
On-off-on fluorescence detection for biomolecules by a fluorescent cage through host-guest complexation in water
2022-06-20 07:59HonghongDunFnCoMinjieZhngMengGoLipingCo
Chinese Chemical Letters 2022年5期
Honghong Dun, Fn Co, Minjie Zhng, Meng Go, Liping Co,*
a College of Chemistry and Materials Science, Northwest University, Xi’an 710069, China
b National Engineering Research Center for Tissue Restoration and Reconstruction, Key Laboratory of Biomedical Engineering of Guangdong Province, Key Laboratory of Biomedical Materials and Engineering of the Ministry of Education, Innovation Center for Tissue Restoration and Reconstruction, South China University of Technology, Guangzhou 510006, China
Keywords:Host-guest complexation Tetraphenylethene-based octacationic cage Nucleoside derivatives Fluorescence Cell imaging
ABSTRACT Detection of nucleoside derivatives has paramount importance because they are the essential biomolecular units for all life.Herein, we report a host-guest approach by using a fluorescent tetraphenylethenebased octacationic cage as host and 8-hydroxypyrene-1,3,6-trisulfonic acid trisodium salt (HPTS) as guest and fluorescent indicator to form non-fluorescent 1:1:1 host-(endo-exo)guest complex in water.This new host-(endo-exo)guest complex can be successfully used for detecting nucleosides (e.g., ATP and GTP), DNA(e.g., sm-DNA), and antibiotics (e.g., Penicillin G) with off-on fluorescence response via a competitive hostguest exchange with HPTS as exo-guest in water.Furthermore, this on-off-on fluorescent host-guest complex is also used for cell imaging based on ATP concentration in HeLa cells.Therefore, this study not only provides insight into the construction of a supramolecular probe with on-off-on fluorescence via host-guest complexation and exchange in solution, but also realizes a universal method for detecting and monitoring biomolecules.
The research progress of fluorescence detection systems for biomolecules has received considerable attention in recent years because of the important application in a wide range of fields such as biology, chemical process, and environmental science [1–5].Specifically, nucleoside derivatives, such as ATP, CTP, GTP and UTP, are essential in the regulation of organisms [6–10].They are not only the raw materials for RNA synthesis, but also play an important role in metabolism process.For example, ATP is the "electricity" in organisms; GTP is essential in protein synthesis; UTP is involved in carbohydrate metabolism in organisms; CTP plays an important role in phospholipid synthesis [11,12].However, owing to the solvation of phosphate and sugar groups and the competitive hydrogen bonding of base units with water molecules, their detection and recognition in the aqueous medium still are limited [13–15].On the other hand, most of conventional fluorescent probes exhibit aggregation-caused quenching (ACQ) in water when aggregating and/or binding with biomolecules, which often makes the detection process to employ a fluorescent “turn-off” feature with the high background noise and poor sensitivity [16].Therefore, the use of fluorescent turn-on mechanism can provide a better approach for the detection of biomolecules.
In host-guest chemistry, a variety of macrocyclic hosts, such as crown ethers [17,18], cyclodextrins [19,20], calix[n]arenes [21,22],pillar[n]arenes [23,24], cyclophanes [25,26], cucurbit[n]urils[27,28], and molecular cages [29–36], have been synthesized and their functions have been widely investigated for molecular recognition, drug delivery systems, photoelectric materials, and supramolecular polymers.Especially, molecular cages with various sizes, shapes, and functional groups have been widely developed,because of not only their unique hydrophobic cavity for selectively binding guest molecules but also their reduction-oxidation properties for electronic devices (e.g., semiconductor) and molecular machines (e.g., molecular shuttle and molecular pump) [37–39].Furthermore, molecular cages have also attracted intensive attention for biochemical applications including bioimaging and drug/gene delivery [40–44], photodynamic therapy [45,46], and biosensors [47].
