
黄檗半同胞家系苗期生长差异研究
2022-06-15 04:06郭佳忱张骁杨奇刘剑巩佳琦程广有
特产研究 2022年3期
郭佳忱,张骁※,杨奇,刘剑,巩佳琦,程广有
(1.吉林省林业勘察设计研究院,吉林 长春 130022;2.北华大学林学院,吉林 吉林 132013)
黄檗(Phellodendron amurense Rupr.)为芸香科黄檗属高大落叶乔木,是东北“三大硬阔”之一,具有广泛的应用价值[1-3]。其内皮被称为关黄柏,是传统的中草药,药用效果显著[4,5]。
黄檗材质坚韧,纹理美观,是实木家具的珍贵用材[6]。近年来对黄檗的利用也见于园林绿化行道、嫩叶茶品和美容化妆品等[7,8]。林木生长差异受林木个体基因与所生长的环境条件影响,特别在幼苗生长阶段,合理地进行栽培管理有助于苗木的健康生长[9]。目前已有学者对多个树种苗期构建了生长模型,用以探讨林木苗期生长节律。郭欢欢等[10]对黄连木1 年生实生苗的年生长节律进行模型拟合,并将苗期生长阶段分为出苗期、生长初期、速生期和生长后期。刘梦然等[11]构建了转TaLEA基因小黑杨无性系苗期生长性状的Logistic模型,结果发现苗高、地径均与速生期内的生长量和平均日生长量间呈极显著正相关。蒋燚等[12]对黑荆树苗期生长节律进行了研究,结果发现黑荆树0.5 年生幼苗的生长进程具有明显的间歇性,苗高和地径均有2次生长高峰,其生长过程可划分为生长前期、速生期和盛末期。关于黄檗苗期生长节律的研究较少,本文构建了16 个黄檗半同胞家系的苗期生长模型,对黄檗苗期生长阶段进行了划分,分析了黄檗家系间生长差异,旨在为黄檗苗期选择和栽培管理提供参考。
1 材料与方法
1.1 试验材料
黄檗16 个家系种子取自于吉林省临江林业局黄檗种子园,该园建于1999 年,树种主要来源于小兴安岭和长白山,园区海拔793 m,年平均气温1.4 ℃,年平均降水量830 mm,属温带大陆性季风气候。在种子园的黑龙江Ⅰ区和临江Ⅰ区内选择16 个家系,每个家系选3 个分株,共48 株树。
1.2 试验方法
