
单核苷酸多态性与肾透明细胞癌易感性的相关性研究
2022-07-05 03:13叶真逢习小庆黄高敏黄雅为汪建焜
江西医药 2022年5期
叶真逢,习小庆,黄高敏,黄雅为,汪建焜
(1.南昌大学第二附属医院泌尿外科,南昌 330008;2.景德镇第一人民医院泌尿外科,景德镇 333000)
肾细胞癌中肾透明细胞癌(Kidney Renal Clear Cell Carcinoma, KIRC) 最为常见,约占全部病例的80%[1-3]。 如何识别肾透明细胞癌高危人群,进一步降低发病率及死亡率, 是摆在我们面前的一个难题。 此外,不同群体及个体在相似暴露因素下对肿瘤发生与否及药物治疗反应存在较大差异, 这种差异被称为肿瘤遗传易感性[3]。 因此,准确的分子诊断和预后生物学指标的发展对肾透明细胞癌患者个体化和精准治疗至关重要。
单核苷酸多态性(Single Nucleotide Polymorphism,SNP) 是由单个核苷酸变异引起的DNA 序列多态性,是人类最常见的遗传变异类型。 SNP 是研究不同癌症或癌症特征的重要遗传标记, 被认为是潜在的癌变标志物, 因此对于癌症早期诊断和个性化的靶向治疗有价值[4-5]。 此外有研究表明肾透明细胞癌易感性与遗传SNP 有关, 如醛脱氢酶2(Aldehyde Dehydrogenase 2, ALDH2) 与多种肿瘤的发生和进展密切相关[6],编码PD-1 和PD-L1 分子的基因单核苷酸多态性与肾细胞癌发展相关,并影响肾癌患者预后[7]。 由于与癌症风险相关的SNP 可能会影响预后, 因此对相关SNP 分析可能有助于确定新的癌症预后生物标志物。 故本研究探究单核苷酸多态性与肾透明细胞癌易感性的相关性,旨在为准确识别肾透明细胞癌的高危人群提供科学依据。
1 材料与方法
1.1 TCGA 中突变和转录组数据获取 从公共开源的TCGA 中下载关于SNP 的原始数据格式Masked Somatic Mutation, 选择VarScan2 平台测序的SNP相关数据,并下载mRNA 原始表达数据,收集611份mRNA 数据,其中正常样本72 份,癌症样本539份。 从肾透明细胞癌样本SNP 数据中获得突变基因。将mRNA 原始数据用edger 软件包进行整合和标准化,得到差异表达基因及其表达水平。
1.2 TCGA 突变频率统计 应用perl 软件从原始SNP 相关数据中提取要可视化基因名称、染色体位置、突变类型及样本编号。 将准备好的SNP 突变频率数据用GenVisR 软件包进行整合和可视化,绘制瀑布图。
1.3 突变基因的功能富集和通路分析 为了解这些突变基因的功能,使用R 软件调用clusterProfiler、org.Hs.eg.db、enrichplot、ggplot2 软件包进行功能富集和通路分析,对超过8 个突变样本的基因进行基因本体论(gene ontology,GO)和京都基因和基因组百科全书 (kyoto gene and genome encyclopedia,KEGG)富集分析,完成筛选分子功能、生物过程、细胞组分和KEGG 通路进行突变基因富集。
