
动物机体精氨酸和赖氨酸功能互作效应与机制的研究进展
2022-06-13 12:36张益凡胡良宇王梦芝
中国畜牧杂志 2022年6期
张益凡,徐 颖,胡良宇,王梦芝,2*
(1.扬州大学动物科学与技术学院,江苏扬州 225009;2.新疆农垦科学院省部共建绵羊遗传改良与健康养殖国家重点实验室,新疆石河子 832000)
氨基酸是维持氮平衡必不可缺的物质,可以使动物处于最佳生长状态。不同动物种类所需的必需氨基酸也各不相同。精氨酸和赖氨酸是动物体生长发育环节所需的必需氨基酸,具有多种营养生理功能,能够促进生长发育及维持动物机体的氮平衡。在机体运转的各个过程中都具有重要作用。精氨酸和赖氨酸之间具有拮抗作用,且二者在日粮饲喂中也具有一定配比范围。关于精氨酸和赖氨酸拮抗作用的研究发现,赖氨酸浓度过高时肾脏中精氨酸酶代谢作用明显提高,而精氨酸和赖氨酸在机体各个方面都会呈现拮抗反应,二者在小肠的吸收及肾脏在重吸收方面的竞争作用也十分显著。精氨酸和赖氨酸利用相同类型的载体进行主动转运,若两者同时竞争同一载体,一方浓度过高便会导致另一方重吸收效率降低,同时排出的量也会显著增加。因此,需要进一步研究氨酸和赖氨酸的营养功能及日粮中的适宜比例。本文主要就精氨酸和赖氨酸的营养生理功能,以及二者的功能互作效应及其在日粮中的适宜比例做一综述。
1 精氨酸的理化特性及其对机体的营养生理功能
精氨酸的化学名称为2-氨基-5-胍基戊酸,属于氨基酸中的一种碱性氨基酸,主要以型和型2 种形式存在,发挥作用的只有型精氨酸。精氨酸参与多种机体蛋白质的合成,而参与合成的嘧啶同时也对生成DNA 和RNA 具有有效作用;其代谢产物的作用也十分广泛,包括调节人体免疫、增强免疫反应。精氨酸通过反应生成一氧化氮(NO),同时对特异性细胞具有刺激作用,能够调控免疫能力。除此之外,精氨酸还具有促进组织发育、促进繁殖等作用。
1.1 增强免疫功能 精氨酸及其代谢产物对于调节人体免疫力和提高自身免疫反应能力等都有很大的促进作用。研究表明,精氨酸可以促进激素(如生长激素、胰岛素)生成,加快机体自愈能力;还可以通过正向调控NK 细胞,增强巨噬细胞活性,提高机体抗感染、抗瘤能力。生成NO 必不可少的物质便是精氨酸,精氨酸经过催化反应可以生成产物NO,引起血管扩张,保持血液循环系统通畅,有效地调控免疫反应。闫伟等研究发现,扬州鹅饲粮中适量增加精氨酸可以提高免疫反应能力和法氏囊指数。Lieboldt 等研究发现,精氨酸含量与仔鸡淋巴器官发育具有联系,含量过低时会导致仔鸡的胸腺和脾脏发育迟缓。麻名文等发现,日粮增加精氨酸含量后肉兔的胸腺指数显著变化而脾脏指数无变化。师昆景等、高天试验发现,在胚蛋中直接注射精氨酸可以提高法氏囊指数,但该方法不适用于胸腺指数。