1.2.1 苗期生长试验 黄檗苗期生长试验在吉林市磨盘山实验基地开展。该地距吉林市区15 km,属中温带亚湿润季风气候类型,年平均气温3.9℃,1 月平均气温最低,7 月平均气温最高。全年平均降水量650~750 mm。2015 年5 月上旬,随机选择每株黄檗母树种子1 000粒,均匀散播在2 m 1.2 m 的苗床上,5 月中、下旬陆续有个别黄檗家系种子发芽破土,6 月初种子全部破土后对黄檗各家系分株的苗高和地径进行调查。
1.2.2 生长调查 16 个黄檗半同胞家系子代的出苗率介于7.6%~32.9%,每个子代采用5 点法选择30 株长势良好的幼苗,利用直尺对苗高进行测量,测量苗木与地面的最大高度,精度为0.1 cm;利用游标卡尺对地径进行测量,测量苗木地面以上1 cm 处的直径,精度为0.1 mm。
1.2.3 统计分析 选择的黄檗苗期生长模型为Logistic 方程:
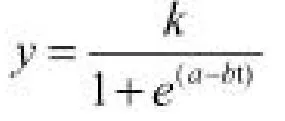
预估精度P:
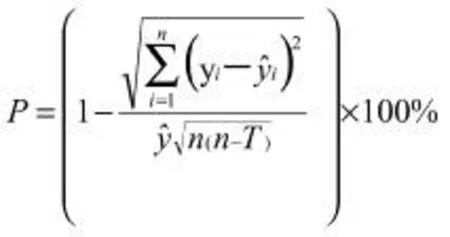
式中y 为黄檗苗高或地径,t 为生长天数,k、a、b为模型参数。n 为模型检验样本数,为16 个;T 为模型中参数个数,为3 个;yi为苗高或地径的观测值为苗高或地径的拟合值,i 为各家系分株。对模型进行求导,可得苗期生长速生点t0=a/b,速生初期t1=(a-1.317)/b,速生末期t2=(a+1.317)/b,速生持续时间t△=t2-t1。
幼苗生长日期差异性分析参考张骁等对黄檗花期差异性分析的方法[3],在本实验中,以2015 年6 月1日为第1 天(记为1),6 月8 日(记为2),以此类推。
根据黄檗苗高和地径速生初期、末期、速生点及持续时间,采用最小距离法对16 个黄檗家系进行聚类分析,确定各黄檗家系生长类型。
本文利用office2017 对数据进行整理,并利用SAS8.0 软件对黄檗半同胞家系苗期生长进行方差分析、多重比较和聚类分析。
2 结果与分析
2.1 模型参数与检验