1.4 PPI 蛋白互作网络的构建 生物网络的构建可以以实际系统规模形式进行扩展, 并提供分子相互作用的可视化表征。 使用STRING 在线数据库11.5 来描述突变基因的PPI 网络,并设置置信评分0.4 作为参考标准, 然后使用Cytoscape 软件可视化生成的PPI 网络。
1.5 基因突变与表达之间的相关性分析 探索突变与基因表达之间的相关性以分析突变在肾透明细胞癌发生中的作用。 将转录组数据标准化后与SNP 数据进行整合,分析基因突变与基因表达的联系, 采用Wilcox 秩和检验分析野生型与突变型基因表达的差异,以P<0.05 为差异有统计学意义。
1.6 基因突变的生存分析 从TCGA 下载原始临床数据中提取每个样本生存时间与生存状态信息, 使用hash 软件包将基因突变信息与生存数据进行合并, 利用survival 软件包绘制突变基因的Kaplan-Meier 生存曲线, 评估突变基因对肾透明细胞癌患者预后的影响。
2 结果
2.1 肾透明细胞癌SNP 数据分析 结果发现,106个基因超过8 例样本中发生了突变(图1)。 基于R软件相关LIMMA 软件包对肾透明细胞癌与正常组织样本进行表达差异分析, 设置|log FC|> 3, P<0.01,获得1366 个差异表达基因(图2)。
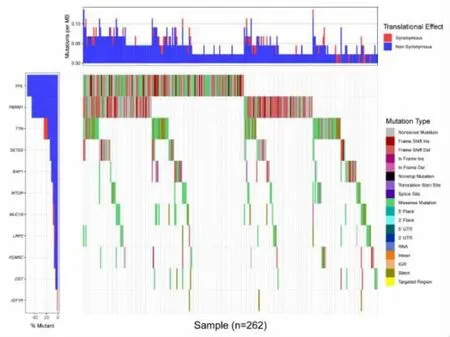
图1 肾透明细胞癌的突变基因瀑布图
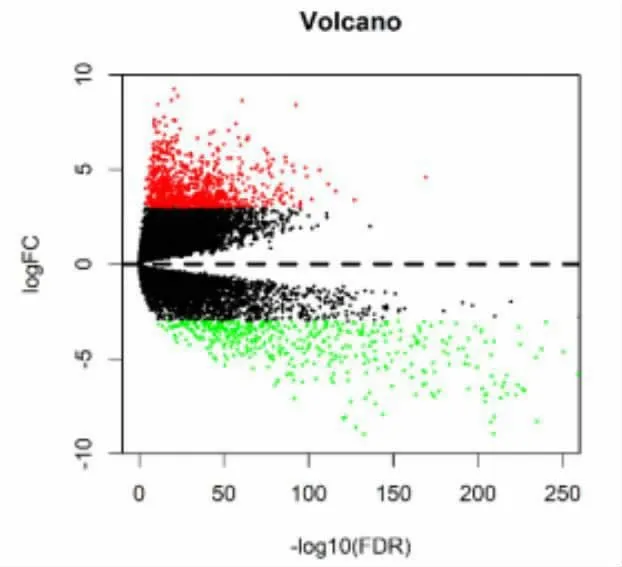
图2 肾透明细胞癌的差异基因火山图
2.2 突变基因的功能富集和通路分析 功能分析显示,在生物过程(Biological Process,BP)中,SNP 突变基因主要富集于肌肉组织的发育; 在分子功能(Molecular Function,MF)中,这些基因主要影响细胞外基质结构成分、ATP 酶活性和肌动蛋白纤维的结合。 在细胞组分(Cellular Component,CC)中,SNP 基因主要定位于细胞质和细胞外胶原纤维(图3)。此外,KEGG 通路富集分析显示,SNP 突变基因在许多癌症信号通路中富集,包括肌醇磷脂-3-激酶(PI3K)-Akt、焦点粘连、细胞外基质受体相互作用等信号通路(图4)。
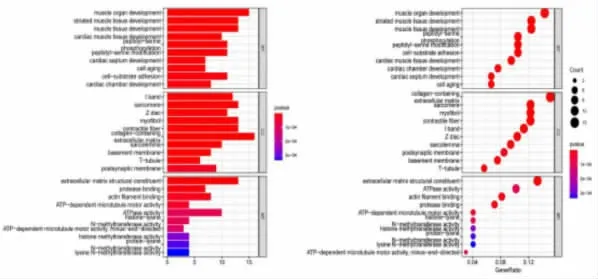
图3 肾透明细胞癌突变基因的GO 功能富集分析
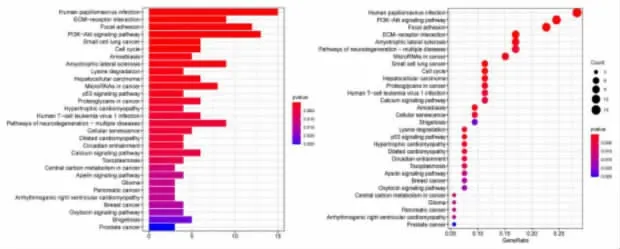
图4 肾透明细胞癌突变基因的KEGG 信号通路富集分析
2.3 突变基因PPI 网络的构建 使用Cytoscape 软件进行可视化, 构建的复杂PPI 网络, 并使用MCODE 功能筛选出重要子模块(图5)。
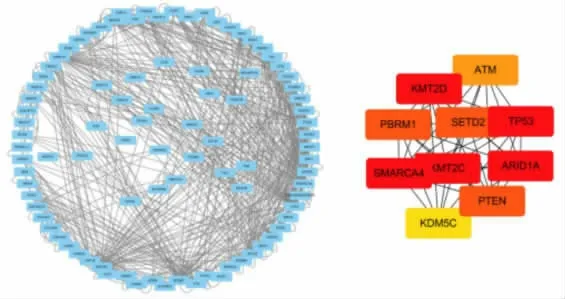
图5 肾透明细胞癌突变基因的PPI 网络及重要子模块