Recently, we have utilized tetraphenylethene (TPE) and pyridinium units as molecular building blocks to construct a series of supramolecular hosts including macrocycles, cages, and frameworks, which exhibited promising applications in host-guest recognitions, light-harvesting systems, and stimuli-responsive fluorescent materials [48–59].Specially, combined with the physicochemical properties of TPE and pyridinium units, the tetraphenylethenebased octacationic cage (1·8Cl-, Scheme 1) possesses a large and rigid binding cavity, positive-charged surfaces, good watersolubility, and excellent fluorescence [52].This fluorescent cationic cage exhibits remarkable affinities toward negative-charged chiral biomoleculesviaa series of CH-π, π-π, hydrophobic, and electrostatic interactions in water to achieve conformationally adaptive chirality with circular dichroism (CD) and circularly polarized luminescence (CPL) responses [57].As a result, this cage is endowed with several features, such as host-guest ability, strong fluorescence, and good water-solubility, which make it well suited for studying stimuli-responsive fluorescence processes through forming host-guest complexes with organic dyes in aqueous media.Two water-soluble pyrene-based dyes, 8-hydroxypyrene-1,3,6-trisulfonic acid trisodium salt (HPTS) and 1,3,6,8-pyrenetetrasulfonic acid tetrasodium salt (PTSA), are selected as guest to form fluorescent host-guest complexes with 1·8Cl-in water.Herein, we report a host-guest approach by using a tetraphenylethene-based octacationic cage as host and HPTS as guest to form non-fluorescent 1:1:1 host-(endo-exo)guest complexes, which is successfully used for detecting nucleosides (e.g., ATP and GTP), DNA (e.g., sm-DNA),and antibiotics (e.g., Penicillin G) with turn-on fluorescence responseviaa competitive host-guest exchange withexo-HPTS in water.Furthermore, this on-off-on fluorescent host-guest complex also is used for cell imaging based on ATP concentration in HeLa cells.
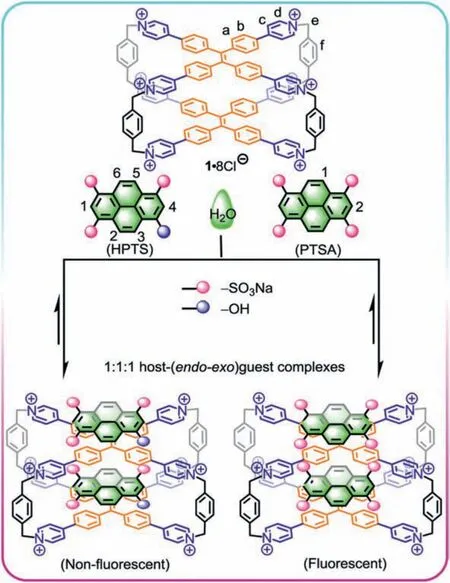
Scheme 1.The formation of host-(endo-exo)guest complexes between 1·8Cl- and HPTS/PTSA.
Initially, the host-guest complexation between 1·8Cl-and HPTS was investigated by employing NMR experiments.The1H NMR titration of 1·8Cl-with HPTS showed the following changes for the proton resonances of both cage and guest (Fig.1a):1) With the gradual addition of HPTS, the proton resonances (Hd-Hc) of pyridinium rings showed downfield shifts while the proton resonances (Ha-Hb) in the central part of the TPE faces displayed different upfield shifts which caused byπ-electron deshielding and shielding of HPTS.2) The proton resonances (H1-H6) corresponding to HPTS shifted upfield, strongly confirming that HPTS molecules are completely encapsulated inside the cavity of cage to form the host-guest inclusion complex.3) The addition of 2.0 equiv.of HPTS induced that all resonances (Ha-Hf) of cage split into two sets of peaks (Ha’-Hf’and Ha’’-Hf’’), because the formation of a new host-(endo-exo)guest complex decreases the symmetry of cage.Based on the results from NMR, the cavity of cage cannot encapsulate two HPTS.Therefore, one HPTS molecule asendoguest is located inside the cavity of cagevia π-π, hydrophobic, and electrostatic interactions, and another HPTS molecule asexo-guest binds on the tetrapyridinium-TPE surface to give a 1:1:1 host-(endo-exo)guest complex (1⊃endo-HPTS)·exo-HPTSviaelectrostatic interactions between the positive-charged surface of cage and the negative-charged sulfonic groups of HPTS.
