脏淋巴细胞具有更明显的抑瘤现象,通过对两组淋巴细胞进行活化刺激,也发现黑素瘤小鼠来源的脾脏淋巴细胞分泌更高的肿瘤杀伤因子。脾脏淋巴细胞易获取、数量多,这一研究发现或许为MM的过继性细胞疗法提供新的思路。
1 材料和方法
1.1 材料
1.1.1 细胞系和实验动物 黑素瘤细胞系B16购自上海盖宁生物科技有限公司,雌性C57BL/6小鼠,SPF级,购自广东省医学实验动物中心;在南方医科大学动物伦理委员会指导下进行。
1.1.2 实验试剂 北美胎牛血清、1640培养基、0.25% Trypsin-EDTA (1X)0.25% Trypsin-EDTA (1X)胰酶(美国Gibco公司);PD-1、Granzyme B、perforin、GAPDH一抗、β-actin一抗(美国Cell Signaling Technology公司);Cyanine5-抗小鼠CD3、FITC-抗小鼠CD8、PE-抗小鼠CD4、PE-cy7抗小鼠Granzyme B、PE-抗小鼠perforin单克隆荧光抗体(美国Biolegend公司); CCK8试剂盒(同仁化学/DOJINDO);Calcein/PI 细胞活性与细胞毒性检测试剂盒、山羊抗兔二抗、BCA蛋白浓度测定试剂盒、红细胞裂解液(上海碧云天生物技术有限公司);刀豆蛋白A、Hass缓冲液(北京索莱宝科技有限公司);TNF-α、IFN-γ ELISA试剂盒(北京达科为生物技术有限公司)。
1.2 方法
1.2.1 细胞培养 小鼠黑素瘤细胞系B16,加入含10%胎牛血清、1%青霉素-链霉素双抗的1640培养基,置于37℃、5% CO2的培养箱中培养,长至70%~80%时,使用胰酶消化后传代以及液氮冻存保种。
1.2.2 荷瘤小鼠模型构建 B16细胞长至70%~80%时,胰酶消化后加入2 mL培养基重悬,离心5 min,去除上清,PBS清洗后用1 mL PBS重悬,计数,细胞量稀释至106/150 μL,小鼠背部备皮,每只每个部位注射150 μL,观察3周。
1.2.3 小鼠脾脏淋巴细胞提取 荷瘤小鼠建模3周后,肿瘤长至7~8 mm时,将正常小鼠与荷瘤小鼠进行解剖,取其脾脏剪碎置于Hass缓冲液中进行研磨,70 μm细胞筛过滤,使用淋巴细胞分离液进行提取,加入2 mL红细胞裂解液裂解5 min,离心并用PBS清洗后,使用1 mL完全培养基重悬,计数、备用。