2.4 基因突变与表达之间的相关性分析 将基因突变与经标准化后基因表达进行整合, 提取在肾透明细胞癌中突变频率最高的5 个基因VHL、PBRM1、TTN、SETD2、BAP1 与IGF1R 的突变及表达数据, 采用wilcox 检验分析野生型与突变型基因表达的差异。 结果显示,PBRM1、SETD2、BAP1这3 个基因的突变与表达之间存在相关性 (P<0.001),而突变体样本中VHL、TTN、IGF1R 的表达量差异无统计学意义(图6)。
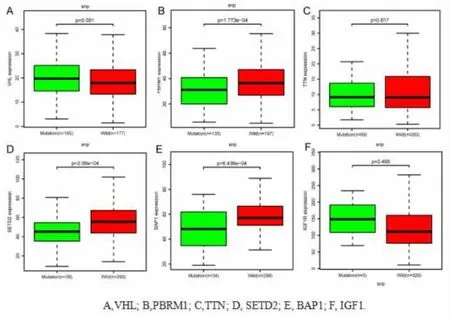
图6 肾透明细胞癌基因突变与表达的相关性分析
2.5 突变基因的生存分析 根据突变将患者分为野生型组和突变型组, 绘制VHL、PBRM1、TTN、SETD2、BAP1 与IGF1R 这6 个突变基因的Kaplan-Meier 曲线。 使用P<0.05 为显著水平,发现这6 个基的突变与患者的预后差异无统计学意义(图7)。
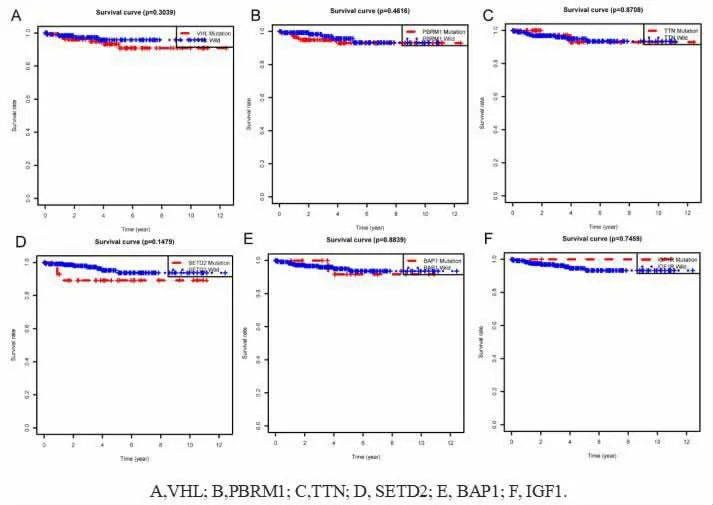
图7 肾透明细胞癌突变基因的生存曲线
3 讨论
肾透明细胞癌是一种复杂的疾病, 是肾癌相关死亡的常见原因。 SNP 与多种癌症相关,如miRNA-146a 与胃癌化疗疗效有关等,故及早发现SNP 突变,对改善患者预后有重要意义[8]。 因此,生物信息学分析突变基因的预后和特定的SNP 基因突变筛查可以为临床医生提供治疗病人和预测预后的新工具。
在本研究中, 为进一步研究突变基因参与的相关分子机制,从而进行功能富集和通路分析,结果显示该类突变基因主要参与细胞外基质结构、肌肉组织发育。 通路富集分析表明,SNP 突变基因主要参与PI3K-Akt、焦点粘连、细胞外基质受体相互作用等在许多癌症中富集的信号通路。 功能富集和通路分析揭示了SNP 突变在疾病进展中的分子机制,以及这些基因在功能水平上的相互作用。对在肾透明细胞癌患者中突变频率最高的5 个基因(VHL、PBRM1、TTN、SETD2、BAP1) 与IGF1R 基因的突变与相应的表达水平进行分析,结果表明,PBRM1、SETD2、BAP1 这3 个 基 因 的 突 变 与 表 达水平之间显著相关, 且突变后的mRNA 表达水平升高, 而突变样本中另外3 个基因VHL、TTN、IGF1R 的表达量无统计学差异。通过绘制该类突变基因的Kaplan-Meier 曲线,并进行生存分析,发现该类基因突变与患者预后无相关性。 这可能与所选样本大多来源于预后较好的早期局部肾透明细胞癌患者,导致生存时间差异不显著。
VHL 蛋白在调节缺氧诱导因子(HIF)的蛋白酶体降解中起关键作用。 VHL 失活不能介导HIF蛋白降解,从而激活这些下游生长因子转录,导致血管生成和细胞增殖增强, 促进VHL 相关疾病中肿瘤发生[9]。 Sato 等[10]研究显示VHL 基因内突变,涉及VHL 染色体缺失,VHL 双等位基因失活是散发性肾透明细胞癌发生的重要机制, 该研究还发现在KIRC 中反复突变的通路包括PI3K-AKTmTOR 信号传导。 而PI3K/AKT 是细胞中重要信号转导通路,与恶性肿瘤转移显著相关,AKT 是PI3K下游直接靶蛋白,AKT 蛋白的激活在癌症发展中具有重要生物学作用。 本研究结果显示,PI3KAKT 信号通路及其他几个与癌症密切相关的信号通路中富集, 如癌症中的焦点粘连和细胞外基质受体相互作用。