精氨酸具有调节免疫的作用,可以抑制也可以促进。当发生炎症反应时,适量精氨酸可以调节炎症反应,促进淋巴细胞增殖分化。马艳弘等研究表明,硒酸精氨酸可以增加活化分子蛋白的表达,同时提高NK 细胞的杀伤活力,从而提高免疫功能。对于肝癌患者术后的饮食要求即适量添加精氨酸,可以让衡量机体免疫水平的指标CD4升高、CD8降低,此时机体处于免疫状态,免疫水平有所提高。在小鼠日粮中添加精氨酸不仅能够优化肠道菌群结构,还可以增强先天性免疫。
1.2 促进肠道发育 肠道是维持机体内环境稳态的重要器官,而精氨酸可以促进肠道吸收消化。研究发现,在仔猪的日粮中加入适量精氨酸,可以使绒毛高度(VH)与隐窝深度(CD)的比值增大,且超氧化物歧化酶活性降低,仔猪肠黏膜生长加快。孟国权等研究发现,当肠道黏膜受脂多糖(LSP)抑制,黏膜屏障受到损害,可以在日粮中补充适量精氨酸来降低内皮质激素-1(ET-1)浓度及过氧化水平。当肠细胞增殖速度过快时可以利用雷帕霉素靶蛋白(mTOR)和Toll 样受体(TLR)信号通路机制进行抑制。当肉仔鸡不慎感染艾美尔球虫时,提高日粮中精氨酸含量,可显著增大VH 与CD 比值,且机体受感染情况能够有所改善。
1.3 提高抗氧化能力 细胞中存在的超氧化物歧化酶、过氧化氢酶对抗氧化作用具有促进作用,增加还原性谷胱甘肽(GSH)含量也可以增强动物的抗氧化能力。相关研究表明,-精氨酸发生氧化反应后,自身可以除去反应留下的残基,其反应产物NO 可以在很大程度上减少氧化应激反应带来的损害,同时提高抗氧化酶活性。研究结果发现,低氧诱导因子(1HIF-1)的表达量在热应激反应下迅速增加,血红素加氧酶-1(HMOX-1)能够有效抑制HIF-1 发生反应后造成的损害。由此表明,精氨酸既能抑制HIF-1m RNA 的表达又能促进HMOX-1m RNA 的表达,达到提高抗氧化能力的作用。
综上可见,精氨酸能够增强免疫功能、促进肠道发育、维持机体平衡、提高抗氧化能力、促进繁殖;且精氨酸与赖氨酸在消化、吸收、重吸收方面都存在着拮抗效应,在营养方面禽类对精氨酸需求更明显。同时,精氨酸在免疫方面的作用十分广泛,其应用前景广阔。但不同精氨酸浓度、动物品种、习性以及所构建的试验模型不同都可能导致研究结果的不同;对于精氨酸和赖氨酸的拮抗作用更待进一步研究,从而寻求各品类中最适宜的日粮精氨酸含量。
2 赖氨酸的理化特性及其对机体的营养生理功能
赖氨酸的化学名称为2,6-二氨基己酸,属于氨基酸中的碱性氨基酸,也是生糖兼生酮氨基酸,主要以型和型2 种形式存在,发挥作用的只有型赖氨酸。赖氨酸主要参与骨骼肌、酶、血清蛋白等多种蛋白质的合成,在进入机体后经小肠腔进入小肠黏膜细胞中,然后进入肝脏发生反应,最后合成蛋白。赖氨酸具有参与能量代谢、促进矿物质的吸收和骨骼生长、增强免疫功能等作用。除此之外,赖氨酸可以加快生长发育速度,具有调节血红蛋白、白蛋白、总蛋白的营养作用。