利用Logistic 模型对黄檗16 个家系苗高和地径进行拟合,见表1。黄檗16 个家系苗高和地径参数检验均达到显著性水平,实测值与拟合值检验精度均大于0.87,说明模型对黄檗苗高和地径的拟合精度较高。其中L1-4 号家系模型拟合最优,检验精度达0.913 0;H1-1 号家系地径拟合精度最优,检验精度达0.969 0。
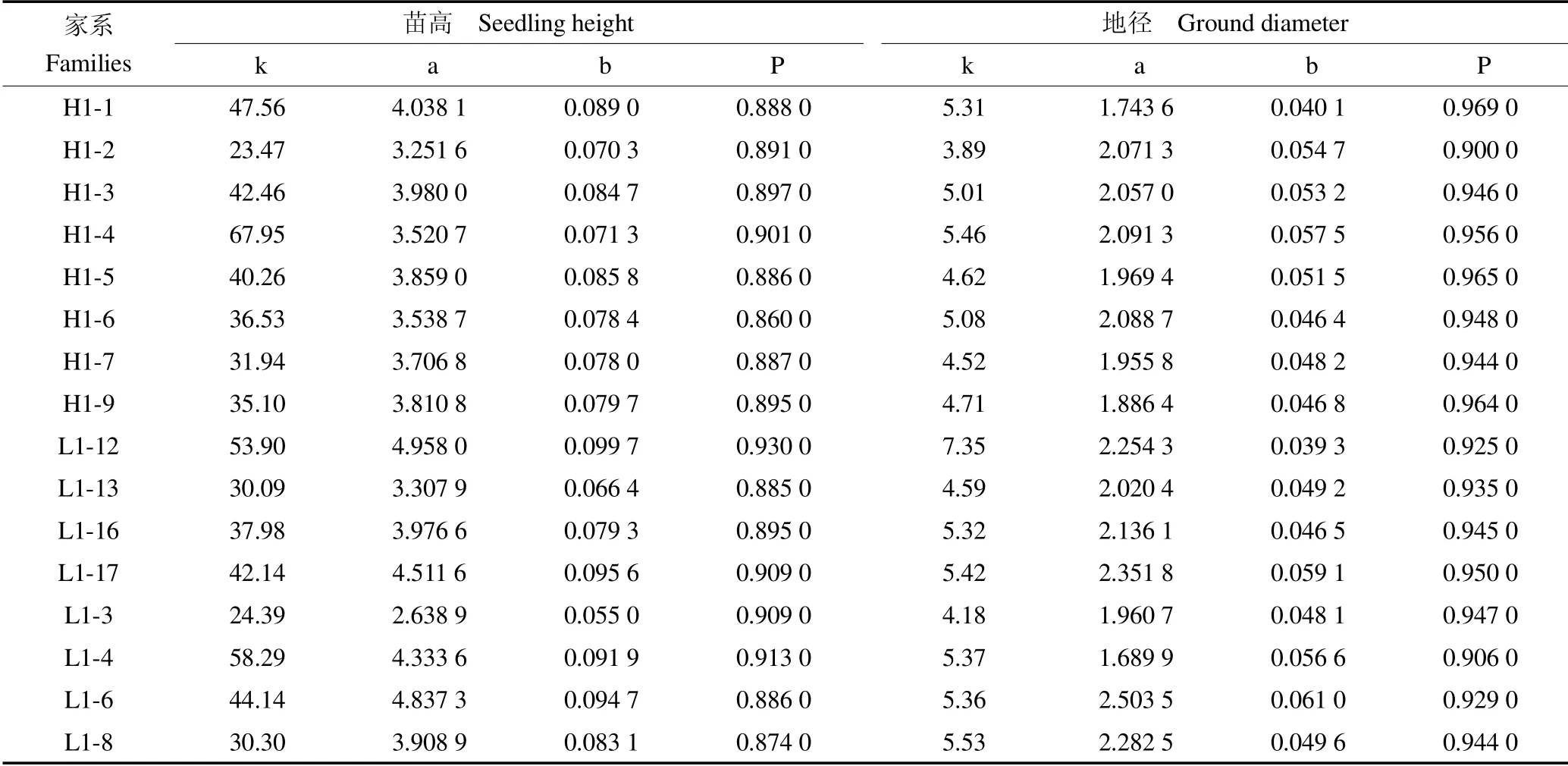
表1 16 个黄檗家系苗高和地径模型参数与检验Table 1 The model parameters and test of seedling height and ground diameter of 16 families
2.2 苗期生长差异
对黄檗家系间苗高和地径速生初期、末期、速生点及持续时间进行方差分析,结果显示,家系间苗高和地径速生初期差异均不显著(F=1.14 和F=1.01),说明遗传因素对苗木生长初期影响较小。家系间苗高速生末期和持续时间差异均达到显著水平(F=2.42*和F=3.12**);家系间地径速生末期和持续时间差异均达到显著水平(地径F=3.86**和F=2.69**)。说明随着黄檗苗木生长,遗传因素对苗期生长的影响逐渐提高。
进一步对黄檗家系间苗高和地径生长速生初期、末期及持续时间进行多重比较见表2 和表3。结果发现,L1-3 号家系苗高生长最早进入速生期,为7 月25日,仅与L1-17、L1-6、L1-16、L1-4 和L1-12 号家系差异显著;L1-13 号家系苗高生长最晚结束速生期,为9月15 日,除与L1-3 号家系差异不显著外,与其他家系差异均达到显著水平;L1-3 号家系苗高速生期持续时间较长,为49 d,除与L1-13 和H1-2 号家系差异不显著外,与其他家系差异均达到显著水平。L1-6 号家系地径生长最早进入速生期,为7 月12 日,与其他家系差异均不显著;H1-1 号家系地径生长最晚结束速生期,为9 月25 日,除与H1-6 号家系差异不显著外,与其他家系差异均达到显著水平;H1-1 号家系地径速生期持续时间最长,为75 d,与其他家系差异均达到显著水平。
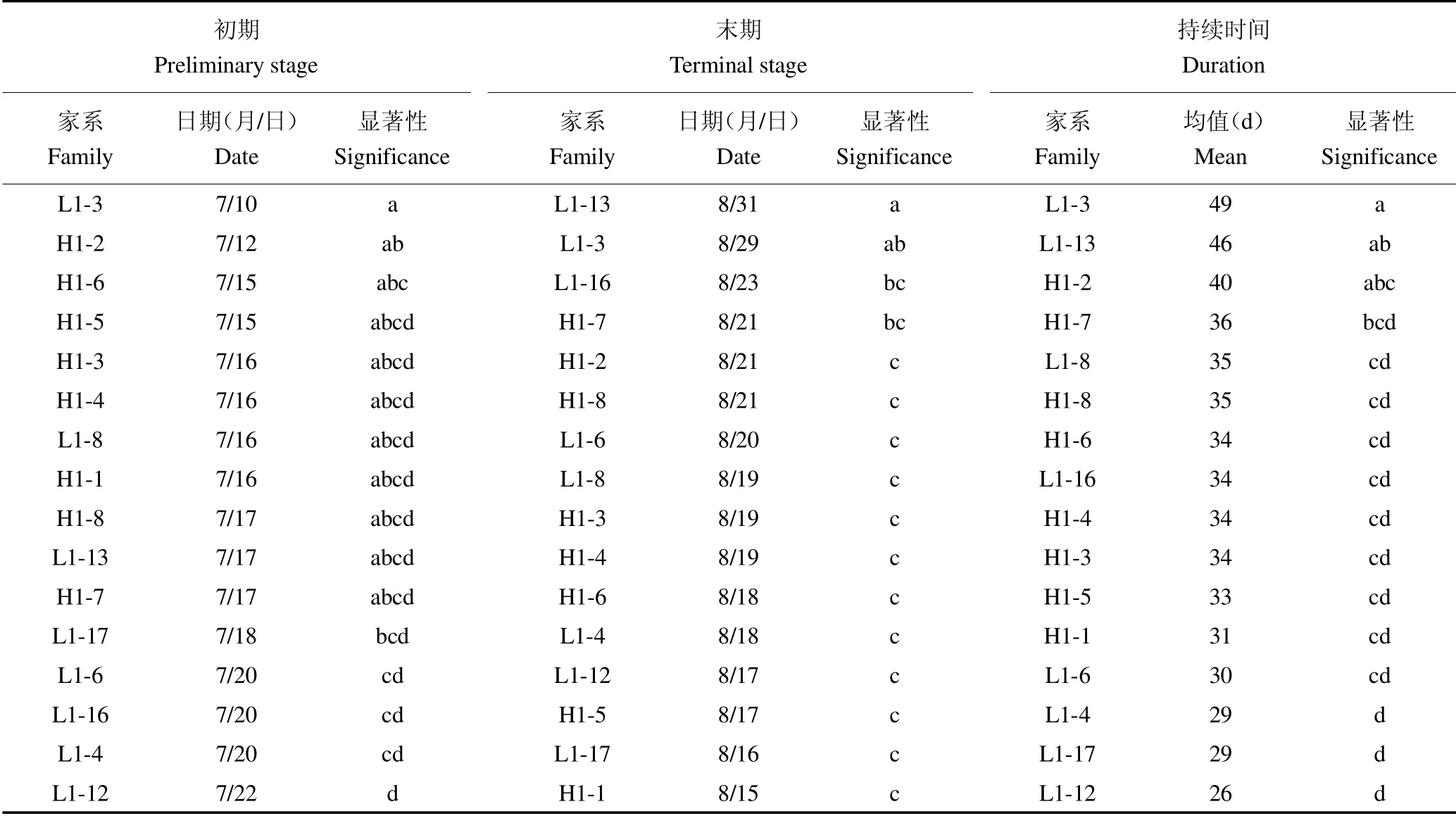
