
花生疮痂病菌蓝光受体EaWC 1基因克隆及生物信息学分析
2022-06-10 03:25李洋张晓天朴静子周如军李自博关海雯
生物技术通报 2022年5期
李洋 张晓天 朴静子 周如军 李自博 关海雯
(沈阳农业大学植物保护学院,沈阳 110866)
光是细胞生物感知信号和适应环境重要的信息载体,在生长发育、有性生殖、次生代谢和昼夜节律等方面发挥着重要调控作用[1-2]。植物病原真菌的各种生理活动也同样受到光调控,众多研究结果表明,光对于真菌的形态建成、次生代谢和致病性等具有显著影响[3-4],但在不同种类真菌中的影响效应明显不同。光照影响布莱克须霉(Phycomyces blakesleeanus)的生长发育和形态建成,孢子囊柄弯曲方向具有明显的光趋向性[5-6],而对于灰霉菌(Botrytis cinerea),光照负调控菌核产生和致病力[7],光刺激竹黄菌(Shiraia bambusicola)分生孢子产量明显增加,但次生代谢产物竹红菌素(hypocrellin)产量却显著降低[8],这些研究证实真菌光效应种间差异显著。
真菌具有复杂且保守的光感系统-光受体(photoreceptor),主要包括蓝光受体、红光/远红光受体和绿光受体3种类型[9],由于蓝光受体更易受光刺激产生光信号,快速启动光感系统,调控下游次生代谢产物编码基因表达而备受关注[10-11]。Crosthwaite等[12]最早从粗糙脉孢菌(Neurospora crassa)中克隆得到第一个真菌蓝光受体基因WC 1,研究证实该受体对真菌昼夜节律、菌丝趋向性、无性发育、有性生殖和类胡萝卜素等次生代谢产物生物合成具有调控作用[13]。目前在构巢曲霉(Aspergillus nidulans)、烟曲霉(Aspergillus fumigatus)、禾谷镰孢菌(Fusarium graminearum)、卷枝毛霉菌(Mucor circinelloides)、深 绿 木 霉(Trichoderma atroviride)、交链格孢(Alternaria alternata)、灰霉菌(B. cinerea)和玉米灰斑病菌(Cercospora zeae maydis)等多种真菌中均筛选鉴定到蓝光受体WC 1同源基因[14-21],光照培养C. zeae maydis野生型菌株和突变体发现,蓝光受体基因CRP1存在能够抑制孢子产生,促进毒力因子尾孢菌素(Cercosporin)积累,CRP1基因缺失明显降低病菌致病性和病斑扩展能力[20]。
花生疮痂病(Elsinoë arachidis)是我国花生生产中的重要病害,发生普遍,危害严重。其致病菌能够产生一种光敏性苝醌类真菌毒素——痂囊腔菌素(elsinochromes,ESC),ESC是痂囊腔菌属重要的毒力因子,生物合成受到光调控[22]。目前,关于光调控ESC生物合成的研究报道相对较少,仅在柑橘疮痂病菌(E. fawcettii)和花生疮痂病菌(E. arachidis)中阐明了光照对ESC的生物合成具有正向调控作用[23-24],而光质对ESC生物合成影响效应及调控机制的相关研究尚未见报道。研究团队前期研究发现,蓝光能够显著促进花生疮痂病菌中ESC生物合成[24],基因组中存在蓝光受体WC 1同源基 因,命名为EaWC 1。在此基础上,本文开展了花生疮痂病菌蓝光受体EaWC 1基因克隆,生物信息学分析和表达模式研究,旨在为蓝光受体WC 1基因功能、ESC毒素生物合成受光调控机制和调控网络研究奠定理论基础。