2.1 参与能量代谢 赖氨酸参与体内脂肪代谢中肉碱的生物合成。肉碱的生物作用十分广泛,可以促进脂肪新陈代谢、促进能量产生及降低胆固醇水平。在脂肪酸发生-氧化反应产生能量时,肉碱可以转化为脂酰肉碱将长链脂肪酸运输到线粒体膜外;在脂肪发生代谢、游离辅酶A 增加时,肉碱可以转化为酰基肉碱将短链酰基运输到线粒体膜外,增加脂肪代谢。肉碱也可以有效保护线粒体呼吸链,提高机体氧化供能。由必需营养素构成肉碱的合成,补充适量赖氨酸可以加快肉碱合成。前期试验发现,赖氨酸在能量代谢中起积极作用。在大鼠上的试验发现,脂肪肝现象主要是由于饲粮中的赖氨酸含量过低,导致肉碱不能合成或合成速度过慢,新陈代谢速度减慢,甘油三酯便会积聚在动物体内。也有试验证明,添加适量赖氨酸可以减少甘油三酯在体内的积聚现象,从而缓解脂肪肝现象,加快生长速度及新陈代谢速度。
2.2 促进矿物质的吸收和骨骼生长 赖氨酸能够与矿物质元素发生螯合反应形成环状结构的小分子配合物,从而促进矿物质吸收。给5 周龄大鼠分组服用不同量的赖氨酸,探索赖氨酸对骨骼肌蛋白的合成与降解的影响,发现赖氨酸可以维持机体稳态,主要是由自噬体-溶酶体系统抑制肌纤维蛋白降解。左云龙等、金俭英给40 名儿童服用-赖氨酸并观察其对生长发育的影响,结果表明添加-赖氨酸提高了儿童体内必需物质元素的浓度,且可以加快生长发育速度,调节血红蛋白、白蛋白、总蛋白的营养作用及促进矿物质吸收。除此之外,也有试验证明人体由于缺铁发生脱发时,可以摄入适量赖氨酸来缓解状况,达到利用赖氨酸促进铁吸收的目的。骨骼主要由胶原蛋白和矿物质成分组成,骨骼的生长与赖氨酸密不可分,添加适量赖氨酸可以促进骨骼肌增长。胶原蛋白由赖氨酸合成,且赖氨酸参与糖基化修饰与胶原分子间的共价交联,维持着骨骼韧性。
2.3 增强免疫功能 赖氨酸是连接抗原和T 细胞的非特异性分子,能够有效保护机体,从而增强免疫功能。当机体内赖氨酸含量过低,便会抑制蛋白质合成,损害自身免疫功能,从而提高动物感染疾病甚至死亡的概率,也会减少抗体产生,导致免疫力低下。因此,需要更多研究来探究日粮中赖氨酸含量过低会导致机体功能下降的问题。
3 精氨酸和赖氨酸在机体的代谢互作
3.1 赖氨酸和精氨酸的相互影响 20 世纪中期的研究表明,精氨酸和赖氨酸之间具有明显的拮抗作用。两者在吸收代谢、合成吸收各方向相互影响;两者的转运系统、载体类型相同,拮抗作用也十分显著。如果赖氨酸过量,会导致精氨酸降解速度加快,那么精氨酸的重吸收效率便会立刻大大降低,随尿液排出的精氨酸含量会随之增加。通过在肉鸡的日粮中添加赖氨酸,控制单一蛋白质为豆粕,改变粗蛋白质水平,结果表明精氨酸浓度会随粗蛋白质水平的提高而降低。调整28 日龄断奶仔猪日粮,仔猪血液中的精氨酸浓度会随赖氨酸水平的提高而降低。