表2 家系间苗高速生末期及持续时间多重比较Table 2 Multiple comparisons of late and duration of rapid growth of seedling height families
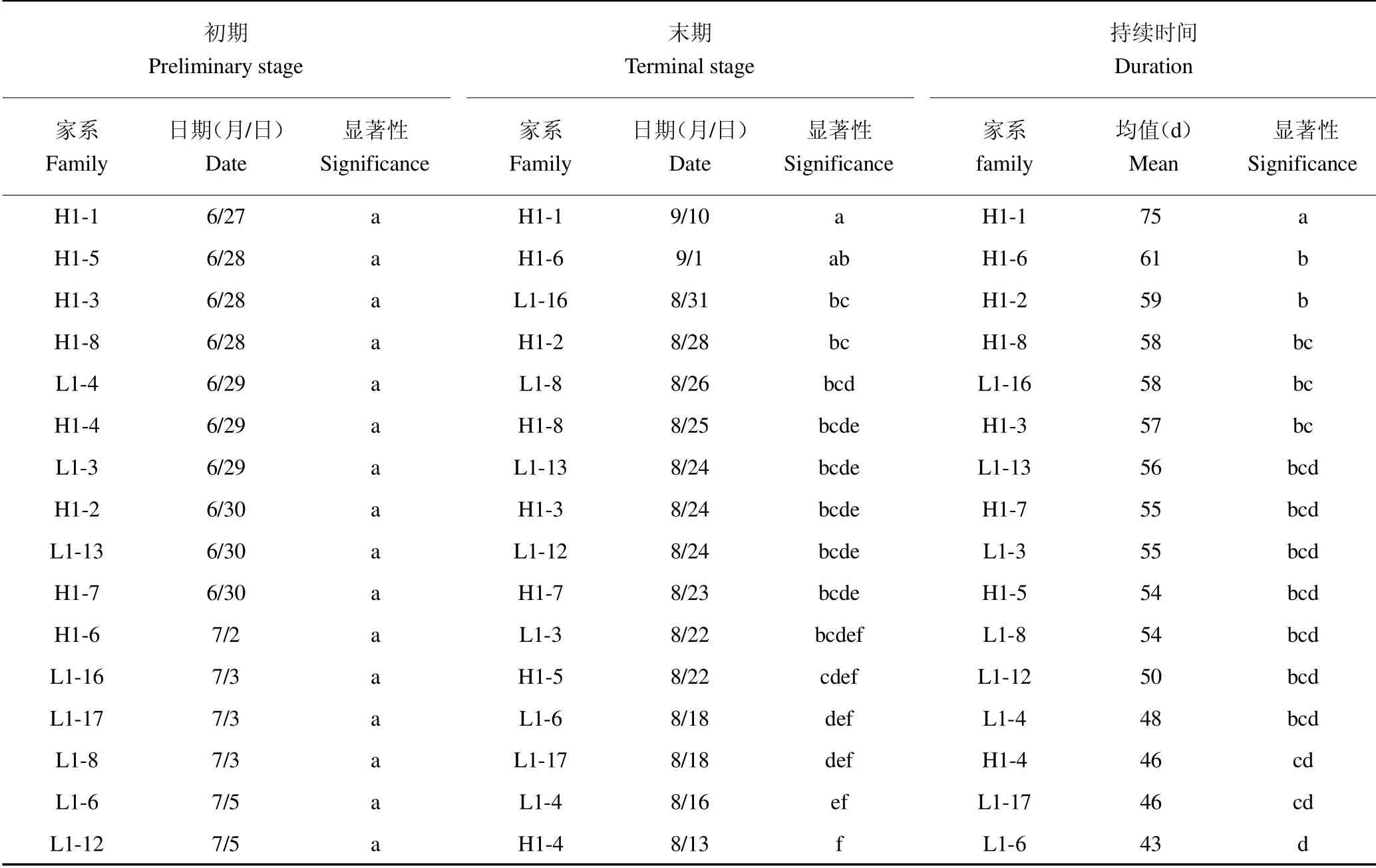
表3 家系间地径速生初期、末期及持续时间多重比较Table 3 Multiple comparisons of early,late and duration of rapid growth of ground diameter families
2.3 速生期聚类分析
根据黄檗苗高和地径速生初期、末期、速生点及持续时间,对黄檗家系进行聚类分析,将16 个家系分为“早型”、“中型”和“晚型”,见图1。其中“早型”家系为L1-3 号和L1-13 号,该类型家系苗期最早进入速生期,最晚结束速生期,且速生期持续时间较长;“晚型”家系有H1-4、L1-4、L1-6、L1-17 和L1-12 号,该类型家系苗期最晚进入速生期,最早结束速生期,且速生期持续时间较短;“中型”家系有H1-9、H1-3、H1-7、H1-5、L1-8、H1-2、H1-6、L1-16 和H1-1 号,该类型家系速生期处于其他两种类型之间。
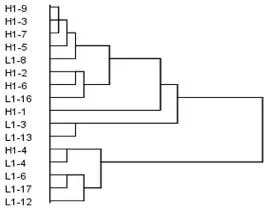
图1 黄檗苗期生长速生期聚类分析Fig.1 The cluster analysis of rapid growth at seedling growth
3 结论与讨论
黄檗半同胞子代苗期生长存在“慢 快 慢”的生长规律,符合“S”型生长曲线,苗高和地径生长模型拟合精度均大于0.8,这与傅大立等[13]对泡桐、魏蕾等[14]对杨树苗期生长的研究结果相近。根据黄檗苗木速生期始末时间,将苗期分为“前慢期”、“速生期”和“后慢期”,其中地径早于苗高进入速生期,且速生期持续时间较长。黄檗家系间生长速生期始期差异不显著,速生期末期、持续时间均达到显著差异。这与周永学等[15]对欧洲黑杨无性系间苗高和地径生长差异的研究结果相近,这说明遗传因素对苗期生长性状的影响较大。在16 个黄檗家系中,L1-3 号家系苗高生长于7 月25 日最早进入速生期,L1-6 号家系地径生长于7 月12 日最早进入速生期,且这两个家系均与多个家系间差异达到显著水平。