1 材料与方法
1.1 材料
花 生 疮 痂 病(Elsinoë arachidis)菌 株LNJH-C01,分离自辽宁锦州花生种植基地。
1.2 方法
1.2.1 花生疮痂病菌DNA提取 菌株采用涂布培养法[24],25℃光照条件下培养14 d,设置光强度为30 μE·m-2·s-1,采用DNA提取试剂盒(天根生化科技有限公司)提取DNA,1.0%琼脂糖凝胶电泳,NanoDrop ND-2000分光光度计检测DNA质量,-20℃保存备用。
1.2.2 基因克隆 根据前期基因组数据进行蓝光受体基因序列引物设计[24],通过Primer 5.0软件设计引 物Eawc-1F:5'- CAGCCATGATGCAAGGAC-3',Eawc-1R:5'-TTGAGTGGGCGAGGCGTGT-3'。25 μL扩增体系,包括Mix酶12.5 μL,ddH2O 9.5 μL,引物各1 μL,DNA模板1 μL。扩增条件为94℃预变性5 min;94℃变性30 s,58℃退火30 s,72℃延伸3 min,40个循环;72℃延伸8 min,1%琼脂糖凝胶电泳。由生工生物工程(上海)股份有限公司测序。
1.2.3 生物信息学分析 用DNAMAN生物软件进行序列比对,利用ORF Finder、Softberry Web Site、ProtParam(https://web.expasy.org/protparam)、NetPhos 3.1 Server(http://www.cbs.dtu.dk/services/NetPhos/)、InterProScan、https://npsa-prabi.ibcp.fr/cgi-bin)、Swiss model(https://www.swissmodel.expasy.org)等在线软件分析开放阅读框、外显子、内含子、编码蛋白理化性质、保守结构域和二三级结构等信息,使用DOG 2.0软件绘制结构域。
1.2.4 系统发育分析 通过使用NCBI数据库,查找蓝光受体WC 1同源基因的氨基酸序列进行BLAST分析,利用MEGA 7.0软件构建系统进化树,基因序列信息见表1。

表1 WC 1同源基因序列信息Table 1 Sequence information of WC 1 homologous gene
1.2.5 花生疮痂病菌EaWC 1基因的表达分析 病菌PDA培养基中黑暗培养14 d后,置于不同光质的LED灯下(白光波长450-465 nm,蓝光波长490-530 nm),光强度为30 μE·m-2·s-1,以黑暗为对照,培养36 h,刮取菌丝,采用RNA提取试剂盒(普洛麦格生物技术有限公司)提取总RNA,置于-80℃保存。使用 HiFiScript cDNA Synthesis Kit反转录试剂盒(南京诺唯赞生物科技有限公司)反转录cDNA。以Actin基因为内参基因,利用Primer Premier 5.0 软件设计qPCR引物:EaWC-1-qF:5'-TATCGCCATCGGCACTCA-3',EaWC-1-qR:5'-CGGATTCGGAAGACTACA-3'。反 应 程 序:94℃ 2 min;94℃ 5 s,60℃ 15 s,72℃ 10 s,循环45次。每处理3次重复,以黑暗培养条件为对照,计算EaWC 1基因的相对表达量。