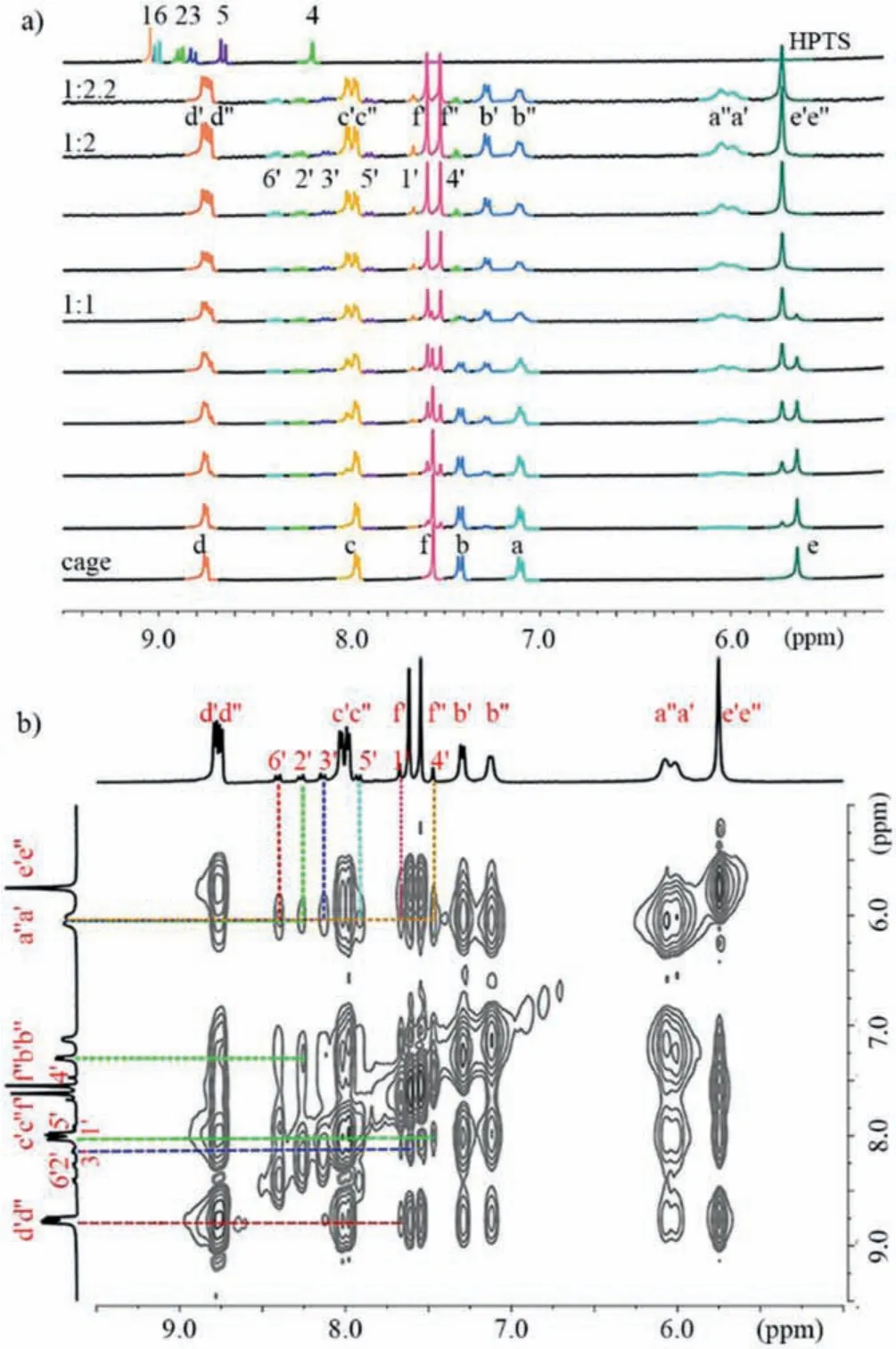
Fig.1.(a) 1H NMR titration (400 MHz, 298 K, D2O) of 1·8Cl- (0.4 mmol/L) titrated with HPTS (0-2.2 equiv.).Here, primes ’and ’’denote the resonances of the inner and outer tetrapyridinium-TPE surfaces within the 1:1:1 host-(endo-exo)guest complex, respectively.(b) NOESY spectrum recorded (400 MHz, 298 K, D2O) for 1·8Cl-(2.0 mmol/L) with HPTS (2.0 equiv.).