3.2 氨基酸不平衡的影响 当猪本身已经具备了较强的自主排出体内过量精氨酸的能力,如果继续添加过量精氨酸和赖氨酸,会导致氨基酸不平衡。增加赖氨酸不能减少过量精氨酸造成负面影响,而过量精氨酸可以提高血液中的精氨酸浓度,降低赖氨酸浓度。研究表明,血清中的精氨酸浓度会随赖氨酸摄入的量增加而降低,而赖氨酸不会随摄入精氨酸含量的改变而改变。试验证明,当赖氨酸浓度提高到2.3%时对机体性能并无明显影响;过量赖氨酸可以提高血液中的赖氨酸浓度而精氨酸保持不变,但可以通过提高精氨酸酶的活性从而促进精氨酸降解。因此,在此情况下,赖氨酸过量并没有与精氨酸发生拮抗作用。
试验发现,赖氨酸浓度增加时,肾脏中共用系统的氨基酸重吸收受阻,尿中尿素氮含量并无影响,而赖氨酸、精氨酸等氨基酸含量增加。若在日粮中逐日提高赖氨酸含量,机体中游离氨基酸的含量会显著增加,而精氨酸会促进蛋白质合成而含量不变。机体内的精氨酸酶和鸟氨酸转氨甲酰酶活性不会因为赖氨酸浓度的提高随之变化。由此可见,摄入过量赖氨酸,不会影响精氨酸的新陈代谢反应,只会导致氨基酸不平衡。姚康等在7~21 日龄哺乳仔猪日粮中添加0.8%精氨酸,虽然仔猪体内精氨酸与赖氨酸的浓度会发生变化,但其生长性能并无明显变化,结果表明精氨酸与赖氨酸在仔猪上并无明显拮抗作用。
精氨酸对提高奶牛产雌率方面具有重要作用。王光辉等报道,在日粮中提高精氨酸含量,奶牛产雌率明显提高,与未提高精氨酸含量的奶牛组差异极为显著。范星根等通过对441 头奶牛于不同时间在日粮添加不同浓度精氨酸,发现母牛产雌率明显提高。精氨酸含量与受胎率有关,可能由于精氨酸与性染色体的睾丸决定因子有关。当机体中精氨酸浓度过低时,可以适当添加赖氨酸或精氨酸;但是当机体中赖氨酸浓度过低时,添加精氨酸并无影响。赖氨酸对提高奶牛泌乳量和乳蛋白率方面具有重要影响。当过瘤胃氨基酸引起乳脂率下降时,可以在日粮中同时添加2 种过瘤胃氨基酸。在荷斯坦奶牛的产前饲粮中添加过瘤胃赖氨酸可以降低体内精氨酸含量,提高乳蛋白含量,且奶牛对蛋白质和氨基酸的吸收利用率提高,随粪便排出的有效物质减少,从而提高利用率,减少污染排放,达到保护环境的目的。
对于大西洋鲑鱼,当日粮中的玉米蛋白适量,日粮中精氨酸含量远远超过赖氨酸含量时,生长发育速度便会加快,对氨基酸饲粮的吸收利用率也能提高;当日粮中赖氨酸含量远远超过赖氨酸时,生长发育速度也会加快,对氨基酸饲粮的吸收利用率也同样能提高。当日粮中精氨酸含量满足日粮所需时,赖氨酸含量的提高对生长发育速度无明显影响。由此也可以说明,精氨酸与赖氨酸在鱼类上并无明显拮抗作用,而是氨基酸不平衡特征较为显著。根据研究发现,鱼类与禽类对精氨酸的需求量都较大,而哺乳动物的尿素循环较发达,精氨酸来源不局限于日粮的饲喂。因此,动物品类的不同也会导致氨基酸营养关系不同。通过研究发现,肾脏精氨酸酶的活性会随摄入赖氨酸的量而发生变化,即摄入过量时活性便会增加。研究发现,调整大西洋鲑鱼日粮中精氨酸与赖氨酸的比例,赖氨酸对精氨酸的吸收能力有影响,而精氨酸对赖氨酸的吸收能力没有影响。
4 精氨酸与赖氨酸的代谢转运通路机制