2 结果
2.1 EaWC 1基因克隆
从花生疮痂病菌中克隆EaWC 1基因(GenBank MZ274346),全长为3 261 bp(图1)。含有一条长度3 261 bp的完整开放阅读框,编码1 086 aa的蛋白,仅含有1个外显子,无内含子。

图1 EaWC 1基因PCR扩增Fig. 1 PCR amplification of EaWC 1 gene
2.2 蛋白结构预测
真菌蓝光受体WC-1具有保守结构域,包括一个LOV(light oxygen voltage)域、两个PAS(Per- Art-Ser)域和一个ZNF(zinc finger)结构域。结构域预测表明,花生疮痂病菌EaWC-1蛋白同样含有LOV(128 aa)、PAS(109 aa和114 aa)和ZNF(52 aa)结构域(图2),花生疮痂病菌结构域分布与N. crassa和C. zeae maydis等真菌中蓝光受体编码蛋白结构域基本吻合,仅在序列长度上存在细微差异。
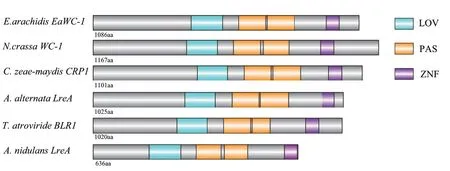
图2 WC-1 蛋白质结构域分析Fig.2 Protein domain analysis of WC-1
2.3 EaWC 1基因同源性分析
蓝光受体WC 1基因的LOV结构域可结合黄素色团感知蓝光。利用DNAMAN对真菌中蓝光受体WC 1同源基因LOV结构域的氨基酸序列进行比对,结果表明,花生疮痂病菌蓝光受体EaWC1的LOV结构域与比对蛋白序列同源性80.1%,拥有11个结合黄素色团所需残基(“↓”表示),以及参与光诱导反应的保守半胱氨酸残基(“*”表示)(图3)。

图3 WC-1蛋白氨基酸序列比对Fig. 3 Amino acid sequence alignment of WC-1 protein
BLAST分析结果表明,EaWC-1编码蛋白与粗糙脉孢菌(N. crassa)、葡萄黑痘病菌(E. ampelina)、 柑橘疮痂病菌(E. fawcettii)和玉米灰斑病菌(C. zeae maydis)等多种真菌的特征基因和预测基因具有较高的同源性(图4)。与葡萄黑痘病菌WC-1同源蛋白相似性最高达97.93%,其次为柑橘疮痂病菌(94.06%),EaWC-1分布在痂囊腔菌属形成的紧密分支中,表明所分析的序列为同源蛋白。
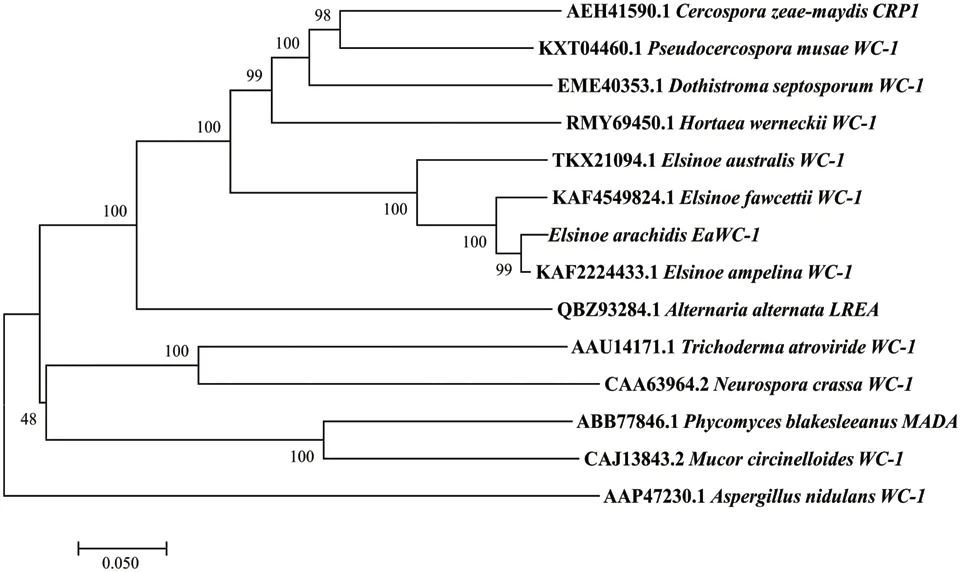
图 4 EaWC-1蛋白序列系统进化树Fig. 4 Phylogenetic tree of EaWC-1 protein sequence
2.4 EaWC 1基因生物信息学分析
2.4.1 理化性质及亲疏水性分析 通过ExPASy网站ProtParam对EaWC 1基因所编码蛋白的理化性质进行分析,分子量为119 563.30 Daltons,理论等电点(pI)为8.81,为碱性氨基酸。该蛋白脂溶性系数为57.92,不稳定系指数为49.71,为不稳定蛋白质;平均亲水性指数为(GRAVY)为-0.739。利用Protscale 对EaWC 1编码蛋白的疏水性/亲水性预测分析,表明EaWC 1编码蛋白在413位的I有最大值为2.133,疏水性最强,在948位的E有最小值为-3.711,亲水性最强,多肽链表现出亲水性(图5)。