Furthermore,1H-1H correlation spectroscopy (COSY) spectra show strong signal peaks between Ha’and Hb’, and Ha’and Hb’’,which indicates that the symmetry of the cage is broken and the 1:1:1 host-(endo-exo)guest complex is formed (Fig.S1 in Supporting information).Besides, nuclear overhauser effect spectroscopy(NOESY) spectra showed the correlation signals between proton H1’-H6’ of HPTS and proton Ha’-Hd’ on the TPE units of cage(Fig.1b), indicating that HPTS and cage form host-guest complex.The single host-guest complex was confirmed by diffusionordered spectroscopy (DOSY), which showed a similar diffusion coefficient [(1.79 ± 0.03) × 10-10m2/s] when compared with that of 1·8Cl-[(1.95 ± 0.05) × 10-10m2/s] in D2O at 298 K (2.0 mmol/L),thus suggesting the formation of a host-guest complex (Figs.S2 and S3 in Supporting information).On the other hand, electrospray ionization time-of-flight mass spectrometry (ESI-TOF-MS)provided further evidence for 1·8Cl-:HPTS = 1:2 (including inner and outer guests) with continuous charge states atm/z663.6344,896.5014, and 877.1946 corresponding to +4 to +3 charge states due to successive loss of the Cl-and Na+counterions.Meanwhile, 1·8Cl-:HPTS = 1:1 with continuous charge states atm/z430.5416, 547.1665, 741.2040 and 1129.7878 corresponding to +5 to +2 charge states due to successive loss of the Cl-and Na+counterions (Figs.S4 and S5 in Supporting information).Owing to the weak binding between cage andexo-HPTS, only one binding constant between 1·8Cl-andendo-HPTS was estimated as (1.85 ±0.13) × 105L/mol by UV-titration experiment (Fig.S6 in Supporting information).
Next, the host-guest behavior of 1·8Cl-and PTSA was investigated in D2O.In1H NMR titration of 1·8Cl-with PTSA, downfield shifts were observed for pyridinium protons (Hc’-Hd’), bridging CH2groups (He’), andp-xylylene moieties (Hf’), while the phenyl proton resonances (Ha’-Hb’) located in the central part of the cage’s TPE units showed apparent upfield shifts as compared to the free cage at 1:1 ratio, which caused byπ-electron deshielding and shielding of PTSA (Fig.S7 in Supporting information).Meanwhile, proton resonances (H1-H2) showed obvious upfield shifts and was split indicating that PTSA was entirely located within the cage and shielded by the cavity of 1·8Cl-.Furthermore, NOE correlation signals were observed between proton H1’-H2’of PTSA and proton Ha’-Hb’on the TPE of cage from the 2D NOESY NMR spectrum.These results further confirm the central location of PTSA in the cage cavity (Fig.S8 in Supporting information).However, with the gradual addition of PTSA, a lot of precipitate began to appear, and when 2.0 equiv.of PTSA was added, host-guest complex was completely precipitated, resulting in the complete disappearance of NMR signal.We speculated that two PTSA with eight negative sulfonic groups and octacationic cage form a poor water-soluble zwitterionic complex (1⊃endo-PTSA)·exo-PTSA through multiple electrostatic interactions.ESI-TOF-MS further provided evidence for the formation of 1⊃endo-PTSA with continuous charge states atm/z553.9076,750.5335, corresponding to +4 to +3 charge states due to successive loss of the Cl-and Na+counterions (Fig.S9 in Supporting information).The binding constants between 1·8Cl-and PTSA was calculated as (8.01 ± 0.09) × 105L/mol for 1⊃endo-PTSA (Fig.S10 in Supporting information).