1.2.4 Western blot 用刀豆蛋白A(ConA)5 μg/mL对淋巴细胞进行刺激,即将脾脏淋巴细胞分四组:正常小鼠脾脏淋巴细胞(NL)、肿瘤小鼠脾脏淋巴细胞(ML)、NL+ConA、ML+ConA,每组细胞数为2×106,铺板刺激24 h,收集细胞,每组用200 μL RIPA裂解液冰上裂解30 min,离心收集上清进行BCA蛋白定量,加1/4的5×loading buffer混匀后100℃金属浴10 min,每组取20 ug蛋白进行10%的十二烷基硫酸钠-聚丙烯酰胺凝胶电泳分离蛋白,湿转至PVDF膜上,用5%脱脂奶粉进行封闭1 h,TBST漂洗三遍,分别与PD-1、Granzyme B、perforin、GAPDH、β-actin等蛋白一抗(1∶1000)于4℃敷育过夜,TBST漂洗三遍后,加入二抗(1∶5000)室温敷育1h,TBST漂洗三遍后进行ECL 化学发光剂显色,Image J分析软件计算各组细胞目的蛋白的相对表达量。
1.2.5 ELISA 对上述分组细胞铺板刺激后离心收集上清,进行TNF-α、IFN-γ ELISA测定。严格按照说明进行检测,检测完成后,10 min内进行酶标仪测量每孔在450 nm发射波长下的吸光度值,标准曲线R2控制在0.998以上,根据标准曲线计算出TNF-α、IFN-γ表达水平。
1.2.6 Calcein/PI荧光染色 收集B16细胞进行24孔板铺板,每孔10万个细胞,4 h细胞贴壁后分3个组,每组3个复孔,即阴性对照组、正常组(NL组)、荷瘤组(ML组),按照1∶20(B16细胞:NL/ML)加入淋巴细胞,37℃、5% CO2培养箱中共培养24 h,去上清,PBS漂洗2遍,加入Calcein AM/PI染色液(1∶500),每孔200 uL,37℃、5% CO2培养箱避光孵育30 min,荧光显微镜观察。
1.2.7 CCK8 收集B16细胞进行96孔铺板,每孔10000个细胞,4 h细胞贴壁后共分为7个组、每组6个复孔。即空白组(无细胞)、阴性对照组、ConA 组、NL组、ML组、NL+ConA组、ML+ConA组,按照1∶20(B16细胞:NL/ML)加入淋巴细胞,ConA 5 μg/mL刺激24 h,每孔加入10 μL CCK8溶液后,37℃、5% CO2培养箱中孵育2 h,进行酶标仪测量每孔在450 nm发射波长下的吸光度值。