图5 蛋白疏水性 Fig. 5 Protein hydrophobicity
2.4.2 编码蛋白磷酸位点及亚细胞定位 对EaWC 1基因编码蛋白是否存在可被激酶磷酸化的氨基酸残基进行分析,结果显示,该基因含有多个可被磷酸化氨基酸残基,说明该蛋白可以被激酶磷酸化后发挥作用(图6)。利用亚细胞定位在线分析工具分析,预测EaWC 1基因编码蛋白可能定位在细胞核。
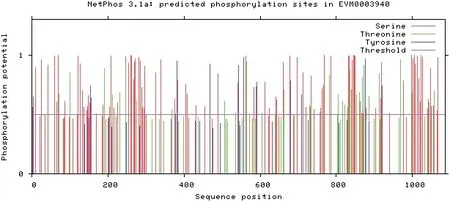
图6 编码蛋白磷酸位点分析Fig.6 Analysis of phosphate sites of encoded protein
2.4.3 蛋白信号肽及跨膜结构预测 使用在线工具分析EaWC-1蛋白是否存在信号肽及信号肽的剪切位点。分析显示,编码蛋白Sec信号肽(Sec/SPI)结构约为0.0005,不存在信号肽的剪切位点(CS),说明蛋白可能是非分泌蛋白。此外,跨膜结构预测表明,EaWC 1基因编码蛋白不存在跨膜螺旋结构域,为非膜蛋白。
2.4.4 蛋白质二三级结构 EaWC 1基因进行了二三级结构预测,其中α螺旋有283个,占比26.06%;β转角共有116个,占比10.68%;无规则卷曲共有509个,占比46.87%(图7)。

图7 蛋白二三级结构预测Fig. 7 Secondary and tertiary structure prediction of protein
2.5 基因表达量分析
光质对E. arachidis的ESC生物合成具有显著影响,白光条件下毒素含量为54 nmol/plug,蓝光处理促进ESC积累,毒素含量为71 nmol/plug,黑暗条件下未检测到毒素产生。qPCR分析表明,白光和蓝光条件下EaWC 1基因均显著上调表达,相对表达量分别为黑暗培养的7.48倍和14.62倍(图8-9),EaWC 1基因的表达模式与ESC毒素含量趋势相同,该基因参与调控ESC生物合成。
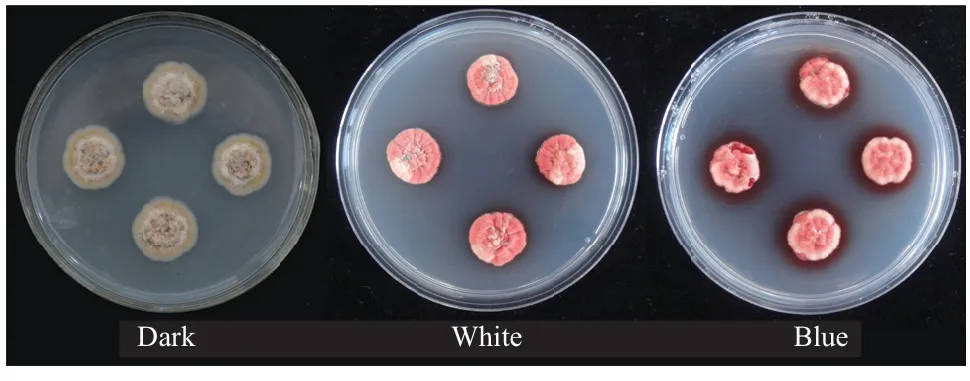
图8 病菌培养特性 Fig. 8 Culture characteristics of E. arachidis

图9 EaWC 1基因表达分析及ESC含量Fig.9 EaWC 1 expression analysis and ESC production light quality
3 讨论
细胞生物中光感应与传导是一个复杂的生物过程,蓝光受体是各种真菌生长发育和次生代谢等生理生化过程所必须的生物感受器。蓝光受体WC 1编码蛋白通常具有3个PAS结构域和一个锌指结构域,其中两个PAS域用于蛋白质-蛋白质互作,锌指结构域用于核定位[25-27],靠近N端的PAS域属于一类特殊的结构域,称为LOV结构域,也称光、氧或电压域。LOV结构域含有一个半胱氨酸残基,光照条件下半胱氨酸与黄素色团(FAD)结合,发生电子转移,形成半胱氨酰复合物,黑暗条件下,此过程发生逆转,完成光周期循环[27-28]。