Subsequently, the photophysical properties of 1·8Cl-with HPTS/PTSA were analyzed by fluorescence and UV-vis experiments in water.As shown in Fig.2a, when HPTS was successively added into the solution of cage in water, the absorbance at 284 nm increased and red-shifted to 290 nm, while the absorbance at 362 nm decreased and red-shifted to 369 nm, indicating the formation of the host-guest complex.At the same time, the absorption peak from 440 nm gradually appears, which may be caused by a strong intermolecular charge-transfer interaction between anionic HPTS and octacationic pyridinium units of cage.A dramatic fluorescence quenching (ΦF= from 0.27 to<0.01) of cage at 540 nm when 2.0 equiv.of HPTS was added, which could be attributed to the strong photoinduced electron transfer (PET) from HPTS to cage based on the formation of the host-guest complex (Fig.2b and Table S1 in Supporting information) [60–62].On the other hand, when PTSA was successively added to cage in water, UV-vis spectroscopy showed that there was weak charge transfer interaction between 1·8Cl-and PTSA (Fig.2c).Therefore, compared with HPTS, the fluorescence spectra of PTSA showed different changes.With the gradual addition of PTSA, the fluorescence intensity of cage at 540 nm gradually increased and reached the maximum at 1:1 ratio, indicating that a 1:1 host-guest complex 1⊃endo-PTSA was formed,which is consistent with the result from NMR titration.The fluorescent enhancement could be contributed to the restriction of intramolecular rotation (RIR) of TPE units of cage when forming the host-guest complex.After the continuous addition of PTSA, the fluorescence intensity of cage showed red shift (Δλmax= 23 nm) and slightly decreased (Fig.2d), accompanying with a precipitation, indicating that a zwitterionic complex (1⊃endo-PTSA)·exo-PTSA was formed.The 1931 CIE chromaticity diagram also confirmed tracks of the fluorescence color change, followed by the titration of HPTS and PTSA (Fig.S11 in Supporting information).
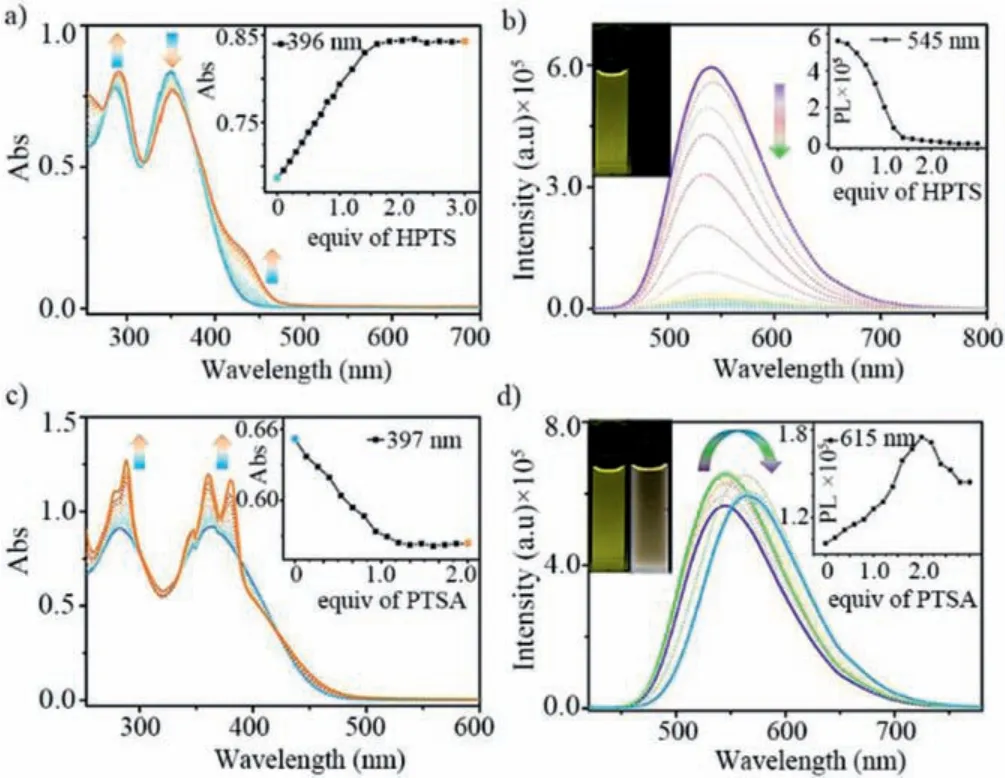
Fig.2.(a) UV-vis and (b) fluorescence spectra of 1·8Cl- titrated with HPTS; (c) UVvis and (d) fluorescence spectra of 1·8Cl- titrated with PTSA.[1·8Cl-] = 10 μmol/L,λex = 410 nm, Ex/Em slit = 1.2 nm, solvent:H2O.