VHL 突变并不是肾透明细胞癌肿瘤发生的唯一驱动力,PBRM1、BAP1、SETD2 等染色质修饰基因也发生高频突变[11]。 PBRM1、SETD2 和BAP1 与VHL 类似, 这表明肾透明细胞癌发病机制的重要组成部分可能是3p 等位基因缺失导致四种肿瘤抑制因子同时发生失活。 而且BAP1 和SETD2 的突变可能是VHL 或PBRM1 丢失的继发事件[12]。Hakimi 等[13]研究发现,PBRM1、BAP1 或SETD2 中任何一个突变的肿瘤更可能出现III 期或更高分期的疾病,具有PBRM1 突变的小肿瘤(<4 cm)更有可能表现出III 期病理特征,BAP1 突变往往发生在Fuhrman III-IV 级肿瘤中, 并且与更差的肿瘤生存时间相关。 因此这些染色质修饰基因发生突变, 可能与晚期、 高病理分期和更差的预后相关。
胰岛素样生长因子1 受体 (IGF1R) 基因位于15q26。 IGF1R 蛋白调节众多下游信号,通常通过调节肿瘤细胞生长和凋亡参与肿瘤发生、 存活和转移[14-15]。IGF1R 包括2 个配体(IGF-1 和IGF-2),两个细胞表面受体(IGF-1R 和IGF-2R)和至少六个IGF 结合蛋白(IGFBP 1-6),控制正常组织生长与大多数器官分化。 其中1%循环IGF-1 蛋白与IGF-1R 结合触发两个信号,即刺激增殖和抗细胞凋亡。 IGF-1 可刺激细胞DNA 合成,上调细胞周期蛋白D1 表达, 促使细胞由G1 期进入S 期,同时通过调节Bcl 和Bax 蛋白,抑制细胞凋亡。 因此IGF-1 抗凋亡作用可使有恶性变倾向的细胞逃避凋亡而促使肿瘤发生。在本研究中,发现IGF1R 在突变样本中表达水平降低,差异无统计学意义,这可能与该基因突变样本比例较少有关。 此外,本研究结果未得到突变与不良预后的有效结果, 这需要进一步的临床研究及测序分析。
综上所述, 本研究通过生物信息学分析为临床诊断和预后评估提供新的思路, 为肾透明细胞癌的后续研究提供了重要的生物信息学基础和相关理论依据。
猜你喜欢
中国现代医生(2022年19期)2022-11-04
中国医学物理学杂志(2022年9期)2022-10-09
中国现代医生(2022年19期)2022-08-25
中国中医药信息杂志(2022年1期)2022-01-13
中国听力语言康复科学杂志(2021年6期)2021-12-21
科学之谜(2018年2期)2018-03-14
中学生理科应试(2017年6期)2017-09-27
销售与管理(2006年9期)2006-09-17
中国美容医学(2004年3期)2004-09-17
中国美容医学(2004年3期)2004-09-17