掌握植物苗期生长规律,适时浇水施肥,合理地进行苗期管理,有利于促进植物苗期生长。涂忠虞等[16]认为生长速生期是苗木生长的关键时期,苗木速生点和速生期持续时间可作为家系选择的重要指标。L1-3号家系苗高速生期持续时间最长,为49 d;H1-1 号家系地径速生期持续时间最长,为75 d。根据黄檗苗期生长节律,将16 个家系分为“早型”、“中型”和“晚型”。其中“早型”家系为L1-3 号和L1-13 号,该类型家系苗期最早进入速生期,最晚结束速生期,且速生期持续时间较长;“晚型”家系有H1-4、L1-4、L1-6、L1-17 和L1-12 号,该类型家系苗期最晚进入速生期,最早结束速生期,且速生期持续时间较短;“中型”家系有H1-9、H1-3、H1-7、H1-5、L1-8、H1-2、H1-6、L1-16 和H1-1 号,该类型家系速生期处于其他两种类型之间。赵曦阳等[17]认为根据苗木的速生期、速生点,制订合理的苗期管理措施,进行集约管理,可有效促进苗木的生长。在生产实践中可根据黄檗苗期生长节律类型筛选相同类型的家系进行集约管理,可有效提高苗圃的经营效率,节约成本。本文仅对黄檗不同家系苗期生长节律进行了研究,对苗期在不同管理措施下苗木生长情况有待于进一步探讨。
猜你喜欢
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
农业灾害研究(2022年2期)2022-05-31
南方农业·下旬(2022年4期)2022-05-24
安徽农学通报(2020年1期)2020-02-29
山东农业科学(2019年11期)2019-12-24
现代农业科技(2019年1期)2019-07-11
林业与生态(2019年6期)2019-07-02
森林工程(2018年1期)2018-05-14
现代农业科技(2017年24期)2018-01-22