Given the on-off fluorescence of octacationic cage with HPTSviahost-(endo-exo)guest complexation, a series of negative-charged nucleoside derivatives (e.g., ATP, AMP, ADP, CTP, GTP and UTP,Scheme S1 in Supporting information) were selected as competitive guests for releasing theexo-HPTS from host-(endo-exo)guest complex (1⊃endo-HPTS)·exo-HPTS in aqueous solution.The competitive fluorescence experiments confirmed that the off-on fluorescence process can be realized by a competitive host-guest exchange, resulting different turn-on fluorescence responses for different nucleosides (Scheme 2).In fluorescence titration experiment(Fig.3a), (1⊃endo-HPTS)·exo-HPTS exhibited various off-on fluorescence responses to nucleosides with different intensities, absolute quantum yields, and lifetimes (Table S1).Interestingly, the fluorescence titrations of (1⊃endo-HPTS)·exo-HPTS with ATP, GTP or UTP in water showed a drastic increase in the intensity of the emission of HPTS centered at 515 nm with high fluorescence intensity ratio(I/I0) of ~31, ~71 and ~25, respectively, exhibiting excellent green fluorescence emission of HPTS (Figs.3a and b).However, other nucleosides, such AMP, ADP, and CTP, just caused low enhancement in fluorescence intensity (Fig.3b and Figs.S12-S17 in Supporting information).These results indicated that the electrostatic interaction between positive-charged pyridinium units of host and negativecharged phosphate groups of guests is main driving force to release theexo-HPTS from (1⊃endo-HPTS)·exo-HPTSviacompetitive hostguest exchange between theexo-HPTS of the host-guest complex and negative-charged nucleoside derivatives.For nucleoside derivatives with different nucleobase and the same number of phosphate groups, such as ATP, UTP, CTP and GTP, the fluorescence intensities of (1⊃endo-HPTS)·exo-HPTS were also different in the titration processes, which indicated that the nucleobase could also affect the binding affinity with 1⊃endo-HPTS through hydrophobic effect andπ-πinteraction.
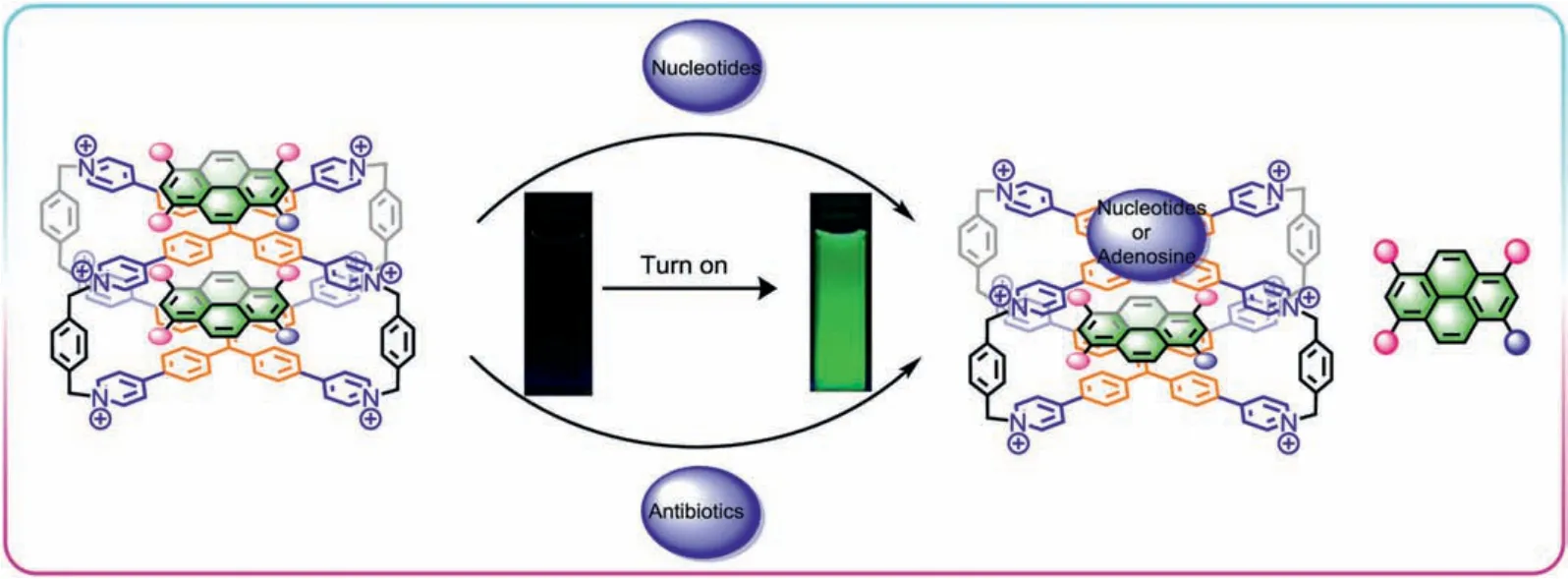
Scheme 2.Schematic illustration of the process of turn-on fluorescence via competitive exclusion host-guest exchange.