1.2.8 流式细胞术 取提前备好的NL/ML各106,分别用100 μL 1×的结合缓冲液重悬,每管分别加入5 μL的Cyanine5-抗小鼠CD3、FITC-抗小鼠CD8、PE-抗小鼠CD4流式抗体,4℃避光孵育20 min;而ConA刺激24 h后的NL/ML,分别用100 μL 1×的结合缓冲液重悬,每管分别加入5 μL的Cyanine5-抗小鼠CD3、FITC-抗小鼠CD8,4℃避光孵育20 min,漂洗后对细胞进行破膜,分别加5 μL PE-cy7抗小鼠Granzyme B、PE-抗小鼠perforin流式抗体,4℃避光孵育20 min,每管再加1mL缓冲液漂洗,离心后用500 μL 1×的结合缓冲液重悬,用流式细胞仪检测。
1.3 统计学方法 本研究使用Image J和GraphPad Prism 8.0进行制图以及实验数据的统计分析,两样本间采用配对样本t检验。检验水准α=0.05,P<0.05,表示差异具有统计学意义。所有实验结果均重复3遍以上。
2 结果
2.1 B16细胞分别与NL/ML共孵育24 h后各组B16细胞的增殖能力以及Calcein AM/PI荧光染色后的结果 NL和ML按照20∶1,分别与B16细胞共孵育24 h后,ML组与阴性对照组以及NL组相比较,B16细胞增殖能力均显著下降,差异具有统计学意义(*t=39.07,*P<0.05;#t=9.39,#P<0.05,图1a); Calcein AM/PI荧光染色后的结果也与之对应,并且NL与ML都依附在B16细胞表面(图1b)。
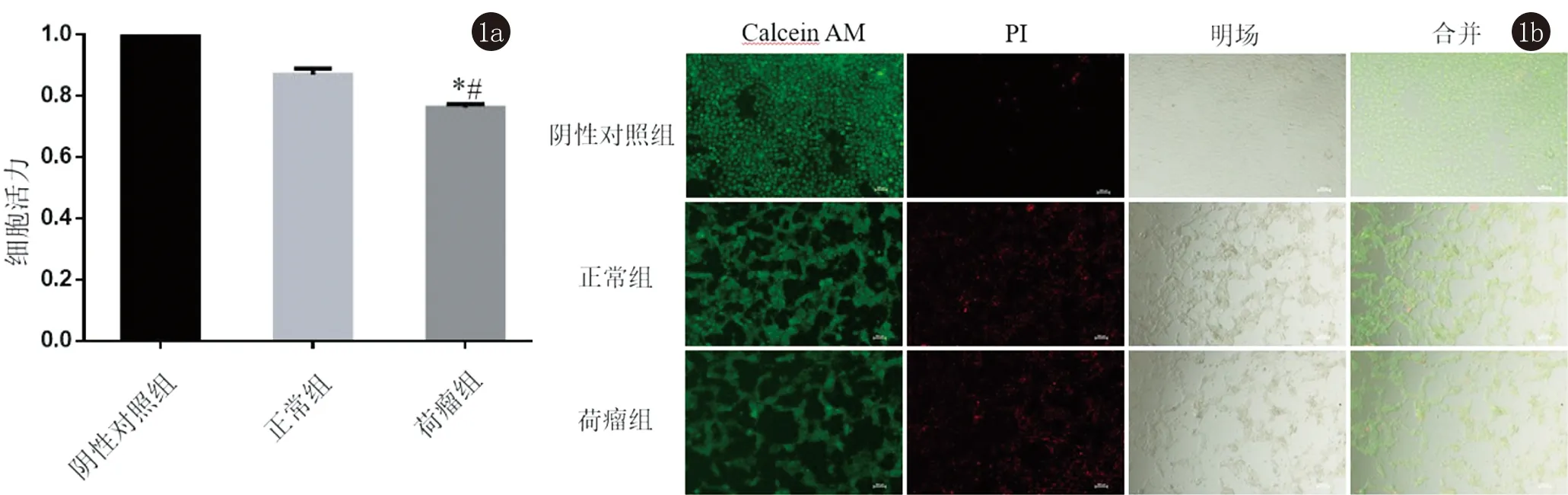
*P<0.05:与阴性对照组比较,# P<0.05:与正常组比较
2.2 B16细胞分别与NL/ML共孵育并加入ConA刺激24 h后各组B16细胞的增殖能力 B16 细胞按比例1∶20与NL/ML共孵育并加入ConA(5 μg/mL)刺激24h后,ConA组细胞增殖能力相对于阴性对照组无差异,排除ConA对于B16细胞的影响;ML组与阴性对照组以及NL组相比较,B16细胞增殖能力均显著下降(*t=69.61,*P<0.05;#t=5.44,#P<0.05,图2)。
2.3 NL组和ML组脾脏淋巴细胞中CD8+、CD4+T细胞比例占比 流式分析发现,在免疫T细胞表达CD3阳性条件下, NL组,中CD8+和CD4+T细胞比例分别为:25.1%、59.9%;在ML组中CD8+和CD4+T细胞比例分别为:31.3%、58.9%。ML组中CD8+T细胞明显高于NL组中CD8+T细胞,差异具有统计学意义(t=11.56,P<0.05,图3)。
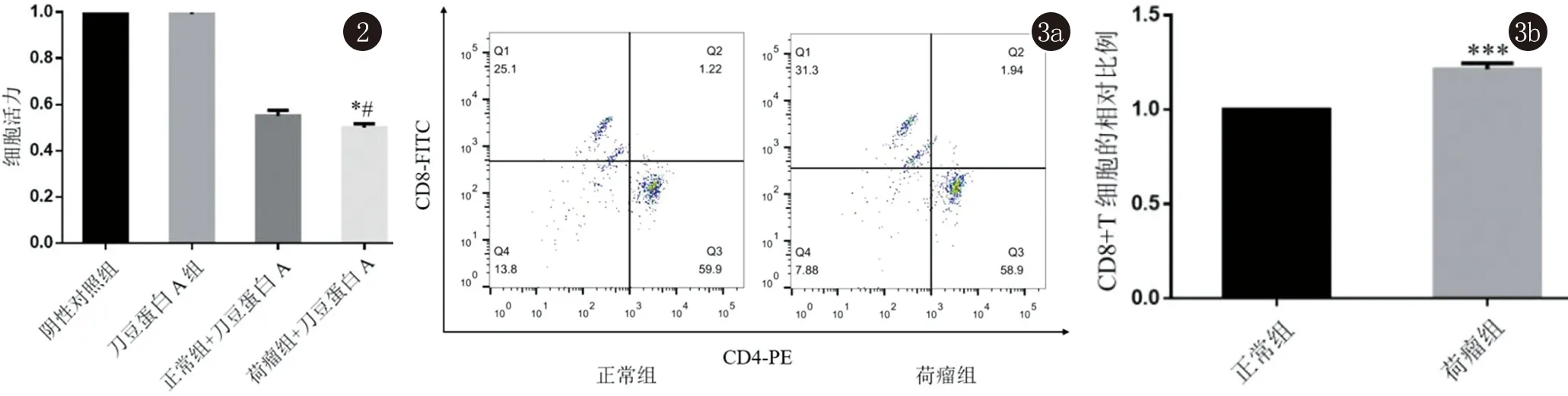
图2 CCK-8法检测细胞增殖能力 *P<0.05:与阴性对照组比较,# P<0.05:与正常组+刀豆蛋白A比较 图3 流式细胞术分析NL和ML中CD8+与CD4+比例占比 *** P<0.001
2.4 NL组和ML组PD-1蛋白表达水平 与正常小鼠脾脏淋巴细胞NL组相比,B16荷瘤小鼠脾脏淋巴细胞ML组中PD-1蛋白表达水平显著下调; 在ConA(5 μg/mL)刺激活化24 h后, ML组PD-1蛋白表达水平依旧较NL组显著下调(图4)。
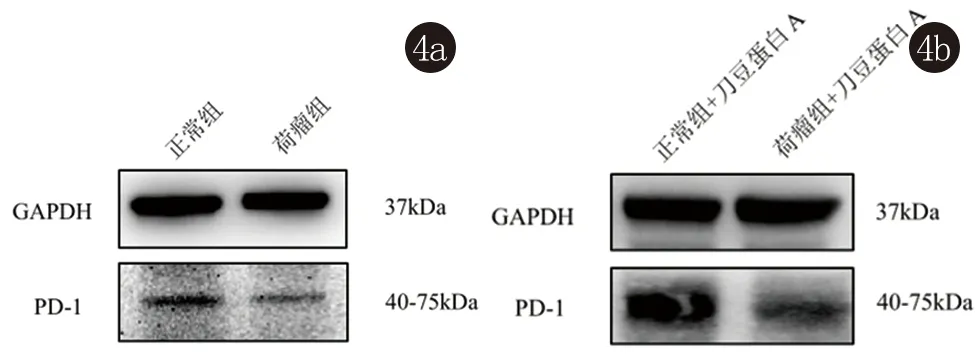
图4 Western blot检测NL、ML、NL +ConA和ML+ConA中 PD-1蛋白表达水平
2.5 在ConA刺激24 h之后,NL组和ML组淋巴细胞颗粒酶B、穿孔素蛋白表达水平以及TNF-α、IFN-γ细胞因子表达水平。
在ConA(5 ug/mL)刺激活化24 h后,ML组中颗粒酶B、穿孔素蛋白表达水平较NL组显著上调(图5a);TNF-α、IFN-γ细胞因子的表达水平都较NL组都显著上升(t1=22.72,P1<0.05;t2=11.56,P2<0.05,图5b);ML组中CD8+T细胞的颗粒酶B表达比例相对NL组更高,ML组(17.8%)较NL组(14.3%)提高了24.5%;而穿孔素的表达比例,ML组(11.3%)较NL组(8.78%)提高了28.7%,(图5c、5d),而未经ConA刺激的淋巴细胞几乎不表达颗粒酶B以及TNF-α、IFN-γ细胞因子。
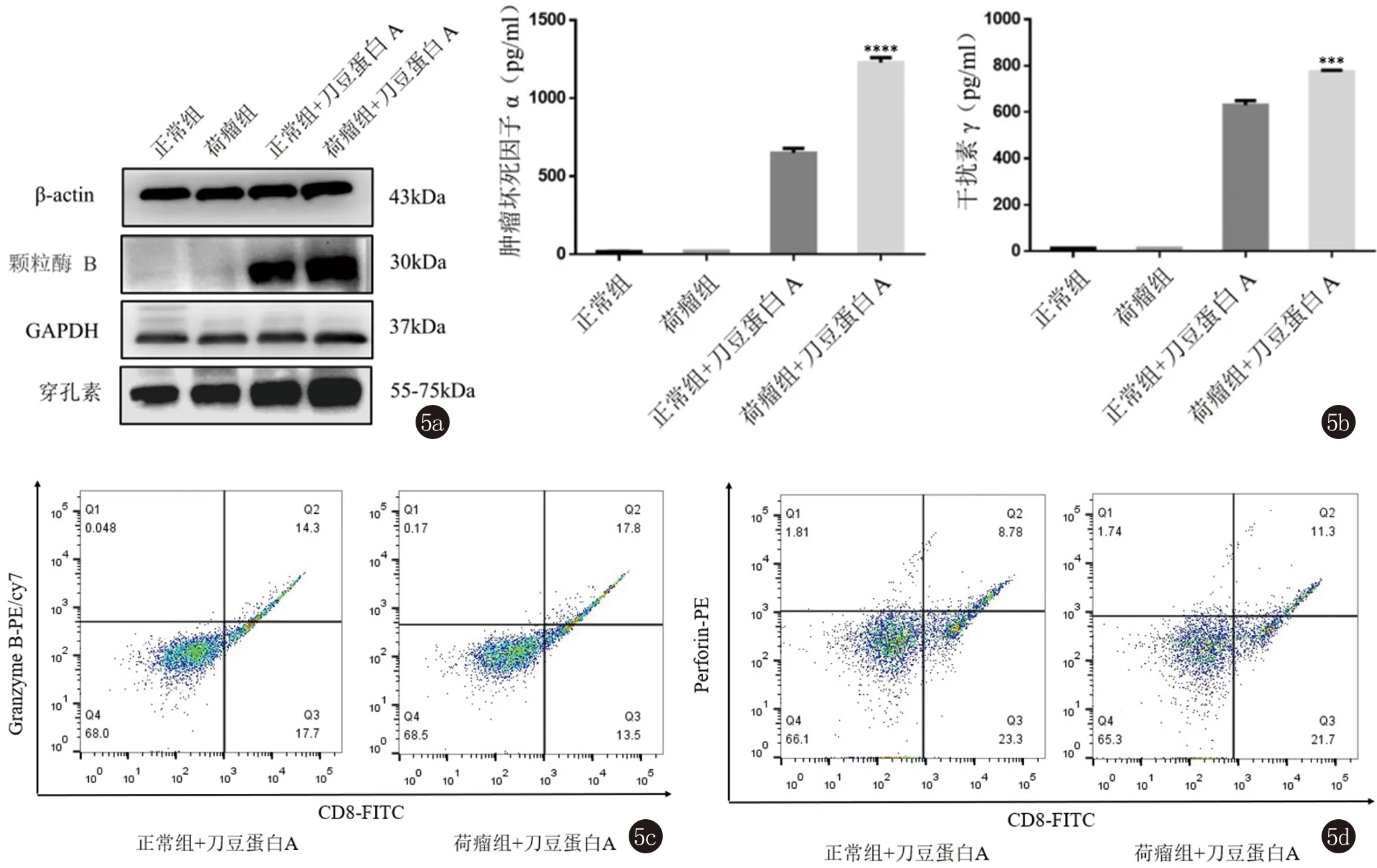
****P<0.0001;***P<0.001
3 讨论
MM极易发生侵袭且早期即可发生转移,常规治疗的疗效不理想,但MM是对免疫治疗相对敏感的恶性肿瘤,近几年免疫治疗在MM中的应用已经取得重大发展。免疫检查点抑制,特别是抗 PD-1 免疫疗法已经彻底改变了MM的治疗,美国 FDA 批准的免疫治疗药物 Pembrolizumab(PD-1抗体)/CTLA-4 抗体等用于治疗MM能明显延长欧美MM患者的生存时间[8,9]。PD-1(programmed death 1)程序性死亡受体1,是一种重要的免疫抑制分子。肿瘤微环境会诱导浸润的T细胞高表达PD-1分子,肿瘤细胞会高表达PD-1的配体PD-L1和PD-L2,导致肿瘤微环境中PD-1通路持续激活,T细胞功能被抑制,无法杀伤肿瘤细胞[10-12]。有研究发现,PD-1的相对水平与T细胞产生细胞因子的能力呈负相关,在PD-1上调的T细胞中发现TNF-α、IL-2和IFN-γ等细胞因子分泌功能受限[13]。Beane等[14]利用锌指核酸酶针对编码PD-1基因,敲低TIL中PD-1的表达,发现敲低PD-1的TIL体外效应功能得到改善,TNF-α、GM-CSF和IFN-γ等多功能细胞因子显著增加,在治疗转移性黑素瘤上也取得可观的效果,并且对机体不产生不利影响。有研究发现,体外刺激后,PD-1缺陷T细胞的IFN-γ分泌和细胞毒能力增加[15,16]。而TNF-α、IFN-γ在MM中发挥着重要作用,TNF-α在体内或体外均有杀死肿瘤细胞或抑制肿瘤细胞增殖的作用。IFN-γ具有抗病毒、抗肿瘤和免疫调控的作用,对于TNF-α具有刺激作用,增强抗肿瘤效应[17,18]。
PD-1虽然存在诱导表达的现象,即在活化的T淋巴细胞上高表达,但在正常的脾脏淋巴细胞中同样也表达PD-1[19],本实验研究发现ML中PD-1表达水平较NL显著下降,在ConA刺激24 h后,ML组中PD-1表达依旧较NL组中显著下降。并且在经过ConA刺激24 h之后,ML组中的TNF-α、IFN-γ表达水平均明显高于NL组(P<0.05),并且在CCK8细胞活力实验结果表明,NL和ML均具有抑制肿瘤细胞增殖的作用,但ML的抑制作用比NL更显著,在ConA刺激下,也证明了这一结果。这提示ML功能可能在肿瘤微环境中不易被抑制,并且分泌更高的细胞因子从而起到更好的杀伤肿瘤细胞的效果。