虽然真菌蓝光受体WC 1基因的结构域高度保守,但基因功能研究证明,该基因在种间的生物功能有所差异,模式真菌粗糙脉孢菌中WC 1基因影响生物钟和类胡萝卜素的生物合成[26],而蓝光受体Fgwc 1基因影响禾谷镰孢菌有性繁殖和无性发育[16]。本研究在花生疮痂病菌中克隆获得蓝光受体EaWC 1基因,对其进行生物信息学和同源聚类分析发现,该基因全长序列为3 261 bp,具有一个完整的开放阅读框,编码长度为1 086个氨基酸的蛋白质。具有明显的蓝光受体编码基因WC 1的保守结构域,即一个LOV结构域,两个PAS域和一个ZNF结构域。系统发育分析发现,EaWC 1与粗糙脉孢菌(N. crassa)、柑橘疮痂病菌(E. fawcettii)和玉米灰斑病菌(C. zeae maydis)等真菌的蓝光受体基因有高同源性,研究结果为花生疮痂病菌蓝光受体基因EaWC 1的功能及毒素光调控模式研究奠定了分子基础。
基因表达是将遗传信息通过转录、剪接和翻译等方式转变成功能产物的所有加工过程,能够体现内部生理生化反映的调控及应答效应,是生物生命活动的基础和关键[29]。光生物学研究表明,蓝光受体WC 1基因的表达调控与多种真菌次生代谢产物的生物合成具有高度关联性。光照能够促进水稻恶苗病菌(F. fujikuroi)中镰孢菌素(Fusarins)的生物合成,此时蓝光受体WcoA基因高表达[30],蓝光受体LreA基因正向调控链格孢菌中Altertoxin(ATX)生物合成,但反向调节Alternariol(AOH)毒素产生[19]。花生疮痂病菌ESC生物合成与光照密切相关,但光调控机制尚不清晰,本文开展了毒素含量与EaWC 1基因表达量相关性研究,结果表明,白光和蓝光条件下EaWC 1基因均上调表达,蓝光培养条件下基因表达量是白光下的2.15倍,其表达模式与ESC毒素含量趋势相同,进而证明了花生疮痂病菌ESC生物合成受到EaWC 1基因调控。目前除粗糙脉孢菌、构巢曲霉和链格孢菌等真菌蓝光受体研究相对深入外,痂囊腔菌属的光响应研究主要集中在生物学表型及表达量研究方面,而基因功能研究相对较少,调控模式和分子机制尚不清晰,应尽快开展蓝光受体EaWC 1基因功能及ESC毒素生物合成光调控机制研究工作,为该病菌的致病机制、病原寄主互作和精准防控提供科学支撑。
4 结论
本研究成功克隆获得了花生疮痂病菌蓝光受体编码基因EaWC 1,全长为3 261 bp,编码蛋白含1 086氨基酸,为不含信号肽和跨膜螺旋结构的碱性亲水蛋白,主要定位于细胞核上。
光照条件显著促进蓝光受体基因EaWC 1上调表达,蓝光条件下基因表达量最高,表达模式与ESC毒素含量趋势相同,光受体基因EaWC 1在花生疮痂病菌ESC毒素生物合成光调控过程中发挥着重要作用。
猜你喜欢
湖北农业科学(2022年11期)2022-07-18
实用肿瘤学杂志(2020年4期)2020-12-08
浙江农业科学(2019年10期)2019-10-23
家庭影院技术(2019年8期)2019-08-27
家庭影院技术(2019年1期)2019-01-21
蔬菜(2019年4期)2019-01-04
家庭影院技术(2018年10期)2018-11-02
山西果树(2015年3期)2015-05-29
中国老年学杂志(2015年9期)2015-01-31
中国果业信息(2014年4期)2014-01-24