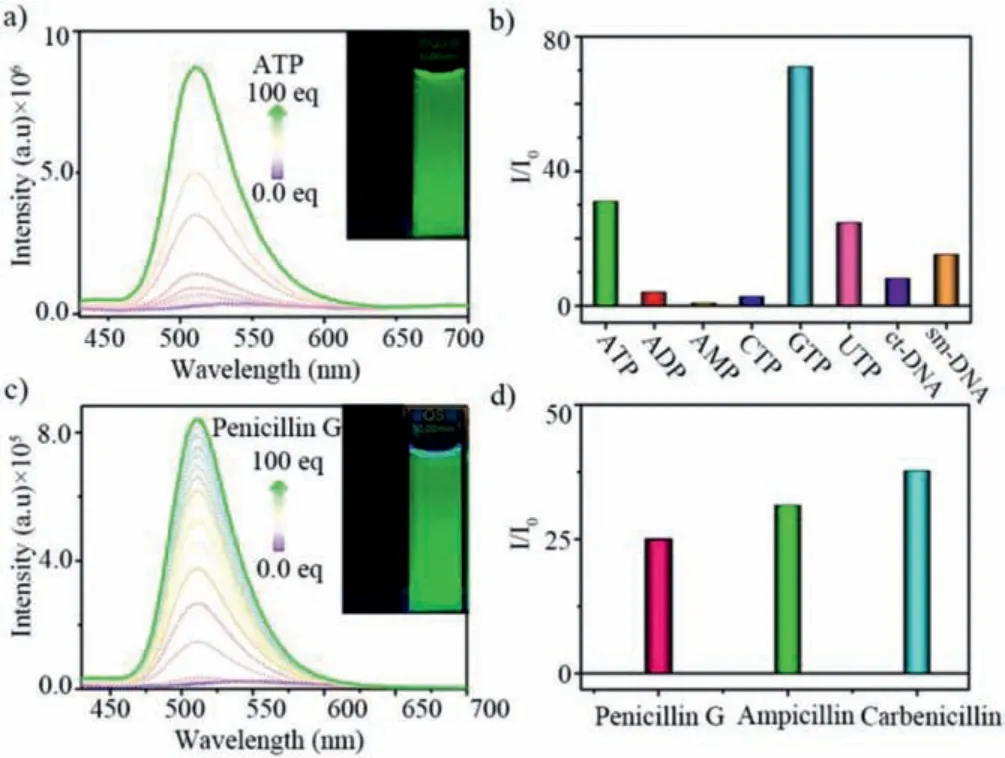
Fig.3.Fuorescence spectra of (1⊃endo-HPTS)·exo-HPTS (10 μmol/L) titrated with(a) ATP (0-100.0 equiv.), (c) Penicillin G (0-100.0 equiv.).The I/I0 of (1⊃endo-HPTS)·exo-HPTS with (b) nucleosides and DNA (5.0 equiv.), (d) antibiotics (75.0 equiv.).
To further verify the mechanism of turn-on fluorescence, NMR titration experiments were carried out in D2O.When ATP was successively added to (1⊃endo-HPTS)·exo-HPTS in D2O, the split proton peaks of 1 in (1⊃endo-HPTS)·exo-HPTS became broaden and converged again, indicating that theexo-HPTS on the outer surface was released after the addition of ATP (Fig.S18 in Supporting information).Theendo-HPTS in the cavity still formed a stable hostguest inclusion complex with cage (Scheme 2).This experimental result is consistent with the fluorescence experiment.
Given the off-on fluorescence responses between (1⊃endo-HPTS)·exo-HPTS and nucleoside derivatives, (1⊃endo-HPTS)·exo-HPTS can be an ideal probe for DNA molecules.As expected,the addition of salmon testes DNA (smDNA) or calf thymus DNA(ctDNA) into the solution of (1⊃endo-HPTS)·exo-HPTS in water induced drastic fluorescence enhancement with ~8 and ~15 times,respectively (Fig.3b and Figs.S19 and S20 in Supporting information).The detection limits of (1⊃endo-HPTS)·exo-HPTS were calculated to be 2.03 μg/μL for smDNA and 1.53 μg/μL for ctDNA, respectively (Figs.S19 and S20 in Supporting information).