穿孔素是储存在细胞毒性T细胞(CTL)和NK细胞胞质的细胞毒颗粒中的糖蛋白,颗粒酶是CTL和NK细胞释放的细胞浆颗粒[20,21]。有研究表明,肿瘤免疫激活的CD8+T细胞主要通过穿孔素颗粒酶-和Fas/Fas配体-途径诱导细胞死亡来执行肿瘤清除[22], 本研究在CD3阳性分选情况下,通过流式分析发现,NL组中CD8+T淋巴细胞和CD4+T淋巴细胞比例分别为:25.1%、59.9%;在ML组中CD8+T淋巴细胞和CD4+T淋巴细胞比例分别为:31.3%、58.9%,其中ML组中CD8+T淋巴细胞比例占比明显高于NL组中CD8+T淋巴细胞(P<0.05)。而CD4+T细胞在两组中差异不大。并且在Western blot中发现,ML组中穿孔素的表达水平较NL组显著上升(P<0.05),而颗粒酶B在未激活的免疫细胞是不表达的;使用ConA刺激24 h后,ML组中穿孔素、颗粒酶B的表达水平也较NL组显著上升(P<0.05);流式分析发现ConA刺激24 h后,ML组中CD8+T细胞表达的颗粒酶B的比例也相对NL组更高,ML组(17.8%)较NL组(14.3%)提高了24.5%;而穿孔素的表达比例,ML组(11.3%)较NL组(8.78%)提高了28.7%。这也提示,ML相对于NL具有更强的抗肿瘤作用。
综上所述,黑素瘤小鼠的脾脏淋巴细胞具有更明显的抑瘤效应,可能通过低表达PD-1以及能够分泌更高的穿孔素、颗粒酶B以及TNF-α、IFN-γ等而实现。作为初步探究,我们在本研究中并未对这种现象是否具有特异性的进行验证,我们将在后面的研究中进行跟进。并且仅在细胞层面得出结论,仍需通过动物实验进一步验证。由于脾脏淋巴细胞易获取、数量多,希望这一研究发现能为MM的过继性免疫治疗提供新的思路。
猜你喜欢
结核与肺部疾病杂志(2022年4期)2022-11-21
中国医药科学(2022年5期)2022-05-05
中国药学药品知识仓库(2021年11期)2021-11-25
保健与生活(2020年5期)2020-03-20
中小学德育(2020年11期)2020-03-18
中国保健营养(2019年7期)2019-10-21
初中生·考试(2019年6期)2019-06-04
特别健康·下半月(2018年2期)2018-04-24
教育界·上旬(2016年12期)2017-05-25