4.1 转运系统的竞争互作 氨基酸转运系统分为酸性、中性和碱性转运系统,多种氨基酸转运系统共同完成机体的吸收转运。精氨酸和赖氨酸属于碱性氨基酸,主要通过相同的载体蛋白介导进行跨膜运输。转运蛋白可以与生物膜内外的物质进行信号交换以及摄取营养物质,其可帮助大部分亲水性化合物通过疏水屏障。在转运氨基酸时,根据是否需要Na协同分为依赖性与非依赖性2 种转运系统。
y+系统和y+L 系统可能决定了小肠细胞对碱性氨基酸的吸收。y+系统属于单向转运系统,主要是逆浓度转运聚集底物;y+L 系统属于反向交换转运,主要是重链4F2hc(CD98)和多个轻链结合形成,主要是转运碱性氨基酸(赖氨酸、精氨酸、组氨酸)。研究发现,两系统具有相似性,当对y+系统进行特异抑制只研究y+L 系统时发现,转运作用发生在肠细胞和肾小管的基底膜内,转入某氨基酸进入细胞时又将某氨基酸转出细胞。y+L 系统属于Na依赖性转运系统,其转运机理即反向交换转运。反向交换转运如图1 所示。经研究发现,适当提高中性氨基酸的浓度,依赖Na逆浓度梯度重吸收进入细胞,小肠内的吸收作用受到刺激,对碱性氨基酸的吸收更加充分,因此排出体外的碱性氨基酸含量减少,碱性氨基酸的利用率能够大大提升。
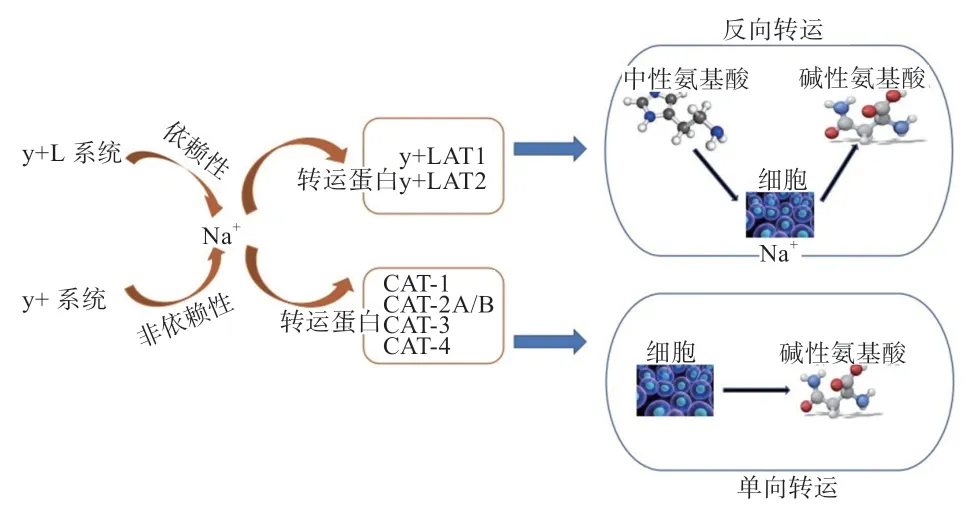
图1 氨基酸转运通路图
y+L 碱性氨基酸转运系统是由y+-型氨基酸转运载体(1y+LAT1)和y+-型氨基酸转运载体2(y+LAT2)各自与重链4F2 结合构成的异二聚体转运蛋白组成。y+LAT1 的作用是转运大分子氨基酸和芳香族氨基酸,可能在癌变细胞、恶性肿瘤上具有重要作用;而y+LAT2 只存在于上皮细胞,可以转运小分子中性氨基酸,y+LAT2 在医学营养上的生理功能尚未发现相关研究。
4.2 代谢通路的影响互作 摄入机体中的精氨酸有一部分在小肠内直接被分解,一部分进入其他组织器官进行吸收消化分解。精氨酸可能在精氨酸酶作用下直接分解为尿素和鸟氨酸,也可能在NOS 作用下直接分解为NO 和瓜氨酸。胃肠道和肾脏相互作用的情况下,瓜氨酸在精氨酸代琥珀酸合成酶和精氨酸代琥珀酸裂解酶的作用下合成内源精氨酸。
当日粮中赖氨酸浓度提高时,奶牛乳腺上皮细胞中的酪蛋白基因表达提高,并且能够有效调控基因表达,影响乳蛋白合成。因此,赖氨酸与奶牛乳腺上皮细胞的基因表达具有联系且具有依赖性,当赖氨酸浓度在0.5~2 mol/L 时能有效调控乳蛋白翻译相关基因表达。
精氨酸与赖氨酸具有相似性,其代谢通路也具有相似性。甲基化主要是发生在精氨酸与赖氨酸的残基上,由转化酶将活性甲基化合物上的甲基催化到其他化合物,从而进行基因表达的调控、蛋白质的调节。甲基化一般发生在氨基酸残基上,精氨酸可以被甲基化1次或2 次,而赖氨酸可以甲基化1 次、2 次或3 次。赖氨酸和精氨酸代谢通路及互作机制如图2 所示。
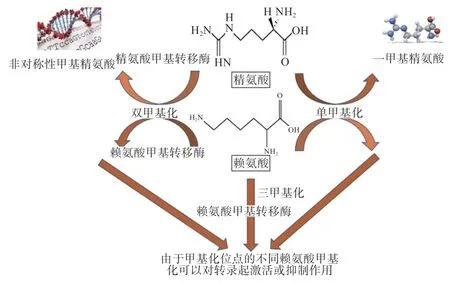