With the abuse of antibiotics, the detection of antibiotics has become a hot spot for scientists (Scheme S1 in Supporting information).Due to the host-guest complex can detect negativecharged molecules with turn-on fluorescence response in water,(1⊃endo-HPTS)·exo-HPTS can be an ideal probe for antibiotics, such as Penicillin G, ampicillin and carbenicillin.Not surprisingly, with the addition of antibiotics, the emission intensity of (1⊃endo-HPTS)·exo-HPTS at 515 nm was significantly enhanced, with ~25,~31 and ~38 times (Figs.3c and d Figs.S21-S23 in Supporting information), respectively.The off-on fluorescence response was successfully realized.These experimental results show that the host-(endo-exo)guest complex is universal and can be used as a probe to detect various substances in water.
The host-(endo-exo)guest complex (1⊃endo-HPTS)·exo-HPTS also was employed for cell imaging.Confocal laser scanning microscopy (CLSM) experiments in HeLa cells were performed to assess whether host-guest probe (1⊃endo-HPTS)·exo-HPTS could be in mitochondrion selectively, where the concentration of ATP is higher than other area in cells (Fig.4).HeLa cells stained with Mito-Tracker Deep Red (a commercially mitochondrial tracker) displayed fluorescence on the red channel (Fig.4a).The cells stained with (1⊃endo-HPTS)·exo-HPTS exhibited fluorescence on the green channel in Fig.4b.The bright-field (Fig.4c) and the merged(Fig.4d) images of the cells showed overlap with commercial mitochondrial dyes, indicating that (1⊃endo-HPTS)·exo-HPTS can serve as a mitochondrion fluorescent probe.CLSM imaging and fluorescence intensity profile confirm that the emission is belonged to free HPTS (Figs.4e and f).

Fig.4.CLSM imaging of HeLa cells stained with Mito-Tracker Deep Red (100 μmol/L) and (1⊃endo-HPTS)·exo-HPTS (50 μmol/L):(a) red channel, (b) green channel, (c) bright field, and (d) merged image.(e) CLSM imaging and (f) fluorescence intensity profile of regions of interest.
In conclusion, we have reported the formation of 1:1:1 host-(endo-exo)guest complexes between TPE-based octacationic cage with organic dyes (e.g., HPTS and PSTA) in water.In these host-(endo-exo)guest complexes, cage can entirely encapsulate one dye asendo-guest inside the hydrophobic cavity and bind with another dye asexo-guest on the outer face throughπ-π, hydrophobic, and electrostatic interactions.Their host-guest behaviours have been investigated by1H NMR, UV-vis, fluorescence experiments.In aqueous media, cage exhibits an on-off fluorescence quenching when binding with HPTS.We utilize this on-off fluorescent hostguest system as a fluorescent probe to selectively detect negativecharged biomolecules including nucleoside derivatives, DNA, and antibiotics to achieve an off-on fluorescence response.We anticipate that this design of on-off-on fluorescent host-guest system has universality for probing biomolecules and cell imaging in water.
Declaration of competing interest
The authors declare no conflict of interest.
Acknowledgments
This work was supported by the National Natural Science Foundation of China (Nos.22122108, 21971208 and 21771145), the Natural Science Basic Research Plan for Distinguished Young Scholars in Shaanxi Province of China (No.2021JC-37), and the Fok Ying Tong Education Foundation (No.171010).
Supplementary materials
Supplementary material associated with this article can be found, in the online version, at doi:10.1016/j.cclet.2021.11.010.
 Chinese Chemical Letters2022年5期
Chinese Chemical Letters2022年5期
- Chinese Chemical Letters的其它文章
- Recent advances in enhancing reactive oxygen species based chemodynamic therapy
- An integrative review on the applications of 3D printing in the field of in vitro diagnostics
- Recent developments of droplets-based microfluidics for bacterial analysis
- Dynamics and biological relevance of epigenetic N6-methyladenine DNA modification in eukaryotic cells
- Recent progress in advanced core-shell metal-based catalysts for electrochemical carbon dioxide reduction
- Recent advances in carbon-based materials for electrochemical CO2 reduction reaction