图2 精氨酸和赖氨酸代谢通路及互作机制
5 精氨酸与赖氨酸的适宜比例
当日粮中的赖氨酸含量过高时,会使动物生长速度降低,饲料利用率降低,蛋白质合成降低,生产性能降低,但是适当提高日粮中的精氨酸含量,可以提高动物生产性能,对蛋白质合成也没有不良影响。日粮中的赖氨酸含量过高时可能导致血清中游离精氨酸减少;精氨酸含量过高时对血清中的游离赖氨酸并无影响。
赖氨酸与精氨酸的最适比例并不是固定的,不同动物的适宜比例也大为不同。孙丰研究提出,肉仔鸡的生产性能不仅受赖氨酸和精氨酸的比例制约,温度也会影响肉仔鸡的性能。对于肉仔鸡,赖氨酸与精氨酸的最适比例是1:1.05~1:1.1。在适宜温度下保持赖氨酸与精氨酸的最适比例,肉仔鸡状态良好,如通过改变温度来提高胸肌重量时,肉仔鸡的胸肌重却直线下降。关于赖氨酸与精氨酸的最适比例,产蛋鸡为1.24:1;南美美利奴羊为1:1。在母猪日粮中添加精氨酸时其直肠温度下降,赖氨酸含量过高会减少采食量,精氨酸含量过高并无明显影响。但也有可能赖氨酸含量过高无明显影响,精氨酸含量过高会减少采食量。因此,针对赖氨酸与精氨酸的适宜比例还需要更多研究。
赖氨酸与精氨酸的最适比例在水生动物上也有所研究,白鲟最适比例1.13:1,幼虾最适比例1:1.12。Kim 等试验发现,虹鳟对于营养成分的需求与NRC(1998)仔猪相比只是精氨酸的需求量有差异,其他营养成分差异并不显著;虹鳟对于精氨酸与赖氨酸的需求量比应该在0.8:1~1.6:1,可根据实际情况进行调整。在日常饲喂时应均衡氨基酸含量,不能因种氨基酸有特殊作用便过量饲喂,可能导致毒害作用大于有效作用。
综上可见,对于精氨酸和赖氨酸的营养推荐量,除考虑单个氨基酸的比例外,还应确定二者在不同条件下的比例。
6 结语
精氨酸和赖氨酸存在于所有动物细胞内,具有重要作用。精氨酸可以增强自身免疫功能,促进肠道发育,提高抗氧化能力;赖氨酸可以促进矿物质的吸收和骨骼生长,参与能量代谢,增强免疫功能。但由于动物品种不同、两者在日粮中摄入浓度不同等多种因素,难以确定在动物体内发生作用的是氨基酸不平衡还是精氨酸与赖氨酸的拮抗作用。目前,在不同动物体内对两者的研究较少,未来需要根据动物的特性、摄入量,深入研究以确定两者之间的作用及最佳浓度比例,提高畜禽生长及生产效益。
猜你喜欢
中国畜牧杂志(2022年6期)2022-06-13
中国食品(2020年13期)2020-07-29
时尚育儿(2018年8期)2018-05-14
恋爱婚姻家庭·养生版(2018年3期)2018-03-24
农民致富之友(2017年4期)2017-04-10
农机使用与维修(2016年10期)2016-11-10
科技与创新(2015年11期)2015-06-16
新校园·中旬刊(2014年7期)2014-10-16
食品与生活(2011年2期)2011-02-21
大众健康(1999年7期)1999-06-